
Differential Responses of Antioxidative System during the Interaction of Soursop Fruits (Annona muricata L.) and Nectria haematococca at Postharvest Storage

 , and
, and
Abstract
:1. Introduction
2. Results
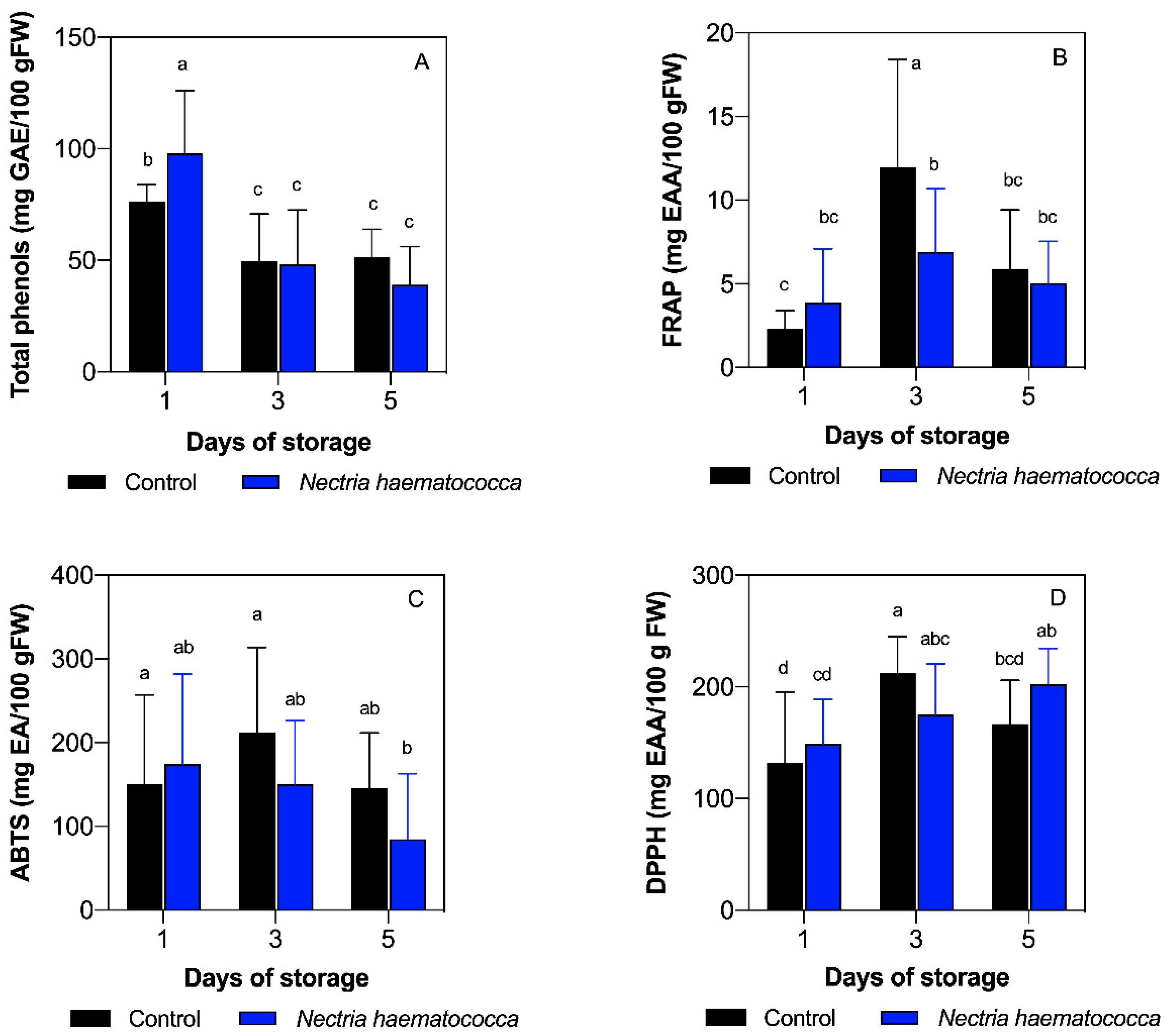
2.1. Total Phenolic Compounds
2.2. Ferric Reducing/Antioxidant Power (FRAP) Capacity
2.3. 2,2′-Azinobis-(3-ethylbenzothiazoline-6-sulfonate) (ABTS•+) Antioxidant Capacity
2.4. 2,2′-Diphenyl-1-picrylhydrazyl Radical (DPPH•) Antioxidant Capacity
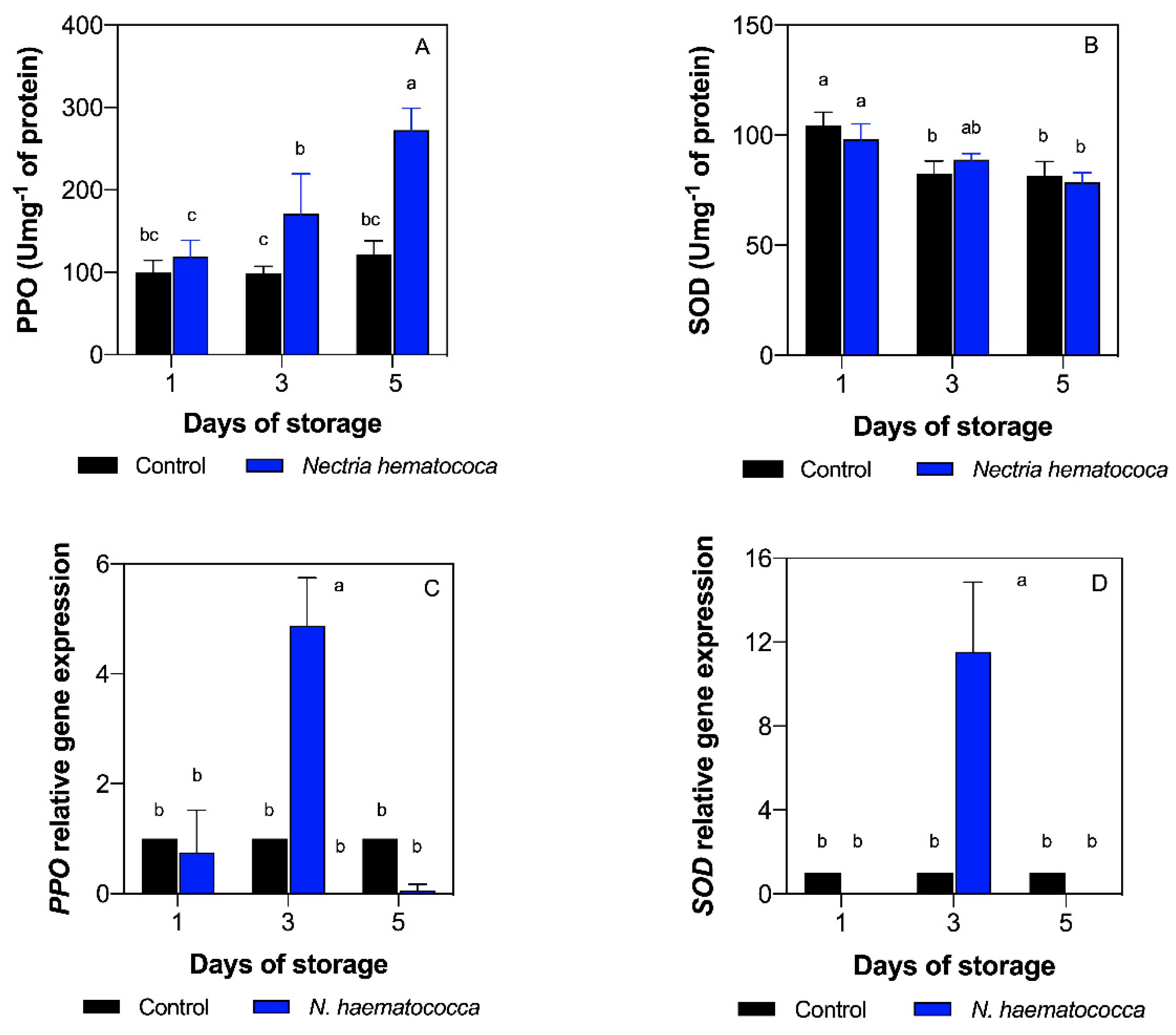
2.5. Polyphenol Oxidase (PPO) and Superoxide Dismutase (SOD) Enzyme Activity
2.6. Relative Gene Expression (PPO and SOD)
2.7. Multivariate Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Inoculation of Fruits
4.3. Analysis of Total Extractable Phenolic Compounds and Antioxidant Capacity
4.3.1. Crude Extract
4.3.2. Total Extractable Phenols
4.3.3. Antioxidant Capacity by the DPPH Method
4.3.4. Antioxidant Capacity by the ABTS•+ Method
4.3.5. Iron Reduction Antioxidant Capacity (FRAP)
4.4. Enzymatic Activity
4.4.1. Crude Enzyme Extraction
4.4.2. Polyphenol Oxidase (EC. 1.14.18.1; PPO)
4.4.3. Superoxide Dismutase (EC. 1.15.1.1; SOD)
4.5. Relative Gene Expression
4.5.1. RNA Extraction and cDNA Synthesis
4.5.2. Gene Selection and Primer Design
4.5.3. PCR
4.5.4. Gene Expression Analysis by qRT-PCR
4.5.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Servicio de Información Agroalimentaria y Pesquera (SIAP). 2019. Available online: https://nube.siap.gob.mx/cierreagricola/ (accessed on 16 June 2021).
- Márquez-Cardozo, C.J.; Villacorta-Lozano, V.; Yepes-Betancur, D.P.; Ciro-Velásquez, H.J.; Cartagena-Valenzuela, J.R. Physiological and physico-chemical characterization of the soursop fruit (Annona muricata L. cv. Elita). Rev. Fac. Nac. de Agron. Medellín. 2012, 65, 6477–6486. [Google Scholar]
- Berumen-Varela, G.; Hernández-Oñate, M.A.; Tiznado-Hernández, M.E. Utilization of biotechnological tools in soursop (Annona muricata L.). Sci. Hortic. 2019, 245, 269–273. [Google Scholar] [CrossRef]
- Mondal, K.; Malhotra, S.P.; Jain, V.; Singh, R. Oxidative stress and antioxidant systems in Guava (Psidium guajava L.) fruits during ripening. Physiol. Mol. Biol. Plants 2009, 15, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, E.; Cruz, M.; Melgarejo, L.M.; Marquínez, X.; Hoyos-Carvajal, L. Histopathological features of infections caused by Fusarium oxysporum and F. solani in purple passionfruit plants (Passiflora edulis Sims). Summa Phytopathol. 2014, 40, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Guerrero, S.E.; Balois-Morales, R.; Bautista-Rosales, P.U.; López-Guzmán, G.G.; Berumen-Varela, G.; Palomino-Hermosillo, Y.A.; León-Fernandez, A.E. Identification of Fungal Pathogens of Mango and Soursop Fruits Using Morphological and Molecular Tools and Their Control Using Papaya and Soursop Leaf and Seed Extracts. Int. J. Agron. 2020, 2020, 8962328. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.; Dicke, M. Plant interactions with microbes and insects: From molecular mechanisms to ecology. Trends Plant Sci. 2007, 12, 564–569. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Melgarejo, A.; Balois-Morales, R.; Palomino-Hermosillo, Y.A.; López-Guzmán, G.G.; Ramírez-Ramírez, J.C.; Cervantes-García, E.; Bautista-Rosales, P.U. Phytochemical and Antioxidant Dynamics of the Soursop Fruit (Annona muricata L.) in response to Colletotrichum spp. J. Food Qual. 2020, 2020, 3180634. [Google Scholar] [CrossRef]
- Delaux, P.M.; Nanda, A.K.; Mathé, C.; Sejalon-Delmas, N.; Dunand, C. Molecular and biochemical aspects of plant terrestrialization. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 49–59. [Google Scholar] [CrossRef]
- Lin, J.; Gong, D.; Zhu, S.; Zhang, L.; Zhang, L. Expression of PPO and POD genes and contents of polyphenolic compounds in harvested mango fruits in relation to Benzothiadiazole-induced defense against anthracnose. Sci. Hort. 2011, 130, 85–89. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Chun, S.C. Expression of PR-protein genes and induction of defense-related enzymes by Bacillus subtilis CBR05 in tomato (Solanum lycopersicum) plants challenged with Erwinia carotovora subsp. carotovora. Biosci. Biotechnol. Bichem. 2016, 80, 2277–2283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harbome, J. Secondary products|phytoalexins, Natural Plant Protection. In Encyclopedia of Applied Plant Sciences, 1st ed.; Thomas, B., Murphy, D.J., Murray, B.G., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 2003; pp. 1152–1159. [Google Scholar]
- Villa-Martínez, A.; Pérez-Leal, R.; Morales-Morales, H.A.; Basurto-Sotelo, M.; Soto-Parra, J.M.; Martínez-Escudero, E. Situación actual en el control de Fusarium spp. y evaluación de la actividad antifúngica de extractos vegetales. Acta Agron. 2015, 64, 194–205. [Google Scholar] [CrossRef]
- Pedras, M.; Yaya, E.; Glawischnig, E. The phytoalexins from cultivated and wild crucifers: Chemistry and biology. Nat. Prod. Rep. 2011, 28, 1381–1405. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Wang, J.; Zhu, R.; Lu, H.; Zheng, X.; Yu, T. Transcript profiling analysis of Rhodosporidium paludigenum-mediated signaling pathways and defense responses in mandarin orange. Food Chem. 2015, 172, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Aritua, V.; Achor, D.; Gmitter, F.G.; Albrigo, G.; Wang, N. Transcriptional and microscopic analyses of citrus stem and root responses to Candidatus Liberibacter asiaticus infection. PLoS ONE 2013, 8, e73742. [Google Scholar] [CrossRef] [Green Version]
- Syhida, M.; Maskat, M.; Suri, R.; Mamot, S.; Hadijah, H. Sourp (Anona muricata L.). Blood hematology and serum biochemistry of Sprague Dawley rats. Int. Food Res. J. 2012, 19, 955–959. [Google Scholar]
- Javanmardi, J.; Kubota, C. Variation of lycopene, antioxidant activity, total soluble solids and weight loss of tomato during postharvest storage. Postharvest Biol. Technol. 2006, 41, 151–155. [Google Scholar] [CrossRef]
- Agrios, G. Plant Pathology, 5th ed.; Elsevier Academic Press: London, UK, 2005; pp. 920–922. [Google Scholar]
- Spoel, S.H.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef]
- Akomolafe, S.; Ajayi, O. A comparative study on antioxidant properties, proximate and mineral compositions of the peel and pulp of ripe Annona muricata (L.) fruit. Int. Food Res. J. 2015, 22, 2381. [Google Scholar]
- Gómez, M.R.; Rodríguez, A. Mecanismos de defensa y respuestas de las plantas en la interacción micorrícica: Una revisión. Rev. Col. Biotecnol. 2012, 14, 271–284. [Google Scholar]
- Balois-Morales, R.; Jiménez-Zurita, J.O.; Alia-Tejacal, I.; López-Guzmán, G.G.; Palomino-Hermosillo, Y.A.; Sánchez-Herrera, L.M. Antioxidant enzymes and antioxidant activity in two soursop selections (Annona muricata L.) from Nayarit, Mexico stored at 15 °C. Rev. Bras. Frutic. 2019, 41, e-083. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.; Alvez, R.; Filgueiras, H. Mudacas relacionadas ao amaciamento da graviola durante a maturacao pós-colheita. Pesqui. Agropecu. Bras. 2006, 41, 1707–1713. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Zurita, J.; Balois-Morales, R.; Alia-Tejacal, I.; Herrera, S.; Mónica, L.; Jiménez-Ruiz, E.; Bello-Lara, J.; García-Paredes, J.; Juárez-López, P. Cold Storage of Two Selections of Soursop (Annona muricata L.) in Nayarit, Mexico. J. Food Qual. 2017, 2017, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sellés-Marchart, S.; Casado-Vela, J.; Bru-Martínez, R. Isolation of a latent polyphenol oxidase from loquat fruit (Eriobotrya japonica Lindl.): Kinetic characterization and comparison with the active form. Arch. Biochem. Biophys. 2006, 446, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Chávez, R.A.; Martínez Peniche, R.Á.; Hernández-Iturriaga, M.; Teixidó-Espasa, N.; Usall-Rodié, J.; Viñas-Almenar, I.; Torres-Sanchis, R. Mechanisms of resistance in postharvest fruit-pathogen interaction. Rev. Chapingo Ser. Hortic. 2015, 21, 185–198. [Google Scholar] [CrossRef]
- Radi, M.; Mahrouz, M.; Jaouad, A.; Tacchini, M.; Aubert, S.; Hugues, M.; Amiot, M.J. Phenolic composition, browning susceptibility and carotenoid content of several apricot cultivars at maturity. HortScience 1997, 32, 1087–1091. [Google Scholar] [CrossRef] [Green Version]
- Vivanco, J.; Cosio, E.; Loloya-Vargas, V.; Flores, H. Mecanismos químicos de defensa en las plantas. Investigación y Ciencia 2005, 341, 68–75. [Google Scholar]
- Thipyapong, P.; Hunt, M.D.; Steffens, J.C. Systemic wound induction of potato (Solanum tuberosum) polyphenol oxidase. Phytochemistry 1995, 40, 673–676. [Google Scholar] [CrossRef]
- Khodadadi, F.; Tohidfar, M.; Mohayeji, M.; Dandekar, A.M.; Leslie, C.A.; Kluepfel, D.A.; Vahdati, K. Induction of polyphenol oxidase in walnut and its relationship to the pathogenic response to bacterial blight. J. Am. Soc. Hort. Sci. 2016, 141, 119–124. [Google Scholar] [CrossRef]
- Mo, Y.; Gong, D.; Liang, G.; Han, R.; Xie, J.; Li, W. Enhanced preservation effects of sugar apple fruits by salicylic acid treatment during post-harvest storage. J. Sci. Food Agric. 2008, 88, 2693–2699. [Google Scholar] [CrossRef]
- Wang, Y.; Ji, D.; Chen, T.; Li, B.; Zhang, Z.; Qin, G.; Tian, S. Production, signaling, and scavenging mechanisms of reactive oxygen species in fruit–pathogen interactions. Int. J. Mol. Sci. 2019, 20, 2994. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Silva, S.; Voigt, E.; Silva, N.; Maia, J.; Aragáo, T.; Silveira, J. Partial oxidative protection by enzymatic and non-enzymatic components an cashew under high salinity. Biol. Plantarum. 2012, 56, 172–176. [Google Scholar] [CrossRef]
- Huan, C.; Jiang, L.; An, X.; Yu, M.; Xu, Y.; Ma, R.; Yu, Z. Potential role of reactive oxygen species and antioxidant genes in the regulation of peach fruit development and ripening. Plant Physiol. Biochem. 2016, 104, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Lightfoot, D.J.; Mcgrann, G.R.; Able, A.J. The role of a cytosolic superoxide dismutase in barley–pathogen interactions. Mol. Plant Pathol. 2017, 18, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Miura, C.; Sugawara, K.; Neriya, Y.; Minato, N.; Keima, T.; Himeno, M.; Namba, S. Functional characterization and gene expression profiling of superoxide dismutase from plant pathogenic phytoplasma. Gene 2012, 510, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, X.; Dong, C.; Zhang, N.; Ban, Z.; Li, L.; Chen, C. Effects of ozone treatment on SOD activity and genes in postharvest cantaloupe. RSC Adv. 2020, 10, 17452–17460. [Google Scholar] [CrossRef]
- Prieto, H.; Utz, D.; Castro, Á.; Aguirre, C.; González-Agüero, M.; Valdés, H.; Campos-Vargas, R. Browning in Annona cherimola fruit: Role of polyphenol oxidase and characterization of a coding sequence of the enzyme. J. Agric. Food Chem. 2007, 55, 9208–9218. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Rosales, P.U.; Calderon-Santoyo, M.; Servín-Villegas, R.; Ochoa-Álvarez, N.A.; Ragazzo-Sánchez, J.A. Action mechanisms of the yeast Meyerozyma caribbica for the control of the phytopathogen Colletotrichum gloeosporioides in mangoes. Biol. Control 2013, 65, 293–301. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Brand-Williams, W.; Culivier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebensmittel Wissenschaft and Technologie. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS•+ radical action decolonization assay free. Rad. Biol. Med. 1999, 26, 1231–1337. [Google Scholar] [CrossRef]
- Yen, G.-C.; Chen, H.-Y. Antioxidant Activity of Various Tea Extracts in Relation to Their Antimut agenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Lamikanra, O. Enzymatic browning of Muscadine grapes products. In Enzymatic Browning and Its Prevention; Lee, C.L., Whitaker, J.L., Eds.; ACS: Washington, DC, USA, 1995; pp. 166–177. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Berumen-Varela, G.; Palomino-Hermosillo, Y.A.; Bautista-Rosales, P.U.; Peña-Sandoval, G.R.; López-Guzmán, G.G.; Balois-Morales, R. Identification of reference genes for quantitative real-time PCR in different developmental stages and under refrigeration conditions in soursop fruits (Annona muricata L.). Sci. Hort. 2020, 260, 108893. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–404. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene | Primer sequence 5′–3′ | Size (bp) |
|---|---|---|
| PPO | Fw: AAGCCAGCATCCGAAGAGAG | 150 |
| Rv: GTCGTAGCTCAGCTGCTTCA | ||
| SOD | Fw: GGCCAAACTCCATCATTG Rv: GCATTTCCAGTGGTCTTG | 97 |
| UBC | Fw: AACCTCTATCCAGTCTCTCCTC | 128 |
| Rv: TGAGATAGTGGAGCAGAGCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubio-Melgarejo, A.; Balois-Morales, R.; Ochoa-Jiménez, V.A.; Casas-Junco, P.P.; Jiménez-Zurita, J.O.; Bautista-Rosales, P.U.; Berumen-Varela, G. Differential Responses of Antioxidative System during the Interaction of Soursop Fruits (Annona muricata L.) and Nectria haematococca at Postharvest Storage. Plants 2021, 10, 1432. https://doi.org/10.3390/plants10071432
Rubio-Melgarejo A, Balois-Morales R, Ochoa-Jiménez VA, Casas-Junco PP, Jiménez-Zurita JO, Bautista-Rosales PU, Berumen-Varela G. Differential Responses of Antioxidative System during the Interaction of Soursop Fruits (Annona muricata L.) and Nectria haematococca at Postharvest Storage. Plants. 2021; 10(7):1432. https://doi.org/10.3390/plants10071432
Chicago/Turabian StyleRubio-Melgarejo, Alejandro, Rosendo Balois-Morales, Verónica Alhelí Ochoa-Jiménez, Paloma Patricia Casas-Junco, José Orlando Jiménez-Zurita, Pedro Ulises Bautista-Rosales, and Guillermo Berumen-Varela. 2021. "Differential Responses of Antioxidative System during the Interaction of Soursop Fruits (Annona muricata L.) and Nectria haematococca at Postharvest Storage" Plants 10, no. 7: 1432. https://doi.org/10.3390/plants10071432