
Acetylsalicylic Acid Suppresses Alcoholism-Induced Cognitive Impairment Associated with Atorvastatin Intake by Targeting Cerebral miRNA155 and NLRP3: In Vivo, and In Silico Study

 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Grouping
2.2. Chemical Reagents and Drugs
2.3. Induction of Alcoholic Brain Injury
2.4. Cognitive Function Assessment; Morris Water Maze (MWM)
2.5. Biochemical Analysis
Real-Time PCR
2.6. Histology Study
- Light microscopic (LM) analysis
- ii.
- Semi-thin sections analysis
- iii.
- Transmission electron microscopic (TEM) analysis
2.7. Morphometric Study
2.8. In Silico Molecular Modelling Study
2.9. Statistical Analysis
3. Results
3.1. Effect of Alcohol and/or ATOR Treatment on Cognitive Function and Assessment of ASA Administration
3.2. Biochemical Analysis
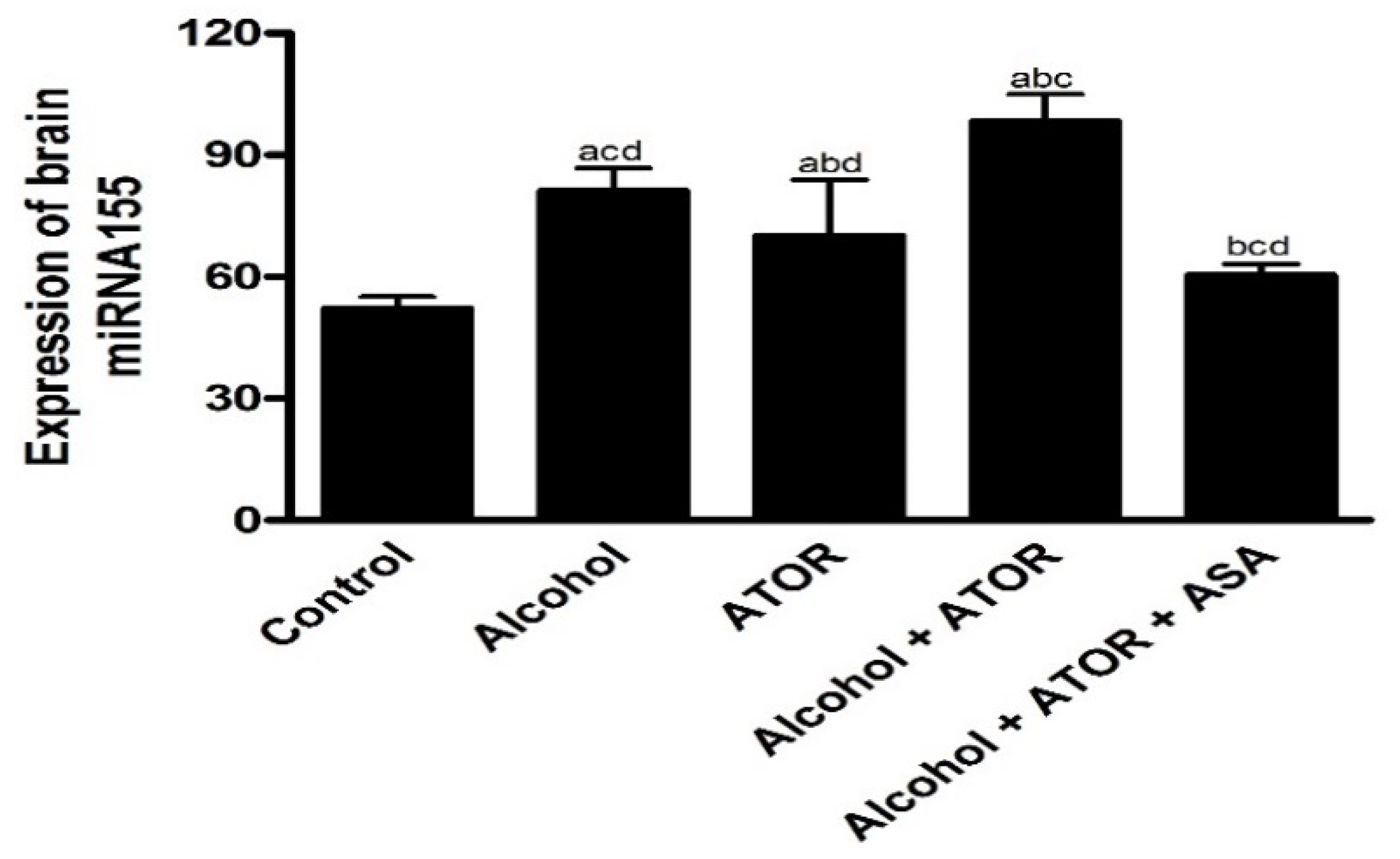
3.2.1. Effect of Alcohol and/or ATOR Treatment on miRNA155 Expression and Influence of ASA Administration
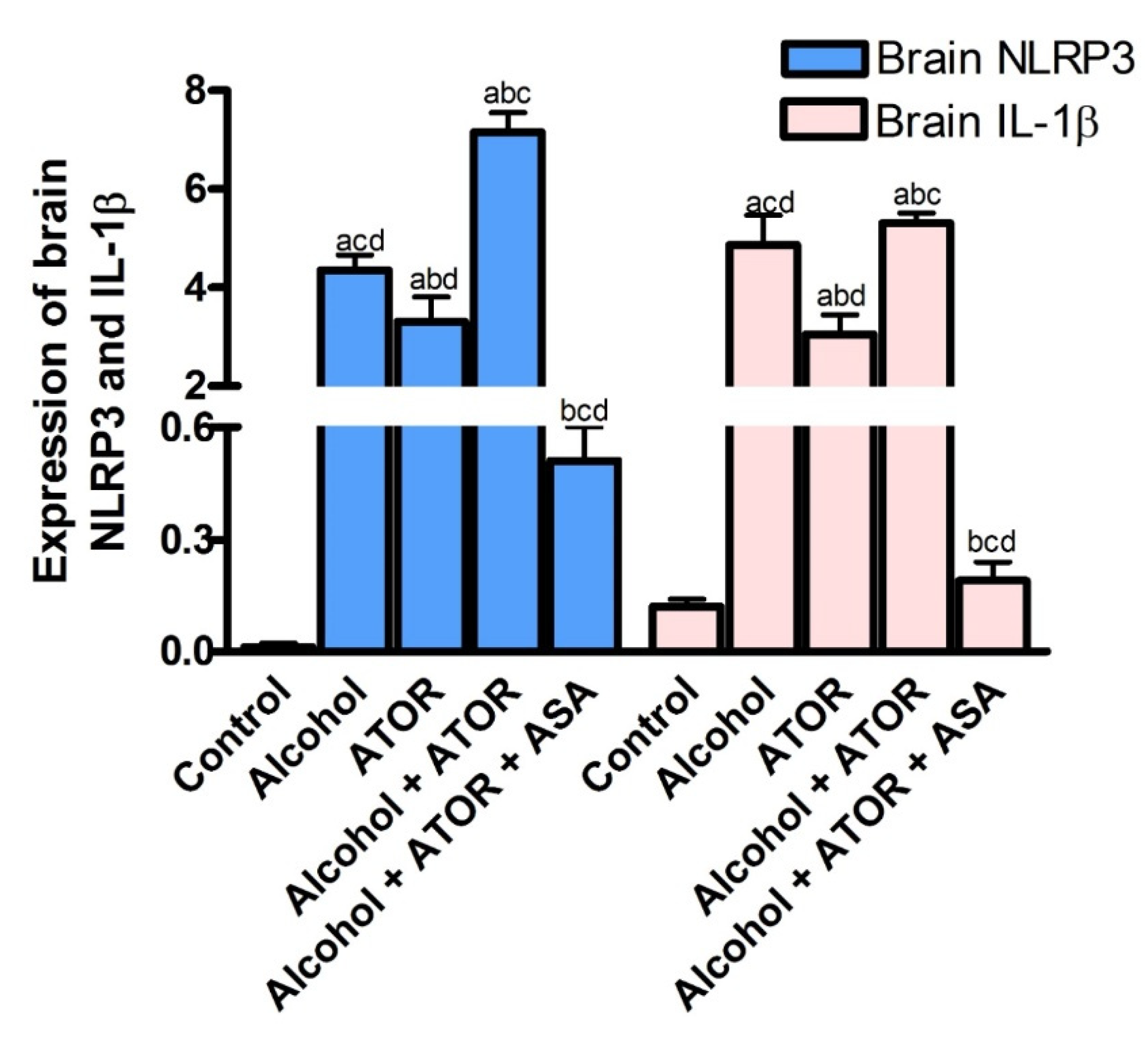
3.2.2. Effect of ASA on Brain NLRP3 and IL-1β Expression after Alcohol and/or ATOR Treatment
3.3. Histological Results
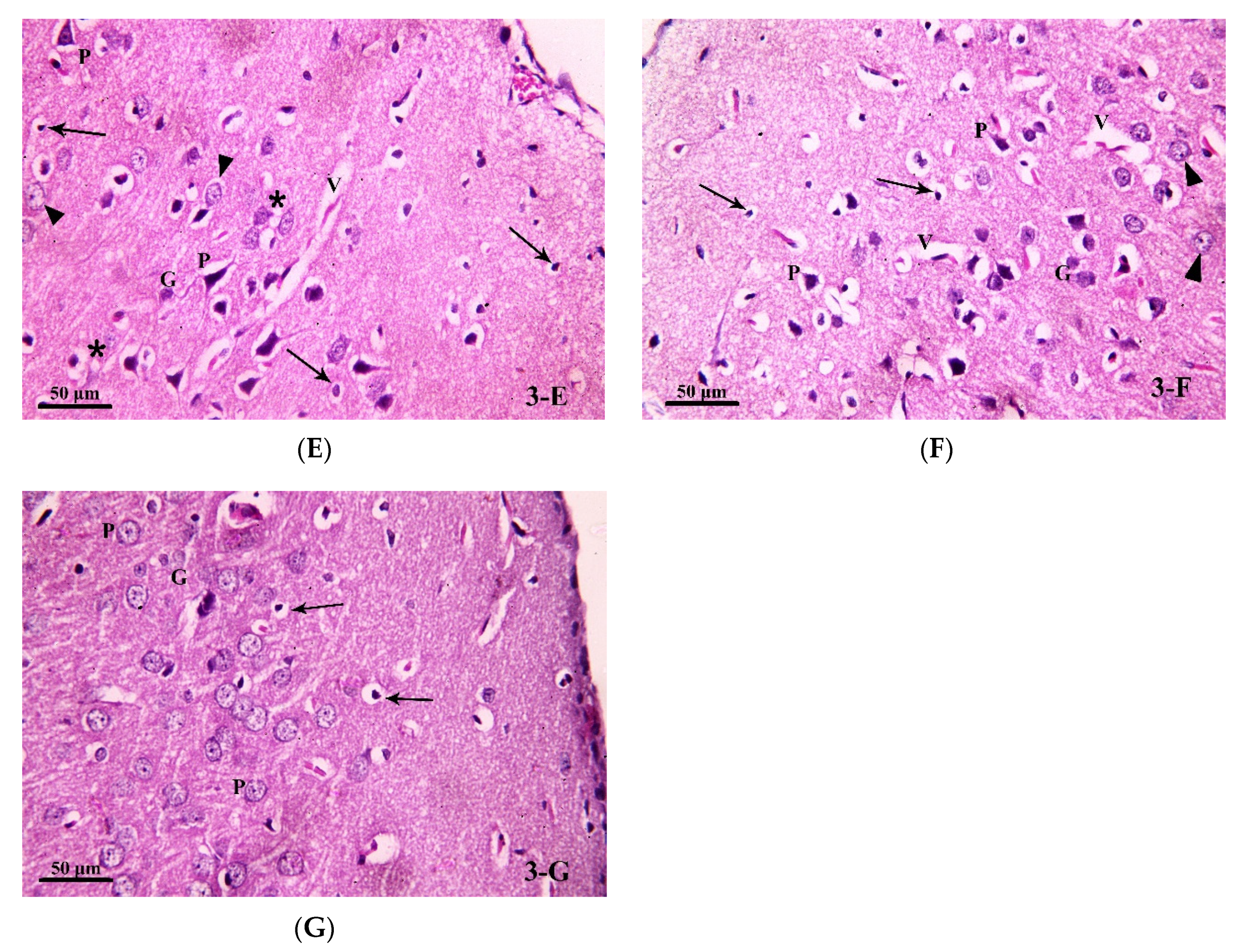
3.3.1. Light Microscopic Analysis
3.3.2. Caspase-8 Immunostained Analysis
3.3.3. Semi-Thin Sections Analysis
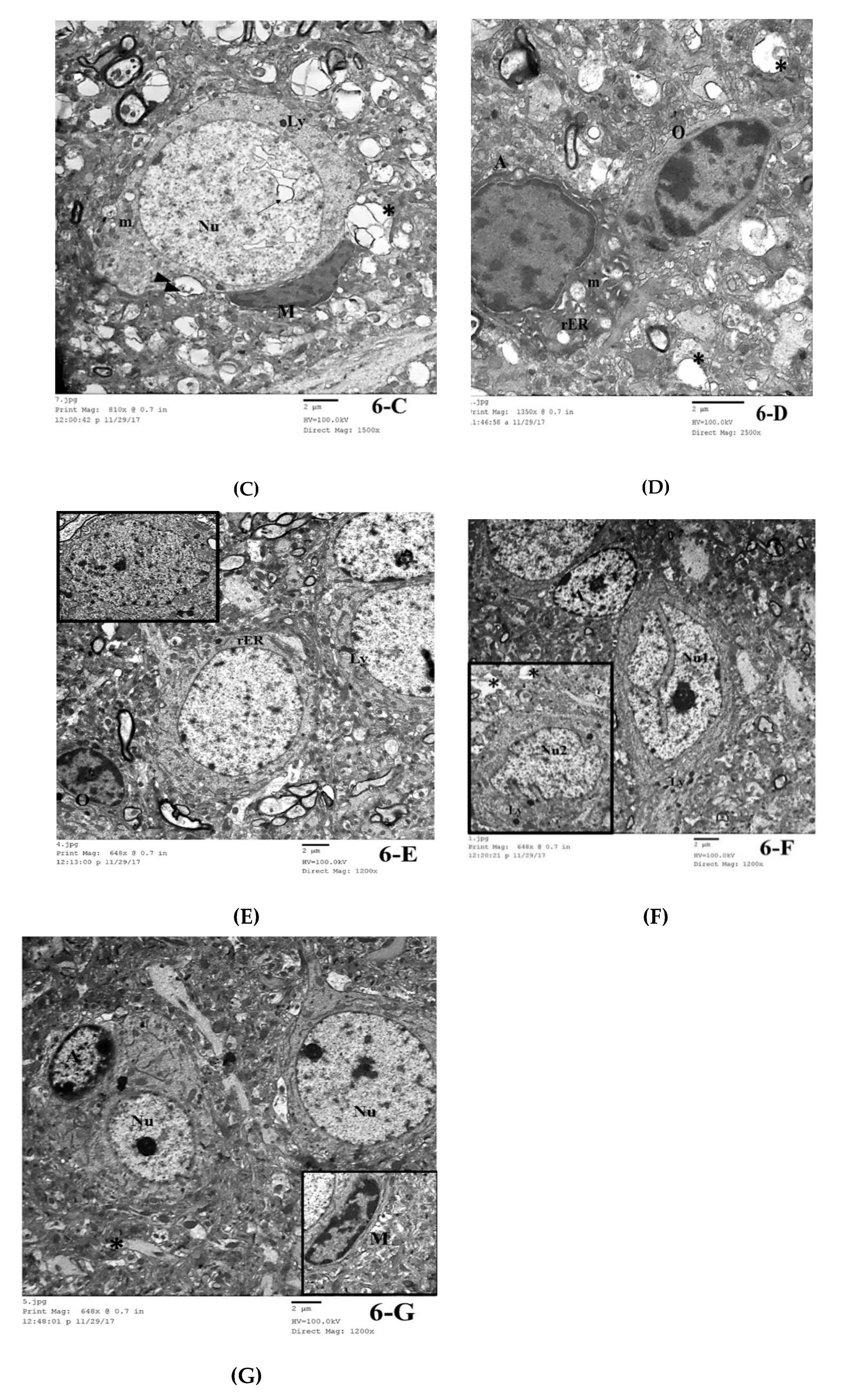
3.3.4. Transmission Electron Microscopic (TEM) Study
3.4. In Silico Molecular Docking Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ishii, T.; Hashimoto, E.; Ukai, W.; Tateno, M.; Yoshinaga, T.; Ono, T.; Watanabe, K.; Saito, S.; Saito, T. Epigenetic regulation in alcohol-related brain damage. Nihon Arukoru Yakubutsu Igakkai Zasshi 2008, 43, 705–713. [Google Scholar]
- Bajaj, J.S. Alcohol, Liver Disease and the Gut Microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Piano, M.R. Alcohol’s Effects on the Cardiovascular System. Alcohol. Res. 2017, 38, 219–241. [Google Scholar] [PubMed]
- Moss, M.; Burnham, E.L. Chronic Alcohol Abuse, Acute Respiratory Distress Syndrome, and Multiple Organ Dysfunction. Crit. Care Med. 2003, 31, S207–S212. [Google Scholar] [CrossRef]
- Osna, N.A.; Donohue, T.M.; Kharbanda, K.K. Alcoholic Liver Disease: Pathogenesis and Current Management. Alcohol. Res. 2017, 38, 147–161. [Google Scholar] [PubMed]
- Crews, F.T. Alcohol-Related Neurodegeneration and Recovery. Alcohol. Res. Health 2008, 31, 377–388. [Google Scholar]
- Friedman, N.P.; Robbins, T.W. The Role of Prefrontal Cortex in Cognitive Control and Executive Function. Neuropsychopharmacology 2022, 47, 72–89. [Google Scholar] [CrossRef] [PubMed]
- Vargas, J.I.; Arrese, M.; Shah, V.H.; Arab, J.P. Use of Statins in Patients with Chronic Liver Disease and Cirrhosis: Current Views and Prospects. Curr. Gastroenterol. Rep. 2017, 19, 43. [Google Scholar] [CrossRef]
- Gu, Y.; Yang, X.; Liang, H.; Li, D. Comprehensive Evaluation of Effects and Safety of Statin on the Progression of Liver Cirrhosis: A Systematic Review and Meta-Analysis. BMC Gastroenterol. 2019, 19, 231. [Google Scholar] [CrossRef] [Green Version]
- Ward, N.C.; Watts, G.F.; Eckel, R.H. Statin Toxicity. Circ. Res. 2019, 124, 328–350. [Google Scholar] [CrossRef]
- Yang, Q.; Liu, R.; Yu, Q.; Bi, Y.; Liu, G. Metabolic Regulation of Inflammasomes in Inflammation. Immunology 2019, 157, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Callaway, J.B.; Ting, J.P.-Y. Inflammasomes: Mechanism of Action, Role in Disease, and Therapeutics. Nat. Med. 2015, 21, 677–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesan, J.T.; Girnary, M.S.; Moss, K.; Monaghan, E.T.; Egnatz, G.J.; Jiao, Y.; Zhang, S.; Beck, J.; Swanson, K.V. Role of Inflammasomes in the Pathogenesis of Periodontal Disease and Therapeutics. Periodontology 2020 2020, 82, 93–114. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; de Alba, E. Structure, Activation and Regulation of NLRP3 and AIM2 Inflammasomes. Int. J. Mol. Sci. 2021, 22, 872. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.; Beane-Ebel, J.E.; Tanaka, Y.; Ning, B.; Husted, C.R.; Henderson, D.C.; Xiang, Y.; Park, I.-H.; Farrer, L.A.; Zhang, H. Exploration of Alcohol Use Disorder-Associated Brain MiRNA–MRNA Regulatory Networks. Transl. Psychiatry 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Tapocik, J.D.; Solomon, M.; Flanigan, M.; Meinhardt, M.; Barbier, E.; Schank, J.; Schwandt, M.; Sommer, W.H.; Heilig, M. Coordinated Dysregulation of MRNAs and MicroRNAs in the Rat Medial Prefrontal Cortex Following a History of Alcohol Dependence. Pharm. J. 2013, 13, 286–296. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Zhao, D.; Zhao, Y.; Wang, Y.; Zhao, Y.; Wen, C. Inhibition of MicroRNA-155 Alleviates Cognitive Impairment in Alzheimer’s Disease and Involvement of Neuroinflammation. Curr. Alzheimer Res. 2019, 16, 473–482. [Google Scholar] [CrossRef]
- Obora, K.; Onodera, Y.; Takehara, T.; Frampton, J.; Hasei, J.; Ozaki, T.; Teramura, T.; Fukuda, K. Inflammation-Induced MiRNA-155 Inhibits Self-Renewal of Neural Stem Cells via Suppression of CCAAT/Enhancer Binding Protein β (C/EBPβ) Expression. Sci. Rep. 2017, 7, 43604. [Google Scholar] [CrossRef] [Green Version]
- Slota, J.A.; Booth, S.A. MicroRNAs in Neuroinflammation: Implications in Disease Pathogenesis, Biomarker Discovery and Therapeutic Applications. Noncoding RNA 2019, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Lippai, D.; Bala, S.; Petrasek, J.; Csak, T.; Levin, I.; Kurt-Jones, E.A.; Szabo, G. Alcohol-Induced IL-1β in the Brain Is Mediated by NLRP3/ASC Inflammasome Activation That Amplifies Neuroinflammation. J. Leukoc. Biol. 2013, 94, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimons, S.; Oggero, S.; Bruen, R.; McCarthy, C.; Strowitzki, M.J.; Mahon, N.G.; Ryan, N.; Brennan, E.P.; Barry, M.; Perretti, M.; et al. MicroRNA-155 Is Decreased During Atherosclerosis Regression and Is Increased in Urinary Extracellular Vesicles During Atherosclerosis Progression. Front. Immunol. 2020, 11, 576516. [Google Scholar] [CrossRef] [PubMed]
- Parsamanesh, N.; Moossavi, M.; Bahrami, A.; Fereidouni, M.; Barreto, G.; Sahebkar, A. NLRP3 Inflammasome as a Treatment Target in Atherosclerosis: A Focus on Statin Therapy. Int. Immunopharmacol. 2019, 73, 146–155. [Google Scholar] [CrossRef]
- Tabaei, S.; Tabaee, S.S. Implications for MicroRNA Involvement in the Prognosis and Treatment of Atherosclerosis. Mol. Cell Biochem. 2021, 476, 1327–1336. [Google Scholar] [CrossRef] [PubMed]
- Schultz, B.G.; Patten, D.K.; Berlau, D.J. The Role of Statins in Both Cognitive Impairment and Protection against Dementia: A Tale of Two Mechanisms. Transl. Neurodegener 2018, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wu, Y.; Ye, L.; Wang, Y.; Zhang, K.; Wang, L.; Huang, Y.; Wang, L.; Xian, S.; Zhang, Y.; et al. Aspirin Alleviates Endothelial Gap Junction Dysfunction through Inhibition of NLRP3 Inflammasome Activation in LPS-Induced Vascular Injury. Acta Pharm. Sin. B 2019, 9, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Veronese, N.; Stubbs, B.; Maggi, S.; Thompson, T.; Schofield, P.; Muller, C.; Tseng, P.-T.; Lin, P.-Y.; Carvalho, A.F.; Solmi, M. Low-Dose Aspirin Use and Cognitive Function in Older Age: A Systematic Review and Meta-Analysis. J. Am. Geriatr. Soc. 2017, 65, 1763–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Kharashi, O.A.; El-Din Aly El-Waseef, D.A.; Nabih, E.S.; Mohamed, D.I. Targeting NLRP3 Inflammasome via Acetylsalicylic Acid: Role in Suppressing Hepatic Dysfunction and Insulin Resistance Induced by Atorvastatin in Naïve versus Alcoholic Liver in Rats. Biomed. Pharmacother. 2018, 107, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.D.; Antoine, D.J.; Shaw, P.J.; Benson, C.; Farhood, A.; Williams, D.P.; Kanneganti, T.-D.; Park, B.K.; Jaeschke, H. Role of the Nalp3 Inflammasome in Acetaminophen-Induced Sterile Inflammation and Liver Injury. Toxicol. Appl. Pharmacol. 2011, 252, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, V.; Chopra, K. Resveratrol Abrogates Alcohol-Induced Cognitive Deficits by Attenuating Oxidative-Nitrosative Stress and Inflammatory Cascade in the Adult Rat Brain. Neurochem. Int. 2013, 62, 861–869. [Google Scholar] [CrossRef]
- Solmaz, V.; Atasoy, Ö.; Erbaş, O. Atorvastatin Has Therapeutic Potential for the Fatty Liver-Induced Memory Dysfunction in Rats, Likely via Its Antioxidant and Anti-Inflammatory Properties. Neurol. Res. 2020, 42, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Melethil, S.; Mason, W.D. The Pharmacokinetics of Aspirin in Rats and the Effect of Buffer. J. Pharmacokinet. Biopharm. 1991, 19, 157–173. [Google Scholar] [CrossRef]
- Mischinger, H.J.; Walsh, T.R.; Liu, T.; Rao, P.N.; Rubin, R.; Nakamura, K.; Todo, S.; Starzl, T.E. An Improved Technique for Isolated Perfusion of Rat Livers and an Evaluation of Perfusates. J. Surg. Res. 1992, 53, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis Model Assessment: Insulin Resistance and β-Cell Function from Fasting Plasma Glucose and Insulin Concentrations in Man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, V.; Chopra, K. Protective Effect of Curcumin against Chronic Alcohol-Induced Cognitive Deficits and Neuroinflammation in the Adult Rat Brain. Neuroscience 2013, 244, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Khedr, S.A.; Elmelgy, A.A.; El-Kharashi, O.A.; Abd-Alkhalek, H.A.; Louka, M.L.; Sallam, H.A.; Aboul-Fotouh, S. Metformin Potentiates Cognitive and Antidepressant Effects of Fluoxetine in Rats Exposed to Chronic Restraint Stress and High Fat Diet: Potential Involvement of Hippocampal c-Jun Repression. Naunyn-Schmiedebergs. Arch. Pharmacol. 2018, 391, 407–422. [Google Scholar] [CrossRef]
- García-Cabezas, M.Á.; John, Y.J.; Barbas, H.; Zikopoulos, B. Distinction of Neurons, Glia and Endothelial Cells in the Cerebral Cortex: An Algorithm Based on Cytological Features. Front. Neuroanat. 2016, 10, 107. [Google Scholar] [CrossRef] [Green Version]
- Seligson, D.; Marino, J.; Dodson, E. Determination of Sulfobromophthalein in Serum. Clin. Chem. 1957, 3, 638–645. [Google Scholar] [CrossRef]
- Dekker, C.; Mattes, H.; Wright, M.; Boettcher, A.; Hinniger, A.; Hughes, N.; Kapps-Fouthier, S.; Eder, J.; Erbel, P.; Stiefl, N.; et al. Crystal Structure of NLRP3 NACHT Domain With an Inhibitor Defines Mechanism of Inflammasome Inhibition. J. Mol. Biol. 2021, 433, 167309. [Google Scholar] [CrossRef]
- Sharif, H.; Wang, L.; Wang, W.L.; Magupalli, V.G.; Andreeva, L.; Qiao, Q.; Hauenstein, A.V.; Wu, Z.; Núñez, G.; Mao, Y.; et al. Structural Mechanism for NEK7-Licensed Activation of NLRP3 Inflammasome. Nature 2019, 570, 338–343. [Google Scholar] [CrossRef]
- El Azab, I.H.; Saied, E.M.; Osman, A.A.; Mehana, A.E.; Saad, H.A.; Elkanzi, N.A. Novel N-Bridged Pyrazole-1-Carbothioamides with Potential Antiproliferative Activity: Design, Synthesis, in Vitro and in Silico Studies. Future Med. Chem. 2021, 13, 1743–1766. [Google Scholar] [CrossRef]
- Gaber, A.; Alsanie, W.F.; Kumar, D.N.; Refat, M.S.; Saied, E.M. Novel Papaverine Metal Complexes with Potential Anticancer Activities. Molecules 2020, 25, 5447. [Google Scholar] [CrossRef] [PubMed]
- Gaber, A.; Refat, M.S.; Belal, A.A.M.; El-Deen, I.M.; Hassan, N.; Zakaria, R.; Alhomrani, M.; Alamri, A.S.; Alsanie, W.F.; Saied, E.M. New Mononuclear and Binuclear Cu(II), Co(II), Ni(II), and Zn(II) Thiosemicarbazone Complexes with Potential Biological Activity: Antimicrobial and Molecular Docking Study. Molecules 2021, 26, 2288. [Google Scholar] [CrossRef] [PubMed]
- Healey, R.D.; Saied, E.M.; Cong, X.; Karsai, G.; Gabellier, L.; Saint-Paul, J.; Del Nero, E.; Jeannot, S.; Drapeau, M.; Fontanel, S.; et al. Discovery and Mechanism of Action of Small Molecule Inhibitors of Ceramidases**. Angew. Chem. 2022, 134, e202109967. [Google Scholar] [CrossRef]
- Mohamed, D.I.; Abou-Bakr, D.A.; Ezzat, S.F.; El-Kareem, H.F.A.; Nahas, H.H.A.; Saad, H.A.; Mehana, A.E.; Saied, E.M. Vitamin D3 Prevents the Deleterious Effects of Testicular Torsion on Testis by Targeting MiRNA-145 and ADAM17: In Silico and In Vivo Study. Pharmaceuticals 2021, 14, 1222. [Google Scholar] [CrossRef]
- Saied, E.M.; El-Maradny, Y.A.; Osman, A.A.; Darwish, A.M.G.; Abo Nahas, H.H.; Niedbała, G.; Piekutowska, M.; Abdel-Rahman, M.A.; Balbool, B.A.; Abdel-Azeem, A.M. A Comprehensive Review about the Molecular Structure of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Insights into Natural Products against COVID-19. Pharmaceutics 2021, 13, 1759. [Google Scholar] [CrossRef]
- Sliwoski, G.; Kothiwale, S.; Meiler, J.; Lowe, E.W. Computational Methods in Drug Discovery. Pharmacol. Rev. 2014, 66, 334–395. [Google Scholar] [CrossRef] [Green Version]
- Sachdeva, A.; Chandra, M.; Choudhary, M.; Dayal, P.; Anand, K.S. Alcohol-Related Dementia and Neurocognitive Impairment: A Review Study. Int. J. High Risk Behav. Addict. 2016, 5, e27976. [Google Scholar] [CrossRef] [Green Version]
- Stavro, K.; Pelletier, J.; Potvin, S. Widespread and Sustained Cognitive Deficits in Alcoholism: A Meta-Analysis. Addict. Biol. 2013, 18, 203–213. [Google Scholar] [CrossRef]
- Spindler, C.; Trautmann, S.; Alexander, N.; Bröning, S.; Bartscher, S.; Stuppe, M.; Muehlhan, M. Meta-Analysis of Grey Matter Changes and Their Behavioral Characterization in Patients with Alcohol Use Disorder. Sci. Rep. 2021, 11, 5238. [Google Scholar] [CrossRef]
- Yang, X.; Tian, F.; Zhang, H.; Zeng, J.; Chen, T.; Wang, S.; Jia, Z.; Gong, Q. Cortical and Subcortical Gray Matter Shrinkage in Alcohol-Use Disorders: A Voxel-Based Meta-Analysis. Neurosci. Biobehav. Rev. 2016, 66, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Chakraborty, G.; Hui, M.; Masiello, K.; Saito, M. Ethanol-Induced Neurodegeneration and Glial Activation in the Developing Brain. Brain Sci. 2016, 6, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osterndorff-Kahanek, E.A.; Tiwari, G.R.; Lopez, M.F.; Becker, H.C.; Harris, R.A.; Mayfield, R.D. Long-Term Ethanol Exposure: Temporal Pattern of MicroRNA Expression and Associated MRNA Gene Networks in Mouse Brain. PLoS ONE 2018, 13, e0190841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zingale, V.D.; Gugliandolo, A.; Mazzon, E. MiR-155: An Important Regulator of Neuroinflammation. Int. J. Mol. Sci. 2021, 23, 90. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Pei, L.; Yao, S.; Wu, Y.; Shang, Y. NLRP3 Inflammasome in Neurological Diseases, from Functions to Therapies. Front. Cell Neurosci. 2017, 11, 63. [Google Scholar] [CrossRef] [Green Version]
- Singhal, G.; Jaehne, E.J.; Corrigan, F.; Toben, C.; Baune, B.T. Inflammasomes in Neuroinflammation and Changes in Brain Function: A Focused Review. Front. Neurosci. 2014, 8, 315. [Google Scholar] [CrossRef] [Green Version]
- Hou, B.; Zhang, Y.; Liang, P.; He, Y.; Peng, B.; Liu, W.; Han, S.; Yin, J.; He, X. Inhibition of the NLRP3-Inflammasome Prevents Cognitive Deficits in Experimental Autoimmune Encephalomyelitis Mice via the Alteration of Astrocyte Phenotype. Cell Death Dis. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Trompet, S.; van Vliet, P.; de Craen, A.J.M.; Jolles, J.; Buckley, B.M.; Murphy, M.B.; Ford, I.; Macfarlane, P.W.; Sattar, N.; Packard, C.J.; et al. Pravastatin and Cognitive Function in the Elderly. Results of the PROSPER Study. J. Neurol. 2010, 257, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Alsehli, A.M.; Olivo, G.; Clemensson, L.E.; Williams, M.J.; Schiöth, H.B. The Cognitive Effects of Statins Are Modified by Age. Sci. Rep. 2020, 10, 6187. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Fernandez, C.H.; Cameron, J.-C.F. Is Statin-Associated Cognitive Impairment Clinically Relevant? A Narrative Review and Clinical Recommendations. Ann. Pharmacother. 2012, 46, 549–557. [Google Scholar] [CrossRef]
- Tiwari, V.; Kuhad, A.; Chopra, K. Epigallocatechin-3-Gallate Ameliorates Alcohol-Induced Cognitive Dysfunctions and Apoptotic Neurodegeneration in the Developing Rat Brain. Int. J. Neuropsychopharmacol. 2010, 13, 1053–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tipyasang, R.; Kunwittaya, S.; Mukda, S.; Kotchabhakdi, N.J.; Kotchabhakdi, N. Enriched Environment Attenuates Changes in Water-Maze Performance and BDNF Level Caused by Prenatal Alcohol Exposure. EXCLI J. 2014, 13, 536–547. [Google Scholar] [PubMed]
- Bathina, S.; Das, U.N. Brain-Derived Neurotrophic Factor and Its Clinical Implications. Arch. Med. Sci. 2015, 11, 1164–1178. [Google Scholar] [CrossRef]
- Stragier, E.; Martin, V.; Davenas, E.; Poilbout, C.; Mongeau, R.; Corradetti, R.; Lanfumey, L. Brain Plasticity and Cognitive Functions after Ethanol Consumption in C57BL/6J Mice. Transl. Psychiatry 2015, 5, e696. [Google Scholar] [CrossRef] [PubMed]
- King, D.S.; Wilburn, A.J.; Wofford, M.R.; Harrell, T.K.; Lindley, B.J.; Jones, D.W. Cognitive Impairment Associated with Atorvastatin and Simvastatin. Pharmacotherapy 2003, 23, 1663–1667. [Google Scholar] [CrossRef]
- Priyanka, S.H.; Thushara, A.J.; Rauf, A.A.; Indira, M. Alcohol Induced NLRP3 Inflammasome Activation in the Brain of Rats Is Attenuated by ATRA Supplementation. Brain Behav. Immun.-Health 2020, 2, 100024. [Google Scholar] [CrossRef]
- Orio, L.; Alen, F.; Pavón, F.J.; Serrano, A.; García-Bueno, B. Oleoylethanolamide, Neuroinflammation, and Alcohol Abuse. Front. Mol. Neurosci. 2019, 11, 490. [Google Scholar] [CrossRef]
- Henriksbo, B.D.; Tamrakar, A.K.; Phulka, J.S.; Barra, N.G.; Schertzer, J.D. Statins Activate the NLRP3 Inflammasome and Impair Insulin Signaling via P38 and MTOR. Am. J. Physiol.-Endocrinol. Metab. 2020, 319, E110–E116. [Google Scholar] [CrossRef]
- Koushki, K.; Shahbaz, S.K.; Mashayekhi, K.; Sadeghi, M.; Zayeri, Z.D.; Taba, M.Y.; Banach, M.; Al-Rasadi, K.; Johnston, T.P.; Sahebkar, A. Anti-Inflammatory Action of Statins in Cardiovascular Disease: The Role of Inflammasome and Toll-Like Receptor Pathways. Clin. Rev. Allergy. Immunol. 2021, 60, 175–199. [Google Scholar] [CrossRef]
- Peng, S.; Xu, L.-W.; Che, X.-Y.; Xiao, Q.-Q.; Pu, J.; Shao, Q.; He, B. Atorvastatin Inhibits Inflammatory Response, Attenuates Lipid Deposition, and Improves the Stability of Vulnerable Atherosclerotic Plaques by Modulating Autophagy. Front. Pharmacol. 2018, 9, 438. [Google Scholar] [CrossRef]
- Shu, N.; Hu, M.; Ling, Z.; Liu, P.; Wang, F.; Xu, P.; Zhong, Z.; Sun, B.; Zhang, M.; Li, F.; et al. The Enhanced Atorvastatin Hepatotoxicity in Diabetic Rats Was Partly Attributed to the Upregulated Hepatic Cyp3a and SLCO1B1. Sci. Rep. 2016, 6, 33072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, R.; Zhu, X.; Wang, J.; Yang, S.; Ma, A.; Xiao, Q.; Song, J.; Pan, X. MicroRNA-155 Promotes the Ox-LDL-Induced Activation of NLRP3 Inflammasomes via the ERK1/2 Pathway in THP-1 Macrophages and Aggravates Atherosclerosis in ApoE −/− Mice. Ann. Palliat. Med. 2019, 8, 67689. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Zeng, J.; Li, W.; Lin, L.; Zhou, X.; Tian, X.; Liu, W.; Zhang, L.; Zhang, X. Silencing of MiR-155 Suppresses Inflammatory Responses in Psoriasis through Inflammasome NLRP3 Regulation. Int. J. Mol. Med. 2018, 42, 1086–1095. [Google Scholar] [CrossRef] [Green Version]
- Afifi, O.K.; Embaby, A.S. Histological Study on the Protective Role of Ascorbic Acid on Cadmium Induced Cerebral Cortical Neurotoxicity in Adult Male Albino Rats. J. Microsc. Ultrastruct. 2016, 4, 36–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garman, R.H. Histology of the Central Nervous System. Toxicol. Pathol. 2011, 39, 22–35. [Google Scholar] [CrossRef]
- Olawale, E.O.; Shunom, A.L.; Abayomi, A. Cerebral Cortex Damage Induced by Acute Oral Alcohol Intake Is Associated with Oxidative Stress in Wistar Rats (Rattus Norvegicus). Anat. J. Afr. 2018, 7, 1113–1120. [Google Scholar] [CrossRef]
- Fortier, C.B.; Leritz, E.C.; Salat, D.H.; Venne, J.R.; Maksimovskiy, A.L.; Williams, V.; Milberg, W.P.; McGlinchey, R.E. Reduced Cortical Thickness in Abstinent Alcoholics and Association with Alcoholic Behavior. Alcohol. Clin. Exp. Res. 2011, 35, 2193–2201. [Google Scholar] [CrossRef] [Green Version]
- Orning, P.; Lien, E. Multiple Roles of Caspase-8 in Cell Death, Inflammation, and Innate Immunity. J. Leukoc. Biol. 2021, 109, 121–141. [Google Scholar] [CrossRef]
- Burguillos, M.A.; Deierborg, T.; Kavanagh, E.; Persson, A.; Hajji, N.; Garcia-Quintanilla, A.; Cano, J.; Brundin, P.; Englund, E.; Venero, J.L.; et al. Caspase Signalling Controls Microglia Activation and Neurotoxicity. Nature 2011, 472, 319–324. [Google Scholar] [CrossRef]
- Gurung, P.; Kanneganti, T.-D. Novel Roles for Caspase-8 in IL-1β and Inflammasome Regulation. Am. J. Pathol. 2015, 185, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Song, P.; Li, N.; Li, X.; Chen, J. A Plastic Stabilizer Dibutyltin Dilaurate Induces Subchronic Neurotoxicity in Rats. Neural. Regen. Res. 2012, 7, 2213–2220. [Google Scholar] [CrossRef]
- Chandra, S.; Jana, M.; Pahan, K. Aspirin Induces Lysosomal Biogenesis and Attenuates Amyloid Plaque Pathology in a Mouse Model of Alzheimer’s Disease via PPARα. J. Neurosci. 2018, 38, 6682–6699. [Google Scholar] [CrossRef] [PubMed]
- Jorda, A.; Aldasoro, M.; Aldasoro, C.; Guerra-Ojeda, S.; Iradi, A.; Vila, J.M.; Campos-Campos, J.; Valles, S.L. Action of Low Doses of Aspirin in Inflammation and Oxidative Stress Induced by Aβ 1-42 on Astrocytes in Primary Culture. Int. J. Med. Sci. 2020, 17, 834–843. [Google Scholar] [CrossRef] [PubMed]
- Raza, H.; John, A. Implications of Altered Glutathione Metabolism in Aspirin-Induced Oxidative Stress and Mitochondrial Dysfunction in HepG2 Cells. PLoS ONE 2012, 7, e36325. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups Latency to Reach Target Quadrant (s) | Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | Day 6 | % Time Spent in Target Quadrant |
|---|---|---|---|---|---|---|---|
| Control group | 6.68 ± 19.9 | 27.88 ± 6.6 | 22.18 ± 8.1 | 15.3 ± 7.5 | 8.37 ± 2.5 | 7.7 ± 4.9 | 47.33 ± 7.5 |
| Alcohol group | 10.9 ± 9.9 a | 67.63 ± 5.4 a | 92.9 ± 7.1 a | 55.52 ± 6.3 a | 45.38 ± 8.2 a | 21.08 ± 7.2 a | 19.25 ± 5.9 a |
| ATOR group | 8.06 ± 17.2 a | 50.77 ± 10.2 a | 80.9 ± 9.8 a | 32.6 ± 5.8 a | 35.33 ± 13.3 a | 19.17 ± 6.4 a | 21.6 ± 10.3 a |
| Alcohol + ATOR group | 13.71 ± 7.3 a | 85.19 ± 8.8 a | 71.46 ± 7.7 a | 48.23 ± 13.2 a | 55.5 ± 11.9 a | 33.17 ± 9.2 a | 12.81 ± 14.9 a |
| Alcohol + ATOR + ASA group | 7.73 ± 16.6 abcd | 33.1 ± 15.1 abcd | 32.23 ± 7.1 abcd | 21.17 ± 11.4 abcd | 21.9 ± 11.0 abcd | 15.08 ± 5.2 abcd | 36.6 ± 18.2 bcd |
| Group | Mean Thickness of Frontal Cortex |
|---|---|
| Group I | 1832.15 ± 198.5 |
| Group II | 1455.60 ± 78.3 a |
| Group III | 1644.70 ± 185.2 b |
| Group IV | 1554.16 ± 69.8 c |
| Group V | 1800.70 ± 167.3 d |
| Group | Area % of Caspase-8 Positive Immune Reaction |
|---|---|
| Group I | 0 |
| Group II | 4.41 ± 0.5 a |
| Group III | 1.75 ± 0.3 b |
| Group IV | 2.73 ± 0.4 c |
| Group V | 0.42 ± 0.1 d |
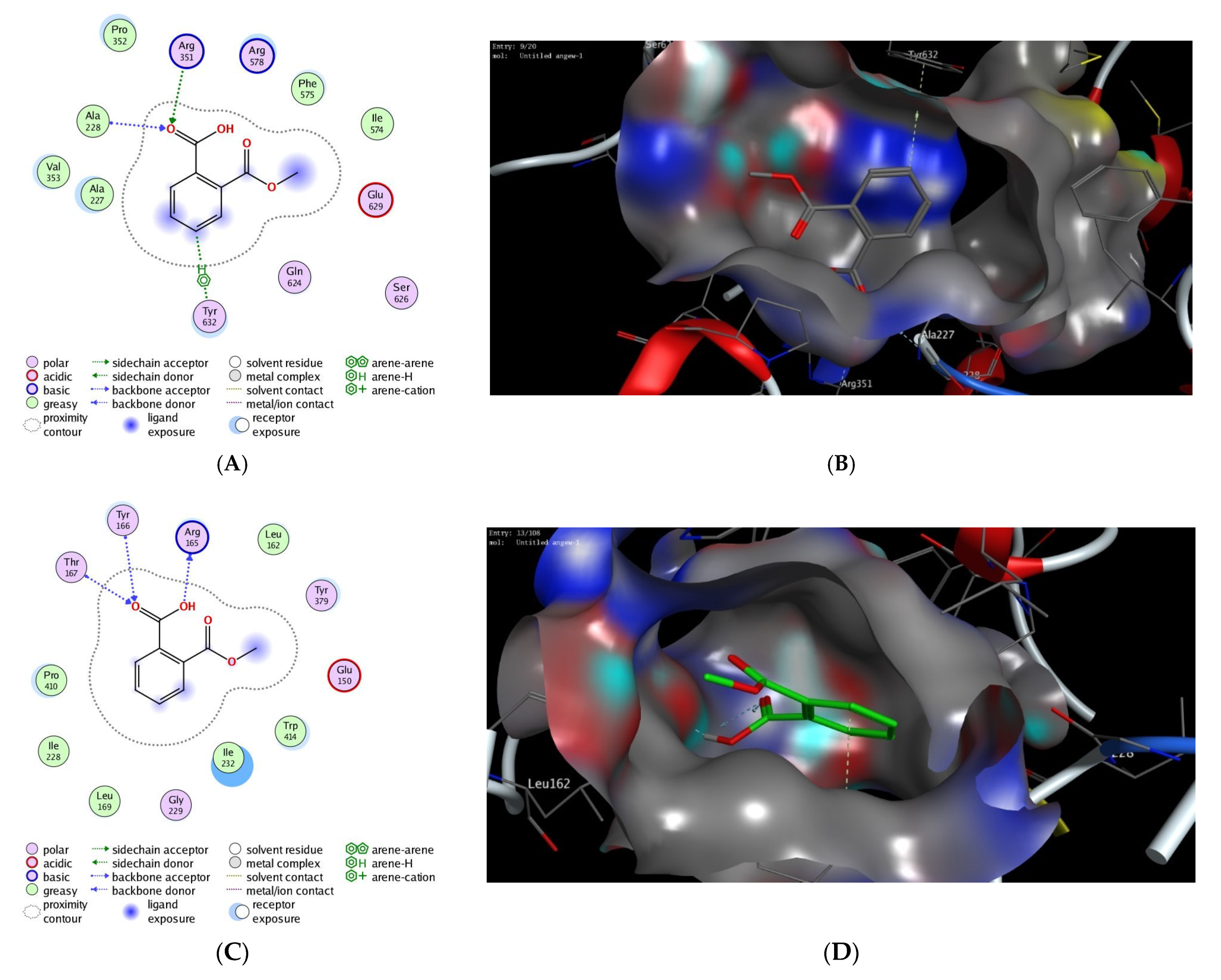
| PDB | Docking Score (kcal/mol) | Interactive Residues | |
|---|---|---|---|
| Hydrophilic Interactions | Hydrophobic Interactions | ||
| 7alv | −12.17 | Ala228, Arg351, Tyr632 | Val353, Ile574, Phe575, Pro352, Ala227 |
| 6npy | −11.68 | Arg165, Thr167, Tyr166 | Leu162, Ile228, Trp414, Leu169, Pro410, Ile232 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed, D.I.; Alaa El-Din Aly El-Waseef, D.; Nabih, E.S.; El-Kharashi, O.A.; Abd El-Kareem, H.F.; Abo Nahas, H.H.; Abdel-Wahab, B.A.; Helmy, Y.A.; Alshawwa, S.Z.; Saied, E.M. Acetylsalicylic Acid Suppresses Alcoholism-Induced Cognitive Impairment Associated with Atorvastatin Intake by Targeting Cerebral miRNA155 and NLRP3: In Vivo, and In Silico Study. Pharmaceutics 2022, 14, 529. https://doi.org/10.3390/pharmaceutics14030529
Mohamed DI, Alaa El-Din Aly El-Waseef D, Nabih ES, El-Kharashi OA, Abd El-Kareem HF, Abo Nahas HH, Abdel-Wahab BA, Helmy YA, Alshawwa SZ, Saied EM. Acetylsalicylic Acid Suppresses Alcoholism-Induced Cognitive Impairment Associated with Atorvastatin Intake by Targeting Cerebral miRNA155 and NLRP3: In Vivo, and In Silico Study. Pharmaceutics. 2022; 14(3):529. https://doi.org/10.3390/pharmaceutics14030529
Chicago/Turabian StyleMohamed, Doaa I., Dalia Alaa El-Din Aly El-Waseef, Enas S. Nabih, Omnyah A. El-Kharashi, Hanaa F. Abd El-Kareem, Hebatallah H. Abo Nahas, Basel A. Abdel-Wahab, Yosra A. Helmy, Samar Zuhair Alshawwa, and Essa M. Saied. 2022. "Acetylsalicylic Acid Suppresses Alcoholism-Induced Cognitive Impairment Associated with Atorvastatin Intake by Targeting Cerebral miRNA155 and NLRP3: In Vivo, and In Silico Study" Pharmaceutics 14, no. 3: 529. https://doi.org/10.3390/pharmaceutics14030529