
Improved Functionality of Integration-Deficient Lentiviral Vectors (IDLVs) by the Inclusion of IS2 Protein Docks

 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Lentiviral Vector Constructs
2.3. Viral Production
2.4. Estimation of Viral Particles Available on the Packaging Cell Supernatant
2.5. Etoposide Exposure
2.6. CRISPR/Cas9 Gene Targeting
2.7. Flow Cytometry
2.8. Viral Vector Genome Estimation
2.9. Estimation of IDLV Efficiency
2.10. Statistical Analysis
3. Results
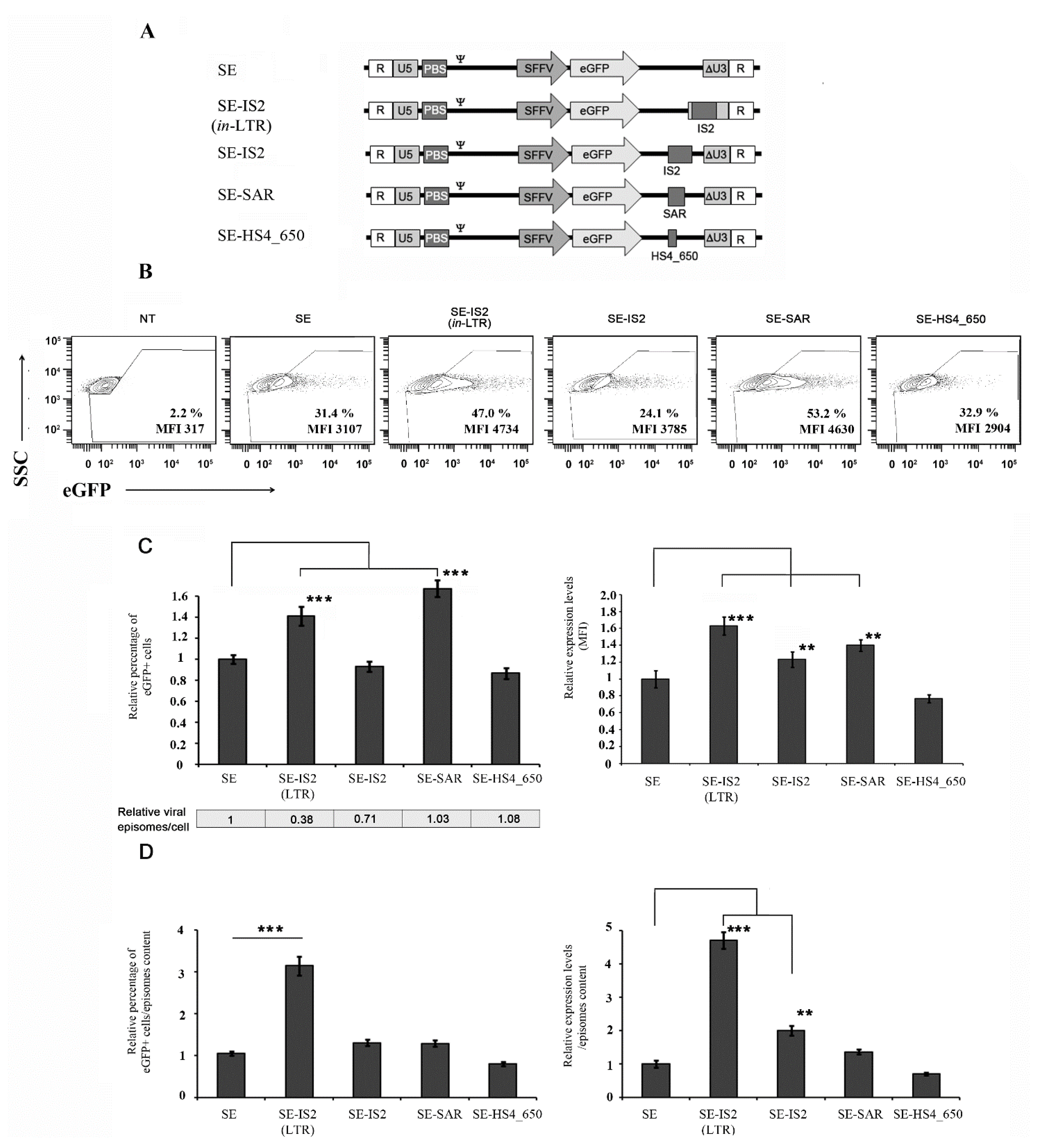
3.1. The Location and Composition of Chromatin Boundary Elements (CBEs) Greatly Affect IDLV Efficiency
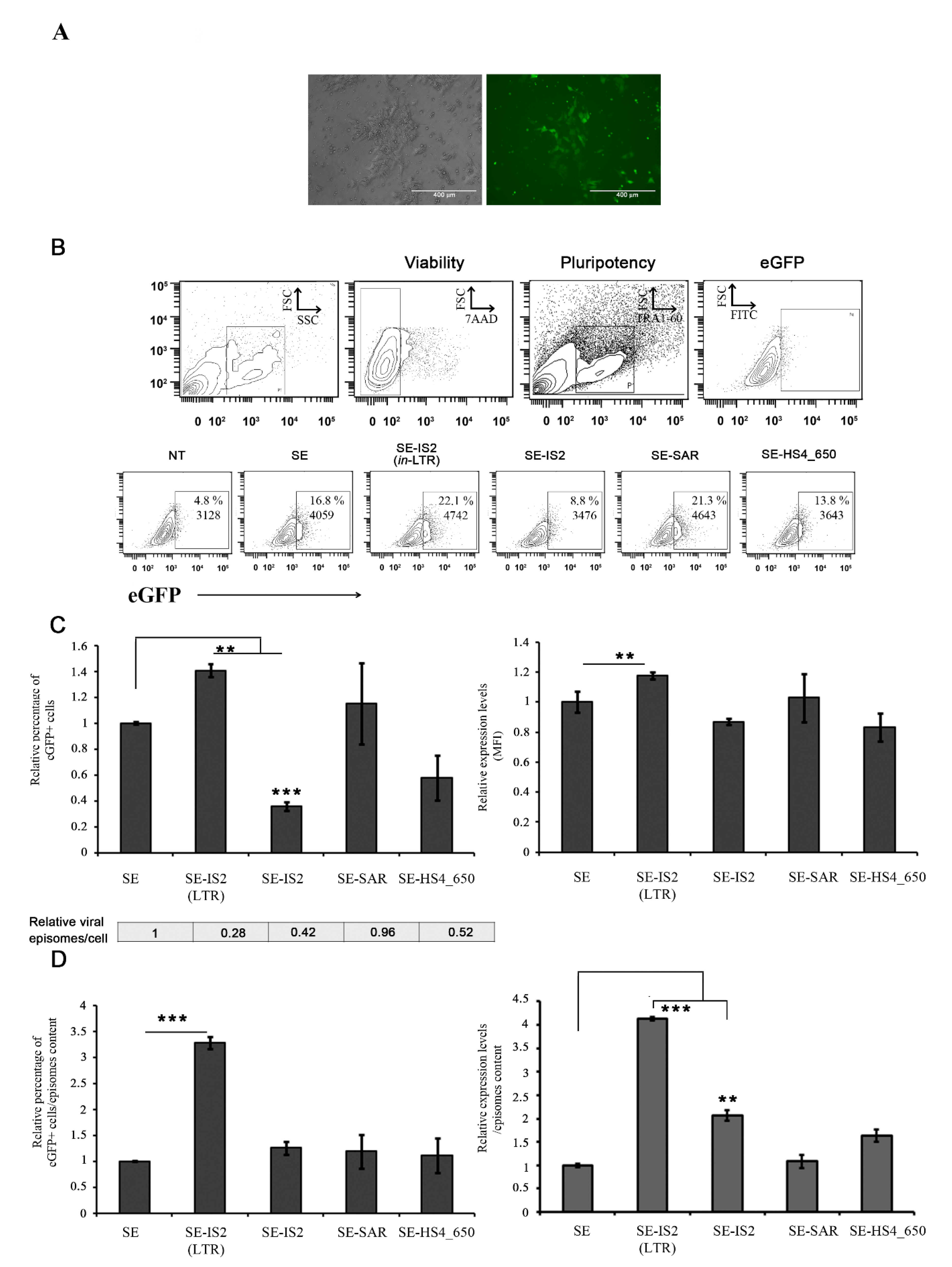
3.2. SE-IS2 (in-LTR) Enhances SE-IDLV Efficiency in Human Induced Pluripotent Stem Cells (hiPSCs)
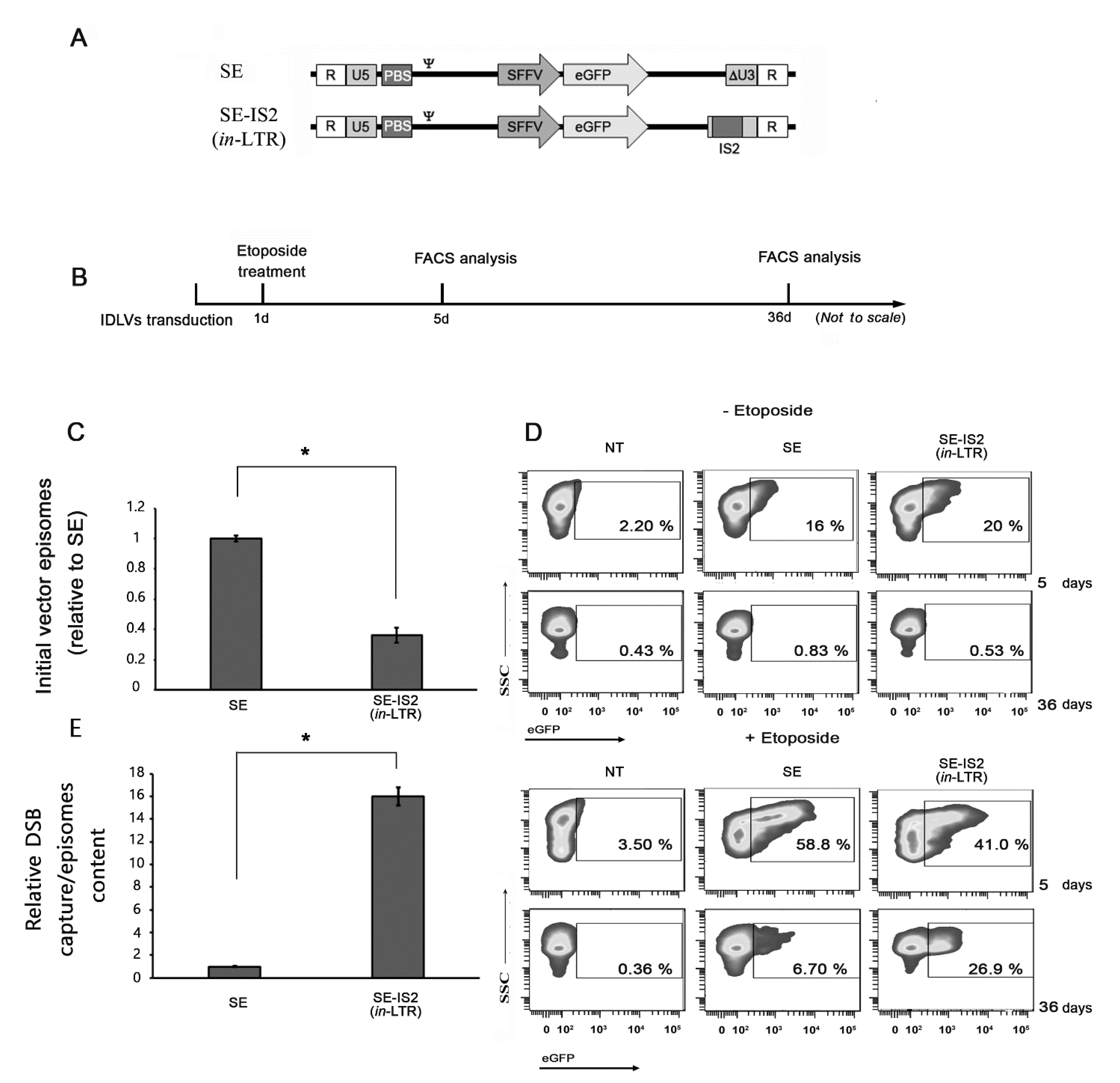
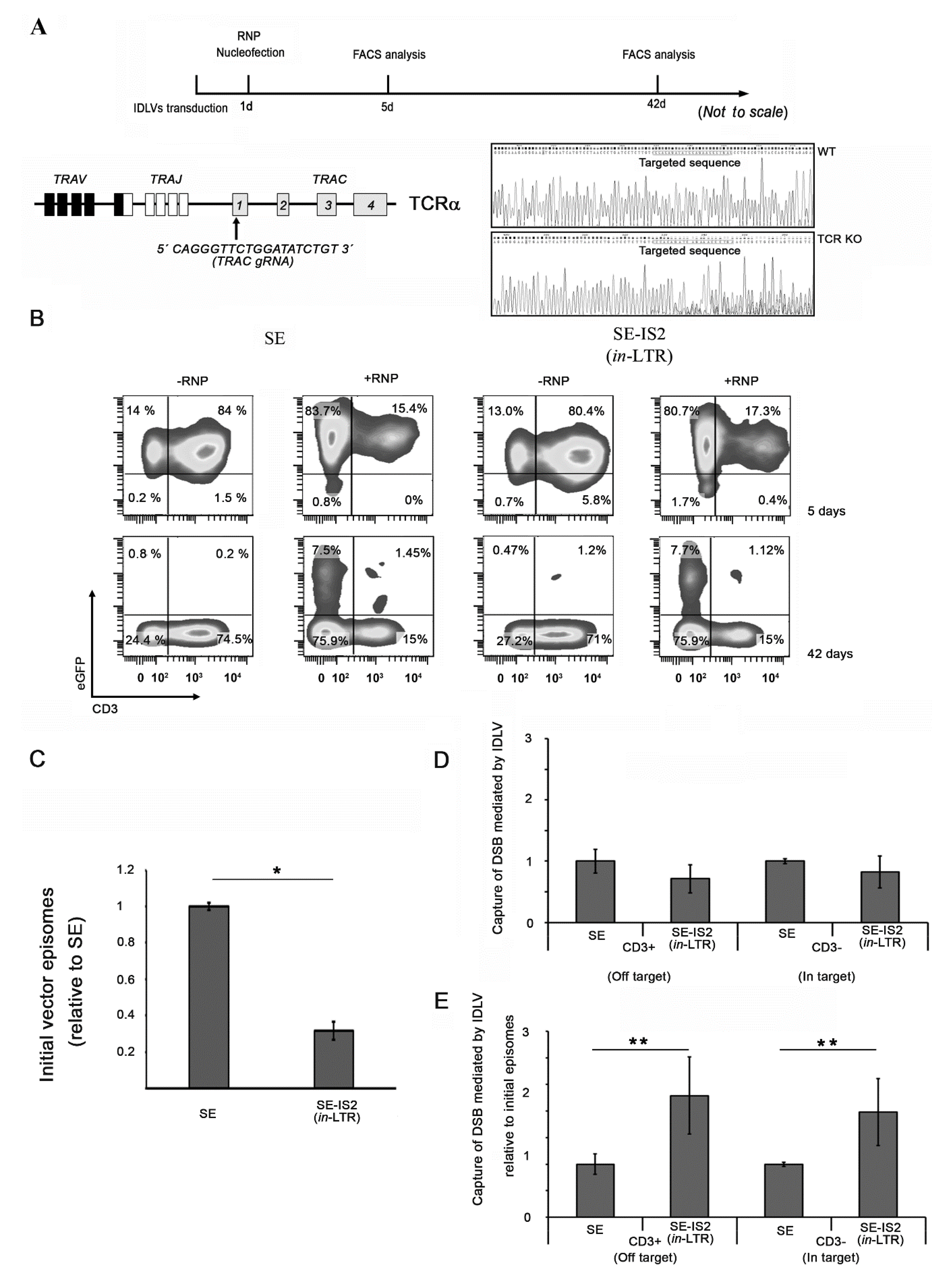
3.3. Inclusion of IS2 Increases the Number of IDLVs Captured by DNA Double-Strand Breaks (DSBs)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naldini, L. Gene therapy returns to centre stage. Nature 2015, 526, 351–360. [Google Scholar] [CrossRef]
- Dunbar, C.E.; High, K.A.; Joung, J.K.; Kohn, D.B.; Qzawa, K.; Sadelain, M. Gene therapy comes of age. Science 2018, 359, eaan4672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naldini, L. Genetic engineering of hematopoiesis: Current stage of clinical translation and future perspectives. EMBO Mol. Med. 2019, 11, e9958. [Google Scholar] [CrossRef] [PubMed]
- Porteus, M.H. A New Class of Medicines through DNA Editing. N. Engl. J. Med. 2019, 380, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Naldini, L. Ex Vivo gene transfer and correction for cell-based therapies. Nat. Rev. Genet. 2011, 12, 301–315. [Google Scholar]
- Ramamoorth, M.; Narvekar, A. Non viral vectors in gene therapy—An overview. J. Clin. Diagn. Res. JCDR 2015, 9, GE01-6. [Google Scholar] [CrossRef]
- Li, C.; Samulski, R.J. Engineering adeno-associated virus vectors for gene therapy. Nat. Rev. Genet. 2020, 21, 255–272. [Google Scholar] [CrossRef]
- Patel, A.; Zhao, J.; Duan, D.; Lai, Y. Design of AAV Vectors for Delivery of Large or Multiple Transgenes. Methods Mol. Biol. 2019, 1950, 19–33. [Google Scholar]
- Liu, K.C.; Lin, B.-S.; Gao, A.-D.; Ma, H.-Y.; Zhao, M.; Zhang, R.; Yan, H.-H.; Yi, X.-F.; Lin, S.-J.; Que, J.-W.; et al. Integrase-deficient lentivirus: Opportunities and challenges for human gene therapy. Curr. Gene Ther. 2014, 14, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Philippe, S.; Sarkis, C.; Barkats, M.; Mammeri, H.; Ladroue, C.; Petit, C.; Mallet, J.; Serguera, C. Lentiviral vectors with a defective integrase allow efficient and sustained transgene expression In Vitro and In Vivo. Proc. Natl. Acad. Sci. USA 2006, 103, 17684–17689. [Google Scholar] [CrossRef] [Green Version]
- Vargas, J., Jr.; Gusella, G.L.; Najfeld, V.; Klotman, M.E.; Cara, A. Novel integrase-defective lentiviral episomal vectors for gene transfer. Hum. Gene Ther. 2004, 15, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.; Cornetta, K. Design and Potential of Non-Integrating Lentiviral Vectors. Biomedicines 2014, 2, 14–35. [Google Scholar] [CrossRef] [Green Version]
- Kampik, D.; Basche, M.; Luhmann, U.F.O.; Nishiguchi, K.M.; Williams, J.A.E.; Greenwood, J.; Moss, S.E.; Han, H.; Azam, S.; Duran, Y.; et al. In situ regeneration of retinal pigment epithelium by gene transfer of E2F2: A potential strategy for treatment of macular degenerations. Gene Ther. 2017, 24, 810–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, V.Y.-C.; Ferreira, S.; Waddington, S.N.; Antoniou, M.N. Haemophilia B curative FIX production from a low dose UCOE-based lentiviral vector following hepatic pre-natal delivery. Curr. Gene Ther. 2016, 16, 231–241. [Google Scholar]
- Suwanmanee, T.; Hu, G.; Gui, T.; Bartholomae, C.C.; Kustchera, I.; von Kalle, C.; Schmidt, M.; Monahan, P.E.; Kafri, T. Integration-deficient lentiviral vectors expressing codon-optimized R338L human FIX restore normal hemostasis in Hemophilia B mice. Mol. Ther. J. Am. Soc. Gene Ther. 2014, 22, 567–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez-Guerrero, A.; Sanchez-Hernandez, S.; Galvani, G.; Pinedo-Gomez, J.; Martin-Guerra, R.; Sanchez-Gilabert, A.; Aguilar-González, A.; Cobo, M.; Gregory, P.; Holmes, M.; et al. Comparison of Zinc Finger Nucleases Versus CRISPR-Specific Nucleases for Genome Editing of the Wiskott-Aldrich Syndrome Locus. Hum. Gene Ther. 2018, 29, 366–380. [Google Scholar] [CrossRef] [PubMed]
- Ortinski, P.I.; O’Donovan, B.; Dong, X.; Kantor, B. Integrase-Deficient Lentiviral Vector as an All-in-One Platform for Highly Efficient CRISPR/Cas9-Mediated Gene Editing. Mol. Ther. Methods Clin. Dev. 2017, 5, 153–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, Y.; Chang, T.; Huang, H.; Yee, J.-K. Integration-defective lentiviral vector mediates efficient gene editing through homology-directed repair in human embryonic stem cells. Nucleic Acids Res. 2017, 45, e29. [Google Scholar] [CrossRef] [Green Version]
- Hoban, M.D.; Romero, Z.; Cost, G.J.; Mendel, M.; Holmes, M.; Kohn, D.B. Delivery of Genome Editing Reagents to Hematopoietic Stem/Progenitor Cells. Curr. Protoc. Stem Cell Biol. 2016, 36, 5B.4.1–5B.4.10. [Google Scholar] [CrossRef] [PubMed]
- Pelascini, L.P.L.; Maggio, I.; Lin, J.; Holkers, M.; Cathomen, T.; Gonçalves, M.A.F.V. Histone deacetylase inhibition rescues gene knockout levels achieved with integrase-defective lentiviral vectors encoding zinc-finger nucleases. Hum. Gene Ther. Methods 2013, 24, 399–411. [Google Scholar] [CrossRef] [Green Version]
- Joglekar, A.V.; Hollis, R.P.; Kuftinec, G.; Senadheera, S.; Chan, R.; Kohn, D.B. Integrase-defective lentiviral vectors as a delivery platform for targeted modification of adenosine deaminase locus. Mol. Ther. J. Am. Soc. Gene Ther. 2013, 21, 1705–1717. [Google Scholar] [CrossRef] [Green Version]
- Martin, F.; Sánchez-Hernández, S.; Gutiérrez-Guerrero, A.; Pinedo-Gomez, J.; Benabdellah, K. Biased and Unbiased Methods for the Detection of Off-Target Cleavage by CRISPR/Cas9: An Overview. Int. J. Mol. Sci. 2016, 17, 1507. [Google Scholar] [CrossRef]
- Gabriel, R.; Lombardo, A.; Arens, A.; Miller, J.C.; Genovese, P.; Kaeppel, C.; Nowrouzi, A.; Bartholomae, C.C.; Wang, J.; Friedman, G.; et al. An unbiased genome-wide analysis of zinc-finger nuclease specificity. Nat. Biotechnol. 2011, 29, 816–823. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Wu, X.; Wang, J.; Wang, Y.; Qiu, Z.; Chang, T.; Huang, H.; Lin, R.-J.; Yee, J.-K. Unbiased detection of off-target cleavage by CRISPR-Cas9 and TALENs using integrase-defective lentiviral vectors. Nat. Biotechnol. 2015, 33, 175–178. [Google Scholar] [CrossRef]
- Girod, P.A.; Zahn-Zabal, M.; Mermod, N. Use of the chicken lysozyme 5′ matrix attachment region to generate high producer CHO cell lines. Biotechnol. Bioeng. 2005, 91, 1–11. [Google Scholar] [CrossRef]
- Zhao, C.P.; Guo, X.; Chen, S.-J.; Li, C.-Z.; Yang, Y.; Zhang, J.-H.; Chen, S.-N.; Jia, Y.-L.; Wang, T.-Y. Matrix attachment region combinations increase transgene expression in transfected Chinese hamster ovary cells. Sci. Rep. 2017, 7, 42805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, R.; Martínez, I.; Petriz, J.; Nadal, M.; Tintoré, X.; Gonzalez, J.R.; Gratacós, E.; Aran, J.M. The beta-interferon scaffold attachment region confers high-level transgene expression and avoids extinction by epigenetic modifications of integrated provirus in adipose tissue-derived human mesenchymal stem cells. Tissue Eng. Part C Methods 2011, 17, 275–287. [Google Scholar] [CrossRef]
- Hagedorn, C.; Antoniou, M.N.; Lipps, H.J. Genomic cis-acting Sequences Improve Expression and Establishment of a Nonviral Vector. Mol. Ther. Nucleic Acids 2013, 2, e118. [Google Scholar] [CrossRef]
- Xue, Z.; Lv, X.; Song, W.; Wang, X.; Zhao, G.-N.; Wang, W.-T.; Xiong, J.; Mao, B.-B.; Yu, W.; Yang, B.; et al. SIRT1 deacetylates SATB1 to facilitate MAR HS2-MAR epsilon interaction and promote epsilon-globin expression. Nucleic Acids Res. 2012, 40, 4804–4815. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Zhang, X.; Wang, T.-Y.; Jia, Y.-L.; Xu, D.-H.; Yi, D.-D. Shortened nuclear matrix attachment regions are sufficient for replication and maintenance of episomes in mammalian cells. Mol. Biol. Cell 2019, 30, 2761–2770. [Google Scholar] [CrossRef] [PubMed]
- Yusufzai, T.M.; Felsenfeld, G. The 5′-HS4 chicken beta-globin insulator is a CTCF-dependent nuclear matrix-associated element. Proc. Natl. Acad. Sci. USA 2004, 101, 8620–8624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, X.-Y.; Jia, Y.-L.; Guo, X.; Wang, Y.-F.; Wang, T.-Y. A Vector Based on the Chicken Hypersensitive Site 4 Insulator Element Replicates Episomally in Mammalian Cells. Curr. Gene Ther. 2017, 16, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Liu, J.; Liu, Q.; Zheng, L.; Wang, Y.; Su, J.; Hu, G.; Zhang, Y. Chicken hypersensitive site-4 insulator increases human serum albumin expression in bovine mammary epithelial cells modified with phiC31 integrase. Biotechnol. Lett. 2013, 35, 529–537. [Google Scholar] [CrossRef]
- Arumugam, P.I.; Urbinati, F.; Velu, C.S.; Higashimoto, T.; Grimes, H.L.; Malik, P. The 3′ region of the chicken hypersensitive site-4 insulator has properties similar to its core and is required for full insulator activity. PLoS ONE 2009, 4, e6995. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, P.I.; Scholes, J.; Perelman, N.; Xia, P.; Yee, J.-K.; Malik, P. Improved human beta-globin expression from self-inactivating lentiviral vectors carrying the chicken hypersensitive site-4 (cHS4) insulator element. Mol. Ther. J. Am. Soc. Gene Ther. 2007, 15, 1863–1871. [Google Scholar] [CrossRef]
- Sanchez-Hernandez, S.; Gutierrez-Guerrero, A.; Martín-Guerra, R.; Cortijo-Gutierrez, M.; Tristán-Manzano, M.; Rodriguez-Perales, S.; Sanchez, L.; Garcia-Perez, J.L.; Chato-Astrain, J.; Fernandez-Valades, R.; et al. The IS2 Element Improves Transcription Efficiency of Integration-Deficient Lentiviral Vector Episomes. Mol. Ther. Nucleic Acids 2018, 13, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Benabdellah, K.; Muñoz, P.; Cobo, M.; Gutierrez-Guerrero, A.; Sánchez-Hernández, S.; Garcia-Perez, A.; Anderson, P.; Carrillo-Gálvez, A.B.; Toscano, M.G.; Martin, F. Lent-On-Plus Lentiviral vectors for conditional expression in human stem cells. Sci. Rep. 2016, 6, 37289. [Google Scholar] [CrossRef]
- Benabdellah, K.; Gutierrez-Guerrero, A.; Cobo, M.; Muñoz, P.; Martín, F. A chimeric HS4-SAR insulator (IS2) that prevents silencing and enhances expression of lentiviral vectors in pluripotent stem cells. PLoS ONE 2014, 9, e84268. [Google Scholar] [CrossRef]
- Urbinati, F.; Arumugam, P.; Higashimoto, T.; Perumbeti, A.; Mitts, K.; Xia, P.; Malik, P. Mechanism of reduction in titers from lentivirus vectors carrying large inserts in the 3′LTR. Mol. Ther. J. Am. Soc. Gene Ther. 2009, 17, 1527–1536. [Google Scholar] [CrossRef]
- Montes, R.; Romero, T.; Cabrera, S.; Ayllon, V.; Lopez-Escamez, J.A.; Ramos-Mejia, V.; Real, P.J. Generation and characterization of the human iPSC line PBMC1-iPS4F1 from adult peripheral blood mononuclear cells. Stem Cell Res. 2015, 15, 614–617. [Google Scholar] [CrossRef] [Green Version]
- Martin, F.; Toscano, M.G.; Blundell, M.; Frecha, C.; Srivastava, G.K.; Santamaría, M.; Thrasher, A.J.; Molina, I.J. Lentiviral vectors transcriptionally targeted to hematopoietic cells by WASP gene proximal promoter sequences. Gene Ther. 2005, 12, 715–723. [Google Scholar] [CrossRef]
- Pattanayak, V.; Ramirez, C.L.; Joung, J.K.; Liu, D.R. Revealing off-target cleavage specificities of zinc-finger nucleases by in vitro selection. Nat. Methods 2011, 8, 765–770. [Google Scholar] [CrossRef] [Green Version]
- Pattanayak, V.; Lin, S.; Guilinger, J.P.; Ma, E.; Doudna, J.A.; Liu, D.R. High-throughput profiling of off-target DNA cleavage reveals RNA-programmed Cas9 nuclease specificity. Nat. Biotechnol. 2013, 31, 839–843. [Google Scholar] [CrossRef]
- Zelensky, A.N.; Schoonakker, M.; Brandsma, I.; Tijsterman, M.; Van Gent, D.C.; Essers, J.; Kanaar, R. Low dose ionizing radiation strongly stimulates insertional mutagenesis in a γH2AX dependent manner. PLoS Genet. 2020, 16, e1008550. [Google Scholar] [CrossRef] [PubMed]
- Koyama, T.; Sun, B.; Tokunaga, K.; Tatsumi, M.; Ishizaka, Y. DNA damage enhances integration of HIV-1 into macrophages by overcoming integrase inhibition. Retrovirology 2013, 10, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, S.Q.; Zheng, Z.; Nguyen, N.T.; Liebers, M.; Topkar, V.V.; Thapar, V.; Wyvekens, N.; Khayter, C.; Iafrate, A.J.; Le, L.P.; et al. GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases. Nat. Biotechnol. 2015, 33, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Bae, S.; Park, J.; Kim, E.; Kim, S.; Yu, H.R.; Hwang, J.; Kim, J.-I.; Kim, J.-S. Digenome-seq: Genome-wide profiling of CRISPR-Cas9 off-target effects in human cells. Nat. Methods 2015, 12, 237–243. [Google Scholar] [CrossRef]
- Tanwar, V.S.; Jose, C.C.; Cuddapah, S. Role of CTCF in DNA damage response. Mutat. Res. 2019, 780, 61–68. [Google Scholar] [CrossRef]
- Hilmi, K.; Jangal, M.; Marques, M.; Zhao, T.; Saad, A.; Zhang, C.; Luo, V.M.; Syme, A.; Rejon, C.; Yu, Z.; et al. CTCF facilitates DNA double-strand break repair by enhancing homologous recombination repair. Sci. Adv. 2017, 3, e1601898. [Google Scholar] [CrossRef] [Green Version]
- Natale, F.; Rapp, A.; Yu, W.; Maiser, A.; Harz, H.; Scholl, A.; Grulich, S.; Anton, T.; Hörl, D.; Chen, W.; et al. Identification of the elementary structural units of the DNA damage response. Nat. Commun. 2017, 8, 15760. [Google Scholar] [CrossRef] [PubMed]
- Ginn, S.L.; Amaya, A.K.; Alexander, I.E.; Edelstein, M.; Abedi, M.R. Gene therapy clinical trials worldwide to 2017: An update. J. Gene Med. 2018, 20, e3015. [Google Scholar] [CrossRef] [PubMed]
- Peluffo, H.; Foster, E.; Ahmed, S.G.; Lago, N.; Hutson, T.H.; Moon, L.; Yip, P.; Wanisch, K.; Caraballo-Miralles, V.; Olmos, G.; et al. Efficient gene expression from integration-deficient lentiviral vectors in the spinal cord. Gene Ther. 2012, 20, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Naldini, L.; Blömer, U.; Gallay, P.; Ory, D.; Mulligan, R.; Gage, F.H.; Verma, I.M.; Trono, D. In Vivo Gene Delivery and Stable Transduction of Nondividing Cells by a Lentiviral Vector. Science 1996, 272, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelascini, L.P.; Janssen, J.M.; Gonçalves, M.A. Histone Deacetylase Inhibition Activates Transgene Expression from Integration-Defective Lentiviral Vectors in Dividing and Non-Dividing Cells. Hum. Gene Ther. 2013, 24, 78–96. [Google Scholar] [CrossRef] [PubMed]
- Knipe, D.M.; Lieberman, P.M.; Jung, J.U.; McBride, A.A.; Morris, K.; Ott, M.; Margolis, D.; Nieto, A.; Nevels, M.; Parks, R.J.; et al. Snapshots: Chromatin control of viral infection. Virology 2013, 435, 141–156. [Google Scholar] [CrossRef]
- Browning, D.L.; Everson, E.M.; Leap, D.J.; Hocum, J.D.; Wang, H.; Stamatoyannopoulos, G.; Trobridge, G.D. Evidence for the in vivo safety of insulated foamy viral vectors. Gene Ther. 2016, 24, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Narwade, N.; Patel, S.; Alam, A.; Chattopadhyay, S.; Mittal, S.; Kulkarni, A. Mapping of scaffold/matrix attachment regions in human genome: A data mining exercise. Nucleic Acids Res. 2019, 47, 7247–7261. [Google Scholar] [CrossRef] [Green Version]
- Maksimenko, O.; Gasanov, N.B.; Georgiev, P. Regulatory Elements in Vectors for Efficient Generation of Cell Lines Producing Target Proteins. Acta Nat. 2015, 7, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Osborn, M.J.; Webber, B.R.; Knipping, F.; Lonetree, C.-L.; Tennis, N.; DeFeo, A.P.; McElroy, A.N.; Starker, C.; Lee, C.; Merkel, S.; et al. Evaluation of TCR Gene Editing Achieved by TALENs, CRISPR/Cas9, and megaTAL Nucleases. Mol. Ther. 2016, 24, 570–581. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.-S.; Gong, S.; Yu, H.-H.; Jung, K.; Johnson, K.A.; Taylor, D.W. Engineered CRISPR/Cas9 enzymes improve discrimination by slowing DNA cleavage to allow release of off-target DNA. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Zhang, L.; Rube, H.T.; Vakulskas, C.A.; Behlke, M.A.; Bussemaker, H.J.; Pufall, M.A. Systematic in vitro profiling of off-target affinity, cleavage and efficiency for CRISPR enzymes. Nucleic Acids Res. 2020, 48, 5037–5053. [Google Scholar] [CrossRef] [PubMed]
- Vakulskas, C.A.; Behlke, M.A. Evaluation and Reduction of CRISPR Off-Target Cleavage Events. Nucleic Acid Ther. 2019, 29, 167–174. [Google Scholar] [CrossRef]
- Cornu, T.I.; Cathomen, T. Targeted Genome Modifications Using Integrase-deficient Lentiviral Vectors. Mol. Ther. 2007, 15, 2107–2113. [Google Scholar] [CrossRef] [PubMed]
- Kantor, B.; Bayer, M.; Ma, H.; Samulski, J.; Li, C.; McCown, T.J.; Kafri, T. Notable Reduction in Illegitimate Integration Mediated by a PPT-deleted, Nonintegrating Lentiviral Vector. Mol. Ther. 2011, 19, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Apolonia, L.; Waddington, S.; Fernandes, C.; Ward, N.J.; Bouma, G.; Blundell, M.; Thrasher, A.J.; Collins, M.K.; Philpott, N.J. Stable Gene Transfer to Muscle Using Non-integrating Lentiviral Vectors. Mol. Ther. 2007, 15, 1947–1954. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortijo-Gutiérrez, M.; Sánchez-Hernández, S.; Tristán-Manzano, M.; Maldonado-Pérez, N.; Lopez-Onieva, L.; Real, P.J.; Herrera, C.; Marchal, J.A.; Martin, F.; Benabdellah, K. Improved Functionality of Integration-Deficient Lentiviral Vectors (IDLVs) by the Inclusion of IS2 Protein Docks. Pharmaceutics 2021, 13, 1217. https://doi.org/10.3390/pharmaceutics13081217
Cortijo-Gutiérrez M, Sánchez-Hernández S, Tristán-Manzano M, Maldonado-Pérez N, Lopez-Onieva L, Real PJ, Herrera C, Marchal JA, Martin F, Benabdellah K. Improved Functionality of Integration-Deficient Lentiviral Vectors (IDLVs) by the Inclusion of IS2 Protein Docks. Pharmaceutics. 2021; 13(8):1217. https://doi.org/10.3390/pharmaceutics13081217
Chicago/Turabian StyleCortijo-Gutiérrez, Marina, Sabina Sánchez-Hernández, María Tristán-Manzano, Noelia Maldonado-Pérez, Lourdes Lopez-Onieva, Pedro J. Real, Concha Herrera, Juan Antonio Marchal, Francisco Martin, and Karim Benabdellah. 2021. "Improved Functionality of Integration-Deficient Lentiviral Vectors (IDLVs) by the Inclusion of IS2 Protein Docks" Pharmaceutics 13, no. 8: 1217. https://doi.org/10.3390/pharmaceutics13081217