
Synergistic In Vitro Antimicrobial Activity of Pomegranate Rind Extract and Zinc (II) against Micrococcus luteus under Planktonic and Biofilm Conditions

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Pomegranate Rind Extract (PRE)
2.3. Characterization of PRE
2.4. Microorganism and Test Solutions
2.5. Microdilution Method
2.6. In Vitro Evaluation Synergistic/Antagonistic Activity of PRE and Zn (II)
2.6.1. Checkerboard Assay
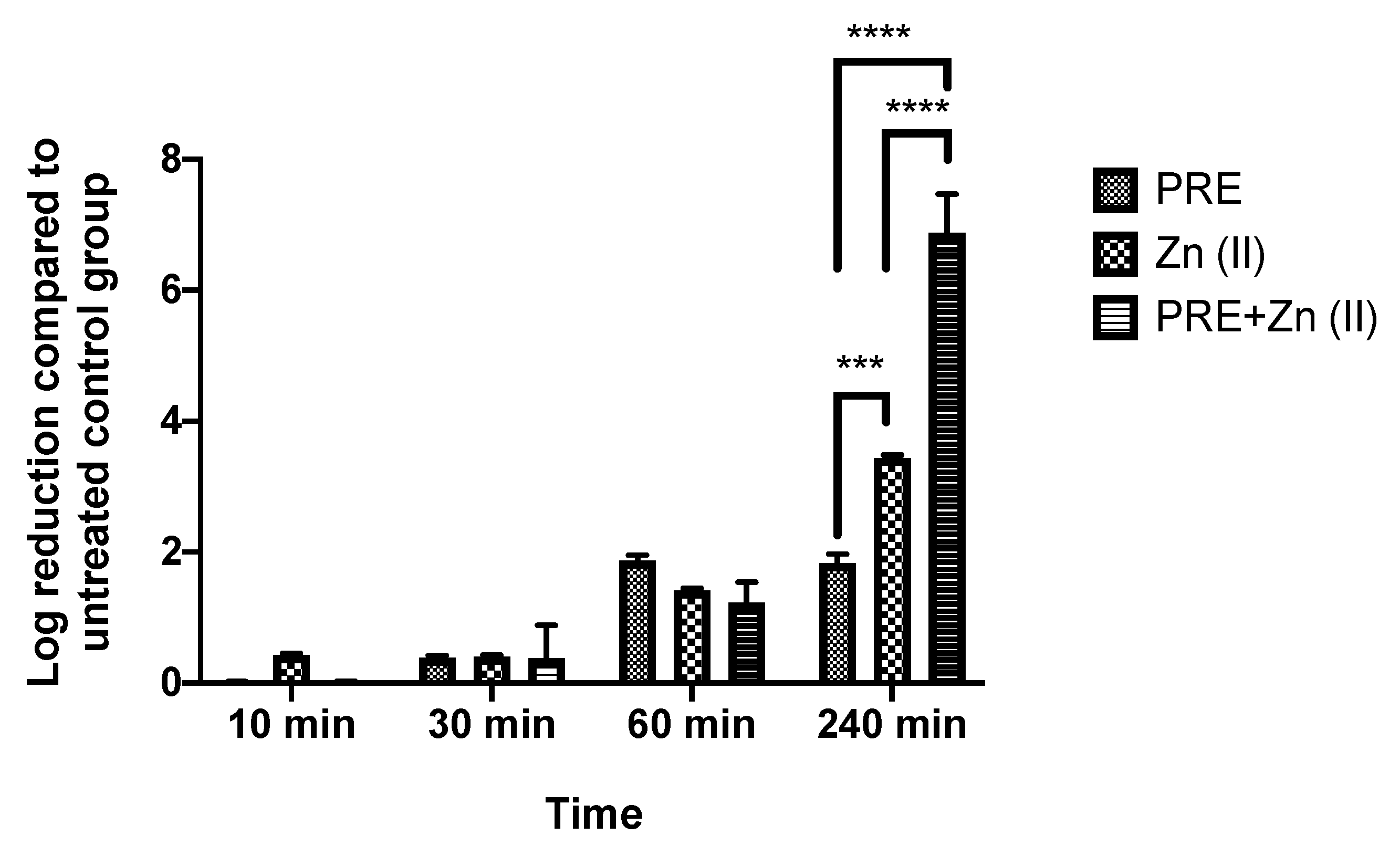
2.6.2. Time-Kill Assay
2.7. In Vitro Biofilm Inhibition and Eradication Activity of PRE, Zn (II), and PRE/Zn (II) Combination
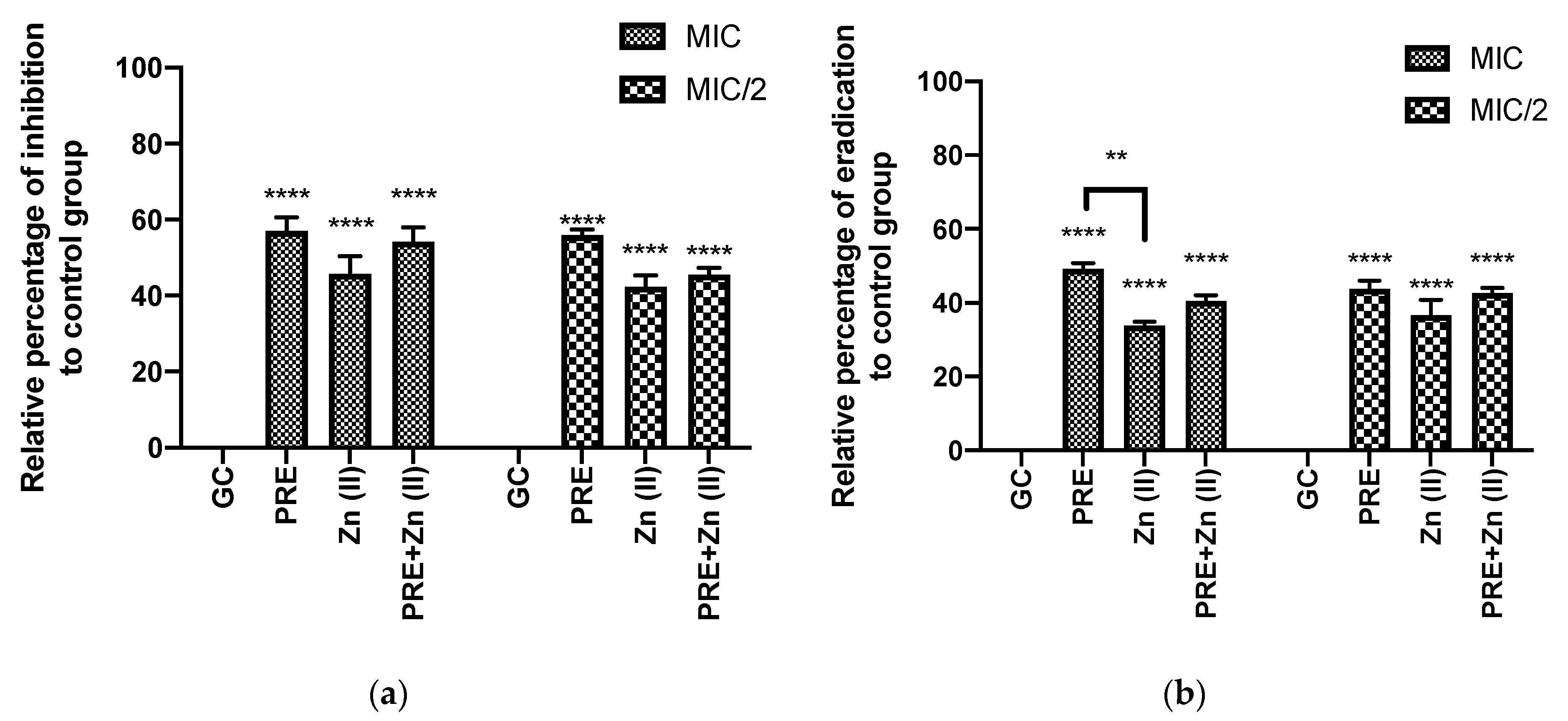
2.7.1. Biofilm Inhibition and Eradication Assay
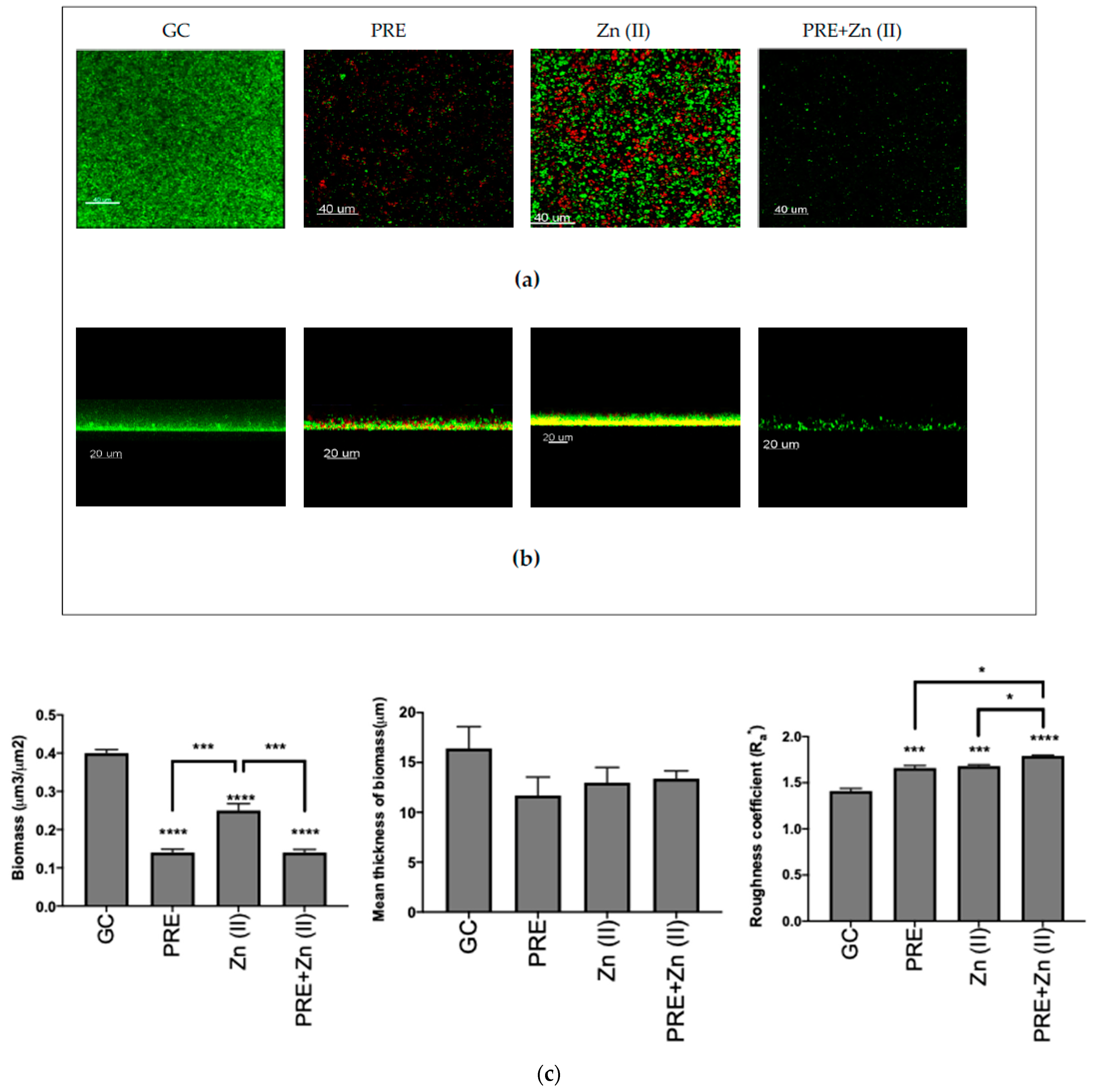
2.7.2. Fluorescence Microscopy Analysis of Biofilm Eradication
2.8. Statistical Analysis
3. Results
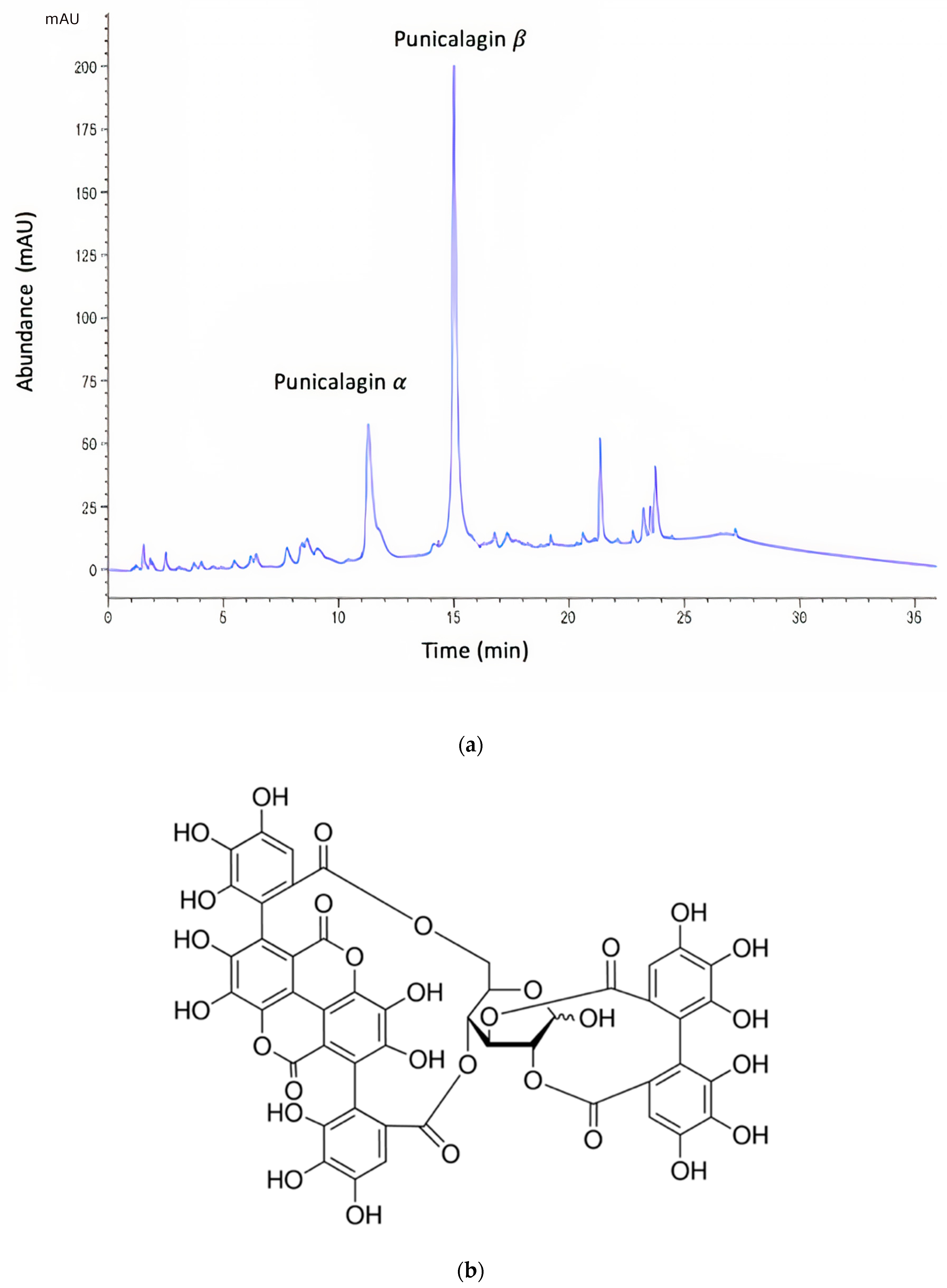
3.1. Characterization of PRE
3.2. Determination of MIC and MBC Values of PRE and Zn (II)
3.3. Determination of Synergistic Activity of PRE/Zn (II) in Combination
3.4. Biofilm Inhibition and Eradication Activity of PRE, Zn (II) and PRE/Zn (II) in Combination
3.4.1. Percentage of Biofilm Inhibition and Eradication through In Vitro Crystal Violet Assay
3.4.2. Fluorescence Microscopy Analysis of Biofilm Eradication
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Stefan, W. Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Bowen, W.H.; Burne, R.A.; Wu, H.; Koo, H. Oral Biofilms: Pathogens, Matrix, and Polymicrobial Interactions in Microenvironments. Trends Microbiol. 2018, 26, 229–242. [Google Scholar] [CrossRef]
- Bernhard, D.; Schwaiger, W.; Crazzolara, R.; Tinhofer, I.; Kofler, R.; Csordas, A. Enhanced MTT-reducing activity under growth inhibition by resveratrol in CEM-C7H2 lymphocytic leukemia cells. Cancer Lett. 2003, 195, 193–199. [Google Scholar] [CrossRef]
- Ramos, E.R.; Hachem, R.; Youssef, S.; Fang, X.; Jiang, Y.; Raad, I. The Crucial Role of Catheters in Micrococcal Bloodstream Infections in Cancer Patients. Infect. Control. Hosp. Epidemiol. 2009, 30, 83–85. [Google Scholar] [CrossRef]
- Ianniello, N.M.; Andrade, D.C.; Ivancic, S.; Eckardt, P.A.; Ramirez, J.C.L. Native valve infective endocarditis due to Micrococcus luteus in a non-Hodgkin’s lymphoma patient. IDCases 2019, 18, e00657. [Google Scholar] [CrossRef]
- Fosse, T.; Toga, B.; Peloux, Y.; Granthil, C.; Bertrando, J.; Sethian, M. Meningitis due to Micrococcus luteus. Infection 1985, 13, 280–281. [Google Scholar] [CrossRef]
- Selladurai, B.M.; Sivakumaran, S.; Aiyar, S.; Mohamad, A.R. Intracranial suppuration caused by Micrococcus luteus. Br. J. Neurosurg. 1993, 7, 205–207. [Google Scholar] [CrossRef]
- Wharton, M.; Rice, J.R.; McCallum, R.; Gallis, H.A. Septic arthritis due to Micrococcus luteus. J. Rheumatol. 1986, 13, 659–660. [Google Scholar]
- Peces, R.; Gago, E.; Tejada, F.; Laures, A.S.; Alvarez-Grande, J. Relapsing bacteraemia due to Micrococcus luteus in a haemodialysis patient with a Perm-Cath catheter. Nephrol. Dial. Transplant. 1997, 12, 2428–2429. [Google Scholar] [CrossRef]
- Shanks, D.; Goldwater, P.; Pena, A.; Saxon, B. Fatal Micrococcus sp. infection in a child with leukaemia—A cautionary case. Med. Pediatr. Oncol. 2001, 37, 553–554. [Google Scholar] [CrossRef]
- Richardson, J.; Marples, R.; De Saxe, M. Characters of coagulase-negative staphylococci and micrococci from cases of endocarditis. J. Hosp. Infect. 1984, 5, 164–171. [Google Scholar] [CrossRef]
- Oudiz, R.J.; Widlitz, A.; Beckmann, X.J.; Camanga, D.; Alfie, J.; Brundage, B.H.; Barst, R.J. Micrococcus-Associated Central Venous Catheter Infection in Patients With Pulmonary Arterial Hypertension. Chest 2004, 126, 90–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldock, E.; Surewaard, B.G.J.; Shamarina, D.; Na, M.; Fei, Y.; Ali, A.; Williams, A.; Pollitt, E.J.G.; Szkuta, P.; Morris, P.; et al. Human skin commensals augment Staphylococcus aureus pathogenesis. Nat. Microbiol. 2018, 3, 881–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rensburg, J.J.; Lin, H.; Gao, X.; Toh, E.; Fortney, K.R.; Ellinger, S.; Zwickl, B.; Janowicz, D.M.; Katz, B.P.; Nelson, D.E.; et al. The human skin microbiome associates with the outcome of and is influenced by bacterial infection. MBio 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Miltiadous, G.; Elisaf, M. Native valve endocarditis due to Micrococcus luteus: A case report and review of the literature. J. Med Case Rep. 2011, 5, 251. [Google Scholar] [CrossRef] [Green Version]
- Eady, E.A.; Coates, P.; Ross, J.I.; Ratyal, A.H.; Cove, J.H. Antibiotic resistance patterns of aerobic coryneforms and furazolidone-resistant Gram-positive cocci from the skin surface of the human axilla and fourth toe cleft. J. Antimicrob. Chemother. 2000, 46, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Liebl, W.; Kloos, W.E.; Ludwig, W. Plasmid-borne macrolide resistance in Micrococcus luteusaaThe GenBank accession number for the sequence reported in this paper is AF462611. Microbiology 2002, 148, 2479–2487. [Google Scholar] [CrossRef] [Green Version]
- Szczerba, I. Susceptibility to antibiotics of bacteria from genera Micrococcus, Kocuria, Nesterenkonia, Kytococcus and Dermacoccus. Med. Doświadczalna Mikrobiol. 2003, 55, 79. [Google Scholar]
- Erbasan, F. Brain abscess caused by Micrococcus luteus in a patient with systemic lupus erythematosus: Case-based review. Rheumatology international. 2018, 38, 2323–2328. [Google Scholar] [CrossRef]
- Dürst, U.N.; Bruder, E.; Egloff, L.; Wüst, J.; Schneider, J.; O Hirzel, H. Micrococcus luteus: A rare pathogen of valve prosthesis endocarditis. Z. Kardiol. 1991, 80. [Google Scholar]
- Maki, D.G. Infections caused by intravascular devices used for infusion therapy: Pathogenesis, prevention, and management. Infect. Assoc. Indwelling Med. Devices 1994, 2, 155–212. [Google Scholar]
- Maki, D.G.; Stolz, S.M.; Wheeler, S.; Mermel, L.A. Prevention of central venous catheter-related bloodstream infection by use of an antiseptic-impregnated catheter: A randomized, controlled trial. Ann. Intern. Med. 1997, 127, 257–266. [Google Scholar] [CrossRef]
- Williamson, E.M.; Evans, F.J.; Wren, R.C. Potter’s New Cyclopaedia of Botanical Drugs and Preparations; Daniel Company Limited: Saffron Walden (GB), UK, 1988. [Google Scholar]
- Seeram, N.P.; Zhang, Y.; Reed, J.; Krueger, C.; Vaya, J. Commercialization of pomegranates. In Pomegranates: Ancient Roots to Modern Medicine; Seeram, N.P., Schulman, R.N., Heber, D., Eds.; CRC Press: Boca Raton, FL, USA, 2006; Volume 43, pp. 193–195. [Google Scholar]
- Ismail, T.; Sestili, P.; Akhtar, S. Pomegranate peel and fruit extracts: A review of potential anti-inflammatory and anti-infective effects. J. Ethnopharmacol. 2012, 143, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Adams, L.S.; Seeram, N.P.; Aggarwal, B.B.; Takada, Y.; Sand, D.; Heber, D. Pomegranate Juice, Total Pomegranate Ellagitannins, and Punicalagin Suppress Inflammatory Cell Signaling in Colon Cancer Cells. J. Agric. Food Chem. 2006, 54, 980–985. [Google Scholar] [CrossRef] [PubMed]
- Houston, D.M.; Robins, B.; Bugert, J.J.; Denyer, S.P.; Heard, C.M. In vitro permeation and biological activity of punicalagin and zinc (II) across skin and mucous membranes prone to Herpes simplex virus infection. Eur. J. Pharm. Sci. 2017, 96, 99–106. [Google Scholar] [CrossRef]
- McCarrell, E.M.; Gould, S.W.J.; Fielder, M.D.; Kelly, A.F.; El Sankary, W.; Naughton, D.P. Antimicrobial activities of pomegranate rind extracts: Enhancement by addition of metal salts and vitamin C. BMC Complement. Altern. Med. 2008, 8, 64–67. [Google Scholar] [CrossRef] [Green Version]
- Stewart, G.S.; A Jassim, S.; Denyer, S.P.; Newby, P.; Linley, K.; Dhir, V.K. The specific and sensitive detection of bacterial pathogens within 4 h using bacteriophage amplification. J. Appl. Microbiol. 1998, 84, 777–783. [Google Scholar] [CrossRef]
- Yehia, H.M.; Elkhadragy, M.F.; Moneim, A.E. Antimicrobial activity of pomegranate rind peel extracts. Afr. J. Microbiol. Res. 2011, 5, 3664–3668. [Google Scholar]
- Houston, D. Towards a Nanomedicine-Based Broad-Spectrum Topical Virucidal Therapeutic System. Ph.D. Thesis, Cardiff University, Cardiff, UK, 2011. [Google Scholar]
- A Ainsworth, E.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.; Lee, R.; Hardy, M.; Heber, D. Rapid large scale purification of ellagitannins from pomegranate husk, a by-product of the commercial juice industry. Sep. Purif. Technol. 2005, 41, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; Approved Standard M07-A9; CLSI: Wayne, PA, USA, 2012. [Google Scholar]
- National Committee for Clinical Laboratory Standards. Methods for Determining Bactericidal Activity of Antimicrobial Agents: Approved Guideline M26-A; NCCLS: Wayne, PA, USA, 1999. [Google Scholar]
- Endo, E.H.; Ueda-Nakamura, T.; Nakamura, C.V.; Filho, B.P.D. Activity of Spray-dried Microparticles Containing Pomegranate Peel Extract against Candida albicans. Molecules 2012, 17, 10094–10107. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef] [PubMed]
- Johnston, M.; Lambert, R.; Hanlon, G.; Denyer, S. A rapid method for assessing the suitability of quenching agents for individual biocides as well as combinations. J. Appl. Microbiol. 2002, 92, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Miles, A.A.; Misra, S.S.; Irwin, J.O. The estimation of the bactericidal power of the blood. Epidemiol. Infect. 1938, 38, 732–749. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, F.B.; Chaieb, K.; Zmantar, T.; Kallel, H.; Bakhrouf, A. Adherence assays and slime production of Vibrio alginolyticus and Vibrio parahaemolyticus. Braz. J. Microbiol. 2009, 40, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, S.; Shah, R.; Bhave, M.; Palombo, E. Inhibitory activity of yarrow essential oil on Listeria planktonic cells and biofilms. Food Control. 2013, 29, 125–130. [Google Scholar] [CrossRef]
- Powell, L.; Pritchard, M.F.; Ferguson, E.; Powell, K.A.; Patel, S.U.; Rye, P.; Sakellakou, S.-M.; Buurma, N.J.; Brilliant, C.D.; Copping, J.M.; et al. Targeted disruption of the extracellular polymeric network of Pseudomonas aeruginosa biofilms by alginate oligosaccharides. npj Biofilms Microbiomes 2018, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersbøll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program comstat. Microbiology. 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Ding, K.; Yuan, Q. Determination of Punicalagin Isomers in Pomegranate Husk. Chromatographia 2008, 68, 303–306. [Google Scholar] [CrossRef] [Green Version]
- Opara, L.U.; Al-Ani, M.R.; Al-Shuaibi, Y.S. Physico-chemical Properties, Vitamin C Content, and Antimicrobial Properties of Pomegranate Fruit (Punica granatum L.). Food Bioprocess Technol. 2009, 2, 315–321. [Google Scholar] [CrossRef]
- Naz, S.; Siddiqi, R.; Ahmad, S.; Rasool, S.; Sayeed, S. Antibacterial Activity Directed Isolation of Compounds from Punica granatum. J. Food Sci. 2007, 72, M341–M345. [Google Scholar] [CrossRef]
- Al-Zoreky, N.S. Antimicrobial activity of pomegranate (Punica granatum L.) fruit peels. Int. J. Food Microbiol. 2009, 134, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Meléndez, P.; Capriles, V. Antibacterial properties of tropical plants from Puerto Rico. Phytomedicine 2006, 13, 272–276. [Google Scholar] [CrossRef]
- Dahham, S.S.; Ali, M.N.; Tabassum, H.; Khan, M. Studies on antibacterial and antifungal activity of pomegranate (Punica granatum L.). Am. Eurasian J. Agric. Environ. Sci. 2010, 9, 273–281. [Google Scholar]
- Negi, P.; Jayaprakasha, G. Antioxidant and Antibacterial Activities of Punica granatum Peel Extracts. J. Food Sci. 2003, 68, 1473–1477. [Google Scholar] [CrossRef]
- Duman, A.D.; Ozgen, M.; Dayisoylu, K.S.; Erbil, N.; Durgaç, C. Antimicrobial Activity of Six Pomegranate (Punica granatum L.) Varieties and Their Relation to Some of Their Pomological and Phytonutrient Characteristics. Molecules 2009, 14, 1808–1817. [Google Scholar] [CrossRef] [Green Version]
- Kumari, M.; Mahajan, H.; Joshi, R.; Gupta, M. Development and structural characterization of edible films for improving fruit quality. Food Packag. Shelf Life 2017, 12, 42–50. [Google Scholar] [CrossRef]
- Reddy, M.K.; Gupta, S.K.; Jacob, M.R.; Khan, S.I.; Ferreira, D. Antioxidant, Antimalarial and Antimicrobial Activities of Tannin-Rich Fractions, Ellagitannins and Phenolic Acids from Punica granatum L. Planta Medica 2007, 73, 461–467. [Google Scholar] [CrossRef]
- Xu, Y.; Shi, C.; Wu, Q.; Zheng, Z.; Liu, P.; Li, G.; Peng, X.; Xia, X. Antimicrobial Activity of Punicalagin Against Staphylococcus aureus and Its Effect on Biofilm Formation. Foodborne Pathog. Dis. 2017, 14, 282–287. [Google Scholar] [CrossRef]
- Holland, D.; Bar-Ya’akov, I. Pomegranate: Aspects concerning dynamics of health beneficial phytochemicals and therapeutic properties with respect to the tree cultivar and the environment. In Medicinal and Aromatic Plants of the Middle-East; Springer: Dordrecht, The Netherlands, 2014; pp. 225–239. [Google Scholar]
- Carlsson, J. Dental plaque as a source of salivary streptococci. Odontol. Rev. 1967, 18, 173–178. [Google Scholar]
- Gibbons, R.J.; Houte, J.V. Bacterial Adherence in Oral Microbial Ecology. Annu. Rev. Microbiol. 1975, 29, 19–42. [Google Scholar] [CrossRef]
- Serra, R.; Grande, R.; Butrico, L.; Rossi, A.; Settimio, U.F.; Caroleo, B.; Amato, B.; Gallelli, L.; de Franciscis, S. Chronic wound infections: The role of Pseudomonas aeruginosa and Staphylococcus aureus. Expert Rev. Anti-Infect. Ther. 2015, 13, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Malic, S.; Hill, K.E.; Hayes, A.; Percival, S.L.; Thomas, D.W.; Williams, D. Detection and identification of specific bacteria in wound biofilms using peptide nucleic acid fluorescent in situ hybridization (PNA FISH). Microbiology 2009, 155, 2603–2611. [Google Scholar] [CrossRef] [Green Version]
- Dowd, S.E.; Wolcott, R.D.; Sun, Y.; McKeehan, T.; Smith, E.; Rhoads, D. Polymicrobial Nature of Chronic Diabetic Foot Ulcer Biofilm Infections Determined Using Bacterial Tag Encoded FLX Amplicon Pyrosequencing (bTEFAP). PLoS ONE 2008, 3, e3326. [Google Scholar] [CrossRef]
- Malic, S.; Hill, K.; Playle, R.; Thomas, D.; Williams, D. In vitro interaction of chronic wound bacteria in biofilms. J. Wound Care 2011, 20, 569–577. [Google Scholar] [CrossRef]
- Wu, C.; Labrie, J.; Tremblay, Y.; Haine, D.; Mourez, M.; Jacques, M. Zinc as an agent for the prevention of biofilm formation by pathogenic bacteria. J. Appl. Microbiol. 2013, 115, 30–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, J.J.; Ceri, H.; Stremick, C.A.; Turner, R.J. Biofilm susceptibility to metal toxicity. Environ. Microbiol. 2004, 6, 1220–1227. [Google Scholar] [CrossRef]
- Ivanova, I.A.; Tasheva-Terzieva, E.; Angelov, O.; Krusteva, L.; Andonova, I.; Papazova, K.; Dimova-Malinovska, D.; Dushkin, C. Effect of ZnO thin films on survival of Pseudomonas cells. J. Nanomed Nanotechol. 2012, 3, 2. [Google Scholar]
- Zhao, L.; Chu, P.K.; Zhang, Y.; Wu, Z. Antibacterial coatings on titanium implants. J. Biomed. Mater. Res. Part B Appl. Biomater. 2009, 91, 470–480. [Google Scholar] [CrossRef]
- Eshed, M.; Lellouche, J.; Matalon, S.; Gedanken, A.; Banin, E. Sonochemical Coatings of ZnO and CuO Nanoparticles Inhibit Streptococcus mutans Biofilm Formation on Teeth Model. Langmuir 2012, 28, 12288–12295. [Google Scholar] [CrossRef]
- Jesline, A.; John, N.P.; Narayanan, P.M.; Vani, C.; Murugan, S. Antimicrobial activity of zinc and titanium dioxide nanoparticles against biofilm-producing methicillin-resistant Staphylococcus aureus. Appl. Nanosci. 2014, 5, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Benslimane, S.; Rebai, O.; Djibaoui, R.; Arabi, A. Pomegranate Peel Extract Activities as Antioxidant and Antibiofilm against Bacteria Isolated from Caries and Supragingival Plaque. Jordan J. Biol. Sci. 2020, 13, 403–412. [Google Scholar]
- Bakkiyaraj, D.; Nandhini, J.R.; Malathy, B.; Pandian, S.K. The anti-biofilm potential of pomegranate (Punica granatum L.) extract against human bacterial and fungal pathogens. Biofouling 2013, 29, 929–937. [Google Scholar] [CrossRef]
- Beyenal, H.; Donovan, C.; Lewandowski, Z.; Harkin, G. Three-dimensional biofilm structure quantification. J. Microbiol. Methods 2004, 59, 395–413. [Google Scholar] [CrossRef]
- Battin, T.J.; Kaplan, L.A.; Newbold, J.D.; Cheng, X.; Hansen, C. Effects of Current Velocity on the Nascent Architecture of Stream Microbial Biofilms. Appl. Environ. Microbiol. 2003, 69, 5443–5452. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.; Haagensen, J.A.J.; Heydorn, A.; Molin, S. Metabolic Commensalism and Competition in a Two-Species Microbial Consortium. Appl. Environ. Microbiol. 2002, 68, 2495–2502. [Google Scholar] [CrossRef] [Green Version]
- Yap, R.L.; Mermel, L.A. Micrococcus Infection in Patients Receiving Epoprostenol by Continuous Infusion. Eur. J. Clin. Microbiol. Infect. Dis. 2003, 22, 704–705. [Google Scholar] [CrossRef]
- Hirata, Y.; Makiuchi, Y.; Sata, M.; Tomoike, H.; Wada, A.; Morikane, K.; Okabe, N. Comparative analysis of Micrococcus luteus isolates from blood cultures of patients with pulmonary hypertension receiving epoprostenol continuous infusion. J. Infect. Chemother. 2009, 15, 424–425. [Google Scholar] [CrossRef]
- Mermel, L.A. Prevention of intravascular catheter–related infections. Ann. Intern. Med. 2000, 132, 391–402. [Google Scholar] [CrossRef]
- Al-Wazni, W.S.; Hadi, B.S. Antivirulence effects of pomegranate peel extracts on most common urinary tract infection pathogens in pregnant women. J. Contemp. Med Sci. 2016, 1, 7. [Google Scholar] [CrossRef]
- Mayyas, A.; Abu-Sini, M.; Amr, R.; Akasheh, R.T.; Zalloum, W.; Khdair, A.; Hamad, I.; Aburjai, T.; Darwish, R.M.; Abu-Qatouseh, L. Novel in vitro and in vivo anti-Helicobacter pylori effects of pomegranate peel ethanol extract. Vet. World 2021, 14, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Chi, Y.; Zhang, S. The use of a tea polyphenol dip to extend the shelf life of silver carp (Hypophthalmicthys molitrix) during storage in ice. Food Chem. 2008, 108, 148–153. [Google Scholar] [CrossRef]
- Lei, Y.; Tang, Z.; Liao, R.; Guo, B. Hydrolysable tannin as environmentally friendly reducer and stabilizer for graphene oxide. Green Chem. 2011, 13, 1655–1658. [Google Scholar] [CrossRef]
- Tehranifar, A.; Selahvarzi, Y.; Kharrazi, M.; Bakhsh, V.J. High potential of agro-industrial by-products of pomegranate (Punica granatum L.) as the powerful antifungal and antioxidant substances. Ind. Crop. Prod. 2011, 34, 1523–1527. [Google Scholar] [CrossRef]
- Houston, D.M.; Bugert, J.; Denyer, S.P.; Heard, C.M. Anti-inflammatory activity of Punica granatum L. (Pomegranate) rind extracts applied topically to ex vivo skin. Eur. J. Pharm. Biopharm. 2017, 112, 30–37. [Google Scholar] [CrossRef]
- Celiksoy, V.; Moses, R.L.; Sloan, A.J.; Moseley, R.; Heard, C.M. Evaluation of the in vitro oral wound healing effects of pomegranate (Punica granatum) rind extract and punicalagin, in combination with Zn (II). Biomolecules 2020, 10, 1234. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, R.; Gung, B.W.; Tindall, S.; Gonzalez, J.M.; Halvorson, J.J.; Hagerman, A.E. Polyphenol–Aluminum Complex Formation: Implications for Aluminum Tolerance in Plants. J. Agric. Food Chem. 2016, 64, 3025–3033. [Google Scholar] [CrossRef]
- Lansdown, A.B.G.; Mirastschijski, U.; Stubbs, N.; Scanlon, E.; Ågren, M.S. Zinc in wound healing: Theoretical, experimental, and clinical aspects. Wound Repair Regen. 2007, 15, 2–16. [Google Scholar] [CrossRef]
- Ong, C.-L.Y.; Gillen, C.M.; Barnett, T.; Walker, M.J.; McEwan, A.G. An Antimicrobial Role for Zinc in Innate Immune Defense Against Group A Streptococcus. J. Infect. Dis. 2014, 209, 1500–1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | MIC (mg/L) | MBC (mg/L) |
|---|---|---|
| PRE | 1560 | >1790 |
| Zn (II) | 440 | >1790 |
| FICI (PRE) | FICI (Zn (II)) | FICI | Conclusion | |
|---|---|---|---|---|
| M. luteus | 0.0625 | 0.0625 | 0.125 | Synergy |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Celiksoy, V.; Moses, R.L.; Sloan, A.J.; Moseley, R.; Heard, C.M. Synergistic In Vitro Antimicrobial Activity of Pomegranate Rind Extract and Zinc (II) against Micrococcus luteus under Planktonic and Biofilm Conditions. Pharmaceutics 2021, 13, 851. https://doi.org/10.3390/pharmaceutics13060851
Celiksoy V, Moses RL, Sloan AJ, Moseley R, Heard CM. Synergistic In Vitro Antimicrobial Activity of Pomegranate Rind Extract and Zinc (II) against Micrococcus luteus under Planktonic and Biofilm Conditions. Pharmaceutics. 2021; 13(6):851. https://doi.org/10.3390/pharmaceutics13060851
Chicago/Turabian StyleCeliksoy, Vildan, Rachael L. Moses, Alastair J. Sloan, Ryan Moseley, and Charles M. Heard. 2021. "Synergistic In Vitro Antimicrobial Activity of Pomegranate Rind Extract and Zinc (II) against Micrococcus luteus under Planktonic and Biofilm Conditions" Pharmaceutics 13, no. 6: 851. https://doi.org/10.3390/pharmaceutics13060851