
Realizing the Potential of Camelina sativa as a Bioenergy Crop for a Changing Global Climate


1
MS330/Department of Biochemistry & Molecular Biology, University of Nevada, Reno, NV 89557, USA
2
Department of Agriculture, Veterinary & Rangeland Sciences, University of Nevada, Reno, NV 89557, USA
*
Author to whom correspondence should be addressed.
Plants 2022, 11(6), 772; https://doi.org/10.3390/plants11060772
Submission received: 11 February 2022
/
Revised: 28 February 2022
/
Accepted: 4 March 2022
/
Published: 14 March 2022
(This article belongs to the Special Issue 10th Anniversary of Plants—Recent Advances and Perspectives)
Abstract
:Camelina sativa (L.) Crantz. is an annual oilseed crop within the Brassicaceae family. C. sativa has been grown since as early as 4000 BCE. In recent years, C. sativa received increased attention as a climate-resilient oilseed, seed meal, and biofuel (biodiesel and renewable or green diesel) crop. This renewed interest is reflected in the rapid rise in the number of peer-reviewed publications (>2300) containing “camelina” from 1997 to 2021. An overview of the origins of this ancient crop and its genetic diversity and its yield potential under hot and dry growing conditions is provided. The major biotic barriers that limit C. sativa production are summarized, including weed control, insect pests, and fungal, bacterial, and viral pathogens. Ecosystem services provided by C. sativa are also discussed. The profiles of seed oil and fatty acid composition and the many uses of seed meal and oil are discussed, including food, fodder, fuel, industrial, and medical benefits. Lastly, we outline strategies for improving this important and versatile crop to enhance its production globally in the face of a rapidly changing climate using molecular breeding, rhizosphere microbiota, genetic engineering, and genome editing approaches.
1. Introduction
Camelina sativa (L.) Crantz., also known as false or wild flax, German sesame, gold-of-pleasure, or linseed dodder, is an allohexaploid (2n = 40) oilseed crop within the Brassicaceae [1,2,3,4,5,6]. Interest in C. sativa increased in recent years due to its adaptability to diverse environmental conditions, low requirements for water and nutrients, relatively strong resistance to insect pests and microbial diseases, and unique oil composition and characteristics suitable for the production of food and fodder, biofuels, and bio-based products [1,2,7,8]. These positive agronomic traits and environmental attributes, along with the recent development of methods for transgenesis [9,10,11,12,13] and CRISPR/Cas genome editing [14,15,16], triggered great interest in C. sativa as an industrial oilseed crop. The ongoing interest in C. sativa is documented by the large number of peer-reviewed publications from various databases retrieved when “camelina” was used as a search term, for example, in a query of the ScienceDirect (2309 publications from 1997–2021), Web of Science (1525 publications from 2000–2021) and Agricola (677 publications from 2000–2021) databases (reported on December 30, 2021). The large number of publications and data reported on C. sativa highlight the immense potential of this crop and the interest in genetic improvement to allow it to gain more widespread acceptance and economic viability. Most publications focus on the uses of C. sativa oil and meal, the composition of oil and fatty acids, its genetics and breeding, its physiology, and its production and agronomic management. Agronomic management refers to the practice of minimizing input factors such as fertilizer, irrigation, tillage, herbicides, fungicides, and insecticides to maximize crop yield outputs such as seed yield, oil content, and biodiesel production. Many reviews report on various aspects of its general use as an oilseed crop [2,4,17,18,19,20,21,22,23,24], and as a platform for the production of biofuels [25,26,27,28] and industrial lipids [1,6,8]. After providing some essential background, we summarize the present status of C. sativa research and identify areas for its future improvement with a particular emphasis on enhancing the climate resilience of this highly versatile crop.
2. Origin and Distribution
Camelina sativa is an ancient crop, known as early as 4000 BCE in Auvervier, Switzerland [29] with evidence of widespread cultivation throughout northern Europe from Southern Scandinavia [6,30] to central Asia (eastern Turkey), from 700–900 BCE [31] to the Iron Age (100 CE–250 BCE) [24]. Archaeological sites revealed evidence that C. sativa was cultivated for food and oil production in Scandinavia, Romania, and eastern Turkey during the late Stone Age and middle Bronze Age (1800 BCE) [32], with widespread availability during the late Bronze age (1200 BCE) [30]. C. sativa cultivation declined during the Medieval Age, but grew during the last century throughout northern, central and eastern Europe, the Balkans, and Russia and to some extent in North America [33]. However, much of its former cultivation all but disappeared, being replaced almost entirely by rapeseed (Brassica napus L.) [24].
C. sativa was likely introduced to the Americas as a weed in flax, thus giving rise to the name false flax [4]. Today, C. sativa is cultivated throughout the northern USA and southern Canada [34]. Within the USA, C. sativa is grown effectively in the Pacific Northwest, across the North and Central Plains into the Corn Belt region [35], and in the arid Southwest with irrigation [36,37]. Within Canada, C. sativa is grown widely from the western Prairie Provinces [38,39] to the eastern Maritime Provinces [40].
3. Genetic Diversity and Morphological Variation
C. sativa belongs to the tribe Camelineae within the Brassicaceae. The similarity between C. sativa and Arabidopsis thaliana [41,42] makes A. thaliana a useful reference for the development of genetic and genomic tools in C. sativa [34]. The genus Camelina has up to 11 species, revealing taxonomic dissimilarity with its center of diversity in Eurasia (Russia or Ukraine) [43,44,45]. Among 11 species, five species, namely C. sativa, C. microcarpa, C. rumelica, C. alyssum, and C. hispida, are found in Europe and three species, namely C. sativa, C. microcarpa, and C. alyssum, are found in the USA and Canada [11]. Among these species, only C. sativa and C. microcarpa are cultivated [11]. C. sativa is an allohexaploid species with 2n = 6x = 40 [34,46]. Other Camelina species such as C. hispida (Boiss.) Hedge (2n = 2x = 14), C. neglecta (2n = 2x = 12) [44], and C. laxa C.A. Mey. (2n = 2x= 12) [47] are diploid. Other species are polyploid, including C. rumelica Velen., tetraploid (2n = 4x = 26); C. microcarpa Andrz. ex DC with three cytotypes: diploid (2n = 2x = 12), tetraploid (2n = 4x = 26), and hexaploid (2n = 6x = 40) [48]; and C. sativa, hexaploid (2n = 2x= 40) [49,50]. Morphologically, C. sativa and C. microcarpa are very similar except for the smaller seed size of C. microcarpa [51]. This phenotypic similarity suggests that C. sativa could potentially be the domesticated form of C. microcarpa [43]. Greater variations in chromosome counts within species might arise from intraspecific ploidy variations or inaccurate records suggesting past taxonomic misidentification [52]. Investigations into the genome structure of 193 Camelina accessions revealed three subpopulations, with two represented by domesticated C. sativa accessions and one composed of C. microcarpa species that included a newly designated C. neglecta diploid (née C. microcarpa) species [53]. Recent molecular phylogenies derived from chloroplast genome sequencing of 84 individuals revealed low intragenic variation across the Camelina genus [54]. However, cytotypes and chromosome counts across 82 individuals confirmed that the tetraploid C. microcarpa (or C. neglecta-like) is the proposed maternal parent and the diploid C. hispida is the proposed paternal parent of C. sativa [53,54].
Resembling other genera within the Brassicaceae, Camelina species are dicotyledonous with high morphological plasticity. Biologically, Camelina species can be annual spring or biennial winter types [2], with some species requiring vernalization to induce flowering [55]. Most domesticated C. sativa are spring types, whereas most wild relatives are winter types. Winter types are well adapted as cover crops in double- or relay-cropping systems, with soybean (Glycine max L. Merr.) and other short-seasoned summer crops due to their early maturity in the Great Plains and the Upper Midwestern USA [3,35,56,57,58]. C. sativa typically has a short growing cycle of 85–120 days [4,5,22,50,59]. Growing degree day (GDD) requirements for C. sativa during its complete life cycle (with up to 75% ripe silicles) generally range from 1200 to 1300 °C with a base temperature of 5 °C [60]. Plant height ranges from 30 to 120 cm depending upon cultivars, growing season, and amount of nitrogen fertilizer used [4,36,59,61,62]. Stems are either hairy or smooth, are branched, and become lignified when mature. Leaves are arrow-shaped and pointed, approximately 5–8 cm long with smooth to undulated edges. Flowers are 5–7 mm in diameter and are mostly autogamous [49]. Flowers are pale yellow in color and arranged within inflorescences, called raceme. The silicles are 5–14 mm long, slightly flattened, and pear-shaped, containing 8–15 golden to brown colored seeds at maturity. C. sativa seeds are very small, with 1000 seed weight varying between 0.8 to 1.8 g, depending upon the cultivar and growing environments during seed growth and development [4,22,24,63,64]. C. sativa has a deep taproot system, varying with soil type and growing conditions, which is thought to improve nutrient scavenging. The use of C. sativa as a cover crop can reduce nutrient run-off, particularly for winter annual cultivars [2]. C. sativa can also reduce soil compaction and improve infiltration capacity, similar to other species within the Brassicaceae [2].
4. C. sativa Yield Potential under Hot and Dry Conditions
Global climate change is related to increasing surface aridification and the increasing duration and frequency of droughts in many regions of the world [65,66]. Interest in C. sativa as a low-input crop for use on marginal lands has grown in recent years [50,67,68,69,70,71]. Thus, recent studies investigated C. sativa grown under semi-arid conditions with limited water inputs, with the goal of assessing its performance under reduced water and fertilizer inputs [36,37,59,62,72].
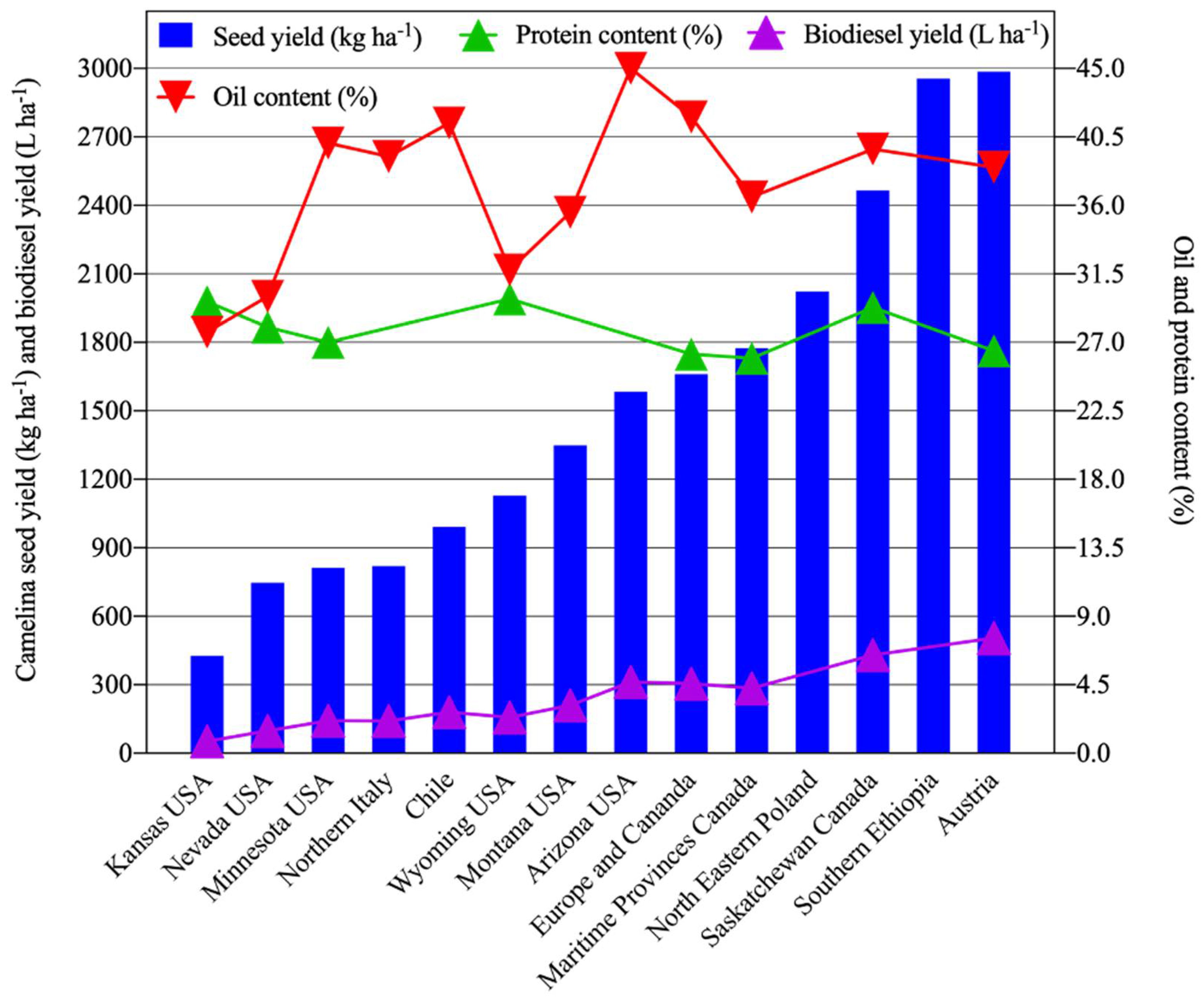
Many studies were conducted across semi-arid or arid regions of the globe with distinct mean annual precipitation levels to compare seed yield (kg ha−1), oil content (%), protein content (%), and biodiesel production (L ha−1) (Figure 1, Table 1). Differences in cultivars (genotypes), water availability, environment, physical and chemical characteristics of soils, and management practices such as irrigation, nitrogen application, sowing date, seeding methods and rates, clearly impact overall C. sativa productivity. Among these studies, the overall mean seed yield of C. sativa was 1410 kg ha−1 (Figure 1, Table 1). The highest reported seed yield ranges were attained in Austria (2419–3625 kg ha−1) [73] and in southern Ethiopia (2795–3200 kg ha−1) [74]. The lowest C. sativa seed yield ranges were reported by studies performed in Kansas [75] and Nevada, USA [36,37,59,62]. Low seed yield of C. sativa may occur due to drought and high temperatures during the flowering and pod filling stage [61,76], as well as poor soil quality [77]. Consistent with these findings, environmental factors (e.g., temperature and rainfall) were responsible for approximately 73% of the variation in seed yield in north-eastern Poland [72]. In contrast, only ~6% of the variation in seed yield was due to genetic factors [72]. Mild weather conditions, along with moderate in-season precipitation, favor higher seed yields [35,39]. Additionally, high C. sativa seed yields were associated with adequate moisture and mild temperatures during seed filling, which is critical for the production and transport of sugars from source to sink tissues [3,77,78]. In contrast, low seed yields were observed under conditions of hot temperature and low precipitation [79,80]. Studies showed an increase in seed yields ranging from 425 to 2867 kg ha−1 as a result of increasing amounts of applied irrigation ranging from 187 to 536 mm [36,69,70,81]. Seasonal water requirements range from 332–490 mm for C. sativa based upon cumulative evapotranspiration (ET) estimates [82,83,84]. Notably, C. sativa might serve as a reliable alternative to Canola in locations where seasonal water supply is less than 250 mm [85]. In addition to increased water inputs, increasing nitrogen fertilizer application can increase seed yield. Seed yields of 1800 kg ha−1 can be attained with 150 kg N ha−1 and water inputs in the range of 320–376 mm [86]. Similar estimates for optimal N input were reported for rainfed field settings [87,88]. The use of organic fertilizers during the production of C. sativa for animal feed reduces its environmental impact [89].
Another important measure of productivity for any oilseed crop is oil and protein production of the seed meal. For C. sativa, oil and protein content varied widely across various parts of the world, with overall mean seed oil and protein contents of ~36% and ~28%, respectively (Figure 1, Table 1). Highest seed oil contents were reported in the range of 39.8–45.7% in Chile [61], followed by 45% in Arizona, USA [84], and 38–43% in western Canada [39]. The lowest seed oil content, ranging from 25.7 to 31.8%, was observed in Nevada, USA [37]. Similarly to seed yield, oil yield is strongly influenced by environmental factors [64,90]. For example, oil content variation was chiefly associated with mean air temperature and water availability during seed development [61].

{kind=link}
{kind=link}
Table 1.
C. sativa seed yield, oil, and protein content reported across various regions of the world.
Table 1.
C. sativa seed yield, oil, and protein content reported across various regions of the world.
| Locations | Seed Yield (kg ha−1) | Seed Oil Content (%) 1 | Seed Meal Protein Content (%) 2 | Biodiesel Yield (L ha−1) 3 | Major Sources of Variation | Reference | |
|---|---|---|---|---|---|---|---|
| Mean | Range | Range | Range | Mean | |||
| Austria | 2986 | 2419–3625 | 37.0–40.0 | 25.0–27.9 | 505 | Nitrogen and sulfur rates | [73] |
| Southern Ethiopia | 2956 | 2795–3200 | - | - | - | Seeding rates and nitrogen fertilizer | [74] |
| Saskatchewan, Canada | 2466 | 2184–2747 | 38 to 43 (39.7) | 27 to 32 (29.3) | 430 | Genotypes and environment | [39] |
| North–Eastern Poland | 2023 | 1700–2210 | - | - | - | Genotypes | [72] |
| Maritime Provinces of Eastern Canada | 1775 | 1638–1911 | 33.8–39.0 (36.6) | 25.0–26.8 (26.0) | 285 | Breeding lines, nitrogen and sulfur rates | [87] |
| Europe and Canada | 1660 | 1100–2700 | (41.8) | (26.2) | 305 | Genotypes and environment | [77] |
| Arizona, USA | 1583 | 1527–1638 | (45) | - | 313 | Nitrogen rates, water use and irrigation scheduling | [84] |
| Montana USA | 1349 | 546–2942 | 33.5–37.6 (36.0) | - | 211 | Cultivars and locations | [78] |
| Wyoming, USA | 1129 | 832–1643 | 31.1–32.4 | 29.3–30.4 | 157 | Nitrogen and sulfur rates | [91] |
| Chile | 991 | 387–2314 | 39.8–45.7 (41.41) | - | 180 | Cultivars, planting dates, and locations | [61] |
| Northern Italy | 820 | 600–940 | (39.2) | 141 | Low input and growing seasons | [71] | |
| Nevada, USA | 899 | 770–1013 | 31.8–33.3 (32.4) | 27.1–28.1 (27.5) | 128 | Cultivars and irrigation rates | [36] |
| 784 | 534–1010 | 31.8–32.6 (32.2) | 26.2–30.4 (28.4) | 111 | Cultivars, nitrogen sources and rates | [59] | |
| 735 | 34–1921 | 26.6–30.8 | - | 69.7 | Cultivars and year | [37] | |
| 570 | 130–921 | 28.4–29.5 | - | 72 | cultivars, sowing date, and methods | [62] | |
| Minnesota, USA | 812 | 650–944 | 39.4–40.7 (40.1) | 26.2–27.9 (27.0) | 143 | Genotypes and seeding rate | [3] |
| Kansas, USA | 427 | 317–503 | 27–29 (27.7) | 29–30 (29.7) | 52 | Cultivar and planting date | [75] |
| Overall mean | 1410 | 36.0 | 27.8 | 208.4 | |||
1 Oil and 2 protein content inside brackets indicate the average oil and protein content (%). 3 Biodiesel yield was estimated by multiplying seed yield (kg ha−1) and oil content (%) using the volumetric conversion factor of 1 kg ha−1 to 0.439 L ha−1 [92].
Compared with seed oil values, protein content generally varied less widely relative to the location of production across the globe, with the lowest and highest ranges of 25.0–26.8% and 27–32%, respectively, reported in Canada, with an overall mean of 27.8% (Figure 1, Table 1). However, more wide-ranging protein contents (23–47%) of C. sativa seeds were reported depending upon local field conditions [60,77]. High air temperatures during flowering and seed filling are generally associated with low oil contents but high protein contents within C. sativa seed [62]. Furthermore, increased N application rates can increase protein contents, with an associated decrease in oil contents [73,87,93,94]. Such increased nitrogen applications are known to increase protein at the expense of fatty acid synthesis due to the competition for carbon skeleton during carbohydrate metabolism [95].
A major use of C. sativa seed oil is biodiesel production. Biodiesel production, which is closely linked with overall oil production, varied widely with a mean of 208 L ha−1 across the globe (Figure 1, Table 1). The highest biodiesel yield (505 L ha−1) was reported in Austria [73], followed by western Canada (430 L ha−1) [39]. A multi-location study performed in Europe and Canada reported a mean biodiesel yield of 305 L ha−1 [77]. The lowest biodiesel yield (52 L ha−1) was reported in Kansas, USA [75]. The wide variation in biodiesel production is likely accounted for by differences in temperature and water availability, including ambient precipitation, length of growing season, and managerial factors such as fertilizer application rates and irrigation practices.
5. Biotic Production Constraints of C. sativa
Weeds are a major constraint to C. sativa production (Berti et al., 2016). C. sativa is considered to be very competitive against weeds with good stand establishment [22,96], due to its ability to produce and release secondary metabolites that prevent the growth of seedlings of neighboring plants [97,98,99]. However, the competitiveness of C. sativa for sunlight, nutrients, and water can be challenged by some weed species such as cheatgrass (Bromus tectorum), green foxtail (Setaria viridis L.), and Russian thistle (Salsola kali L.), among others [2,5,100,101]. Similarly, perennial broadleaf weeds such as field bindweed (Convolvulus arvensis L.), Canada thistle (Cirsium arvense L. Scop.), and skeleton weed (Chondrilla juncea L.) can also present challenges for C. sativa. Notably, C. sativa can exhibit susceptibility to dodder (Cuscuta spp.), a parasitic weed of many crops (D. Neupane, personal observation). C. sativa does not hybridize efficiently with cultivated Brassica species or wild Brassica species within the tribe Camelineae [102]. However, C. sativa does exhibit the ability to outcross with common Brassica weed species, such as North American weeds C. alyssum and C. microcarpa [103]. Fortunately, C. sativa is less competitive and shows lower invasive potential than Canola [104].
5.1. Weed Control
Both chemical and non-chemical approaches for weed control are available for C. sativa production systems. For chemical control, Sethoxydim (Poast®) (2-[(E)-N-ethoxy-C-propylcarbonimidoyl]-5-(2-ethylsulfanylpropyl)-3-hydroxycyclohex-2-en-1-one), a post-emergent and selective grass herbicide, which belongs to the cyclohexanone group, is currently the only registered herbicide for C. sativa in the USA [59,62,105,106]. Tepraloxydim (2-[(E)-N-[(E)-3-chloroprop-2-enoxy]-C-ethylcarbonimidoyl]-3-hydroxy-5-(oxan-4-yl)cyclohex-2-en-1-one) was also used to control post-emergent grass weeds [61]. Similarly, Clethodim (2-[(E)-N-[(E)-3-chloroprop-2-enoxy]-C-ethylcarbonimidoyl]-5-(2-ethylsulfanylpropyl)-3-hydroxycyclohex-2-en-1-one), which is a related post-emergent cyclohexanone herbicide, was successfully used for grassy weed control for C. sativa [36]. Quizalofop (2-[4-(6-chloroquinoxalin-2-yl)oxyphenoxy]propanoic acid), which is a selective, postemergence phenoxy herbicide registered in Canada and other parts of the world, was also used to control annual and perennial grass weeds of C. sativa [2,5,107].
In contrast to selective grass herbicides, C. sativa withstands very few broadleaf herbicides, and few have been approved for use on C. sativa in the USA [2,5,22,107,108]. Dinitroaniline pre-emergent herbicides, such as Trifluralin (2,6-dinitro-N,N-dipropyl-4-(trifluoromethyl)aniline), are widely used in the USA for early season weed control. Pre-emergent herbicides, including dimethenamid-P (2-chloro-N-(2,4-dimethylthiophen-3-yl)-N-[(2S)-1-methoxypropan-2-yl]acetamide), pendimethalin (3,4-dimethyl-2,6-dinitro-N-pentan-3-ylaniline), pyroxasulfone (3-[[5-(difluoromethoxy)-1-methyl-3-(trifluoromethyl)pyrazol-4-yl]methylsulfonyl]-5,5-dimethyl-4H-1,2-oxazole), quinclorac (3,7-dichloroquinoline-8-carboxylic acid), and S-metolachlor (2-chloro-N-(2-ethyl-6-methylphenyl)-N-[(2S)-1-methoxypropan-2-yl]acetamide), were tested at three different concentrations with C. sativa to examine weed control and injury to the crop [109]. Pendimethalin and S-metolachlor caused some injury, but did not lower C. sativa seed yields. Dimethenamid-P, used at a low rate, did not affect plant populations and seed yield; however, when applied at a high rate, it caused 60% injury with a 31% reduction in seed yield [109,110]. Quinclorac did not significantly injure the crop and was considered the safest of all the herbicides tested [109]. C. sativa mutants with resistance to acetolactate synthase (ALS) inhibitors were developed, displaying increased resistance to imazethapyr (5-ethyl-2-(4-methyl-5-oxo-4-propan-2-yl-1H-imidazol-2-yl)pyridine-3-carboxylic acid), sulfosulfuron (1-(4,6-dimethoxypyrimidin-2-yl)-3-(2-ethylsulfonylimidazo[1,2-a]pyridin-3-yl)sulfonylurea), and flucarbazone (3-methoxy-4-methyl-5-oxo-N-[2-(trifluoromethoxy)phenyl]sulfonyl-1,2,4-triazole-1-carboxamide) [110].
Several non-chemical strategies for weed control were shown to be effective for C. sativa [96]. Mechanical weeding is a well-established and useful approach for small scale production systems (e.g., research plots). For large-scale production, early season planting that promotes stand establishment is one of the most effective strategies for overcoming weed competition in C. sativa fields [111]. Furthermore, increasing sowing rates can effectively suppress weeds in C. sativa plots, likely through increased utilization of resources by the crop at the expense of weeds [96]. Mixed cropping of C. sativa grain with peas (Pisum sativum) (or barley (Hordeum vulgare)) afforded significant weed suppression and acted as a smother crop and weed antagonist, leading to enhanced C. sativa seed yields [96,112].
Use of C. sativa as a winter cover crop suppressed weed abundance while increasing cash crop yields [113]. Including C. sativa in crop rotations for winter cereals can suppress winter weed populations and increase grain yields in semi-arid Mediterranean climates [114]. C. sativa was also used in innovative double- and relay-cropping systems with forage sorghum (Sorghum bicolor), maize (Zea mays), and soybean (Glycine max) with an increased potential for biofuel and energy feedstock production [115]. When C. sativa was intersown as a cover crop with maize or soybean, establishment and winter survival rates increased, with the greatest success being achieved with soybean [116]. C. sativa was also used as an intercrop with Jatropha integerrima in semi-arid regions of India with seed production improvements observed when grown alternatively with leguminous fodder crops [117]. Further exploration of the potential allelopathic effects of C. sativa in intercropping systems for weed suppression is needed [99].
5.2. Insect Pests
Insects can limit C. sativa production, but these generally present lower production barriers than weeds. C. sativa is known to be relatively resistant to insect predation and infestation. C. sativa shows resistance to several common insect pests of Canola [118,119]. For example, C. sativa shows high resistance to flea beetles (Phyllotreta spp.), a common insect pest of B. rapa, B. napus, and B. juncea [39,49,120,121]. This resistance is thought to be due to the presence of repellents or the absence of stimulatory volatile phytochemicals [122]. More recent studies confirmed that the presence of quercetin glycosides in C. sativa leaves is likely responsible for flea beetle resistance [123]. In addition to flea beetles, C. sativa was shown to be a poor host for the diamondback moth (Plutella xylostella L.) and root maggots (Delia spp.) [124]. Feeding damage by the Bertha armyworm (Mamestra configurata Walker) on C. sativa was consistent with Canola, but larvae and pupae weighed less when reared on C. sativa leaves [124]. Leafhopper (Macrosteles quadrilineatus Forbes) did not cause damage to C. sativa; however, this insect can transmit aster yellows phytoplasma, which can result in chronic, systemic disease resulting in chlorosis, phyllody, and virescence, malformed seeds, and reduced seed set [124]. C. sativa also appeared to show resistance against the cabbage seedpod weevil (Ceutorhyncus obstrictus (Marsh.) [125]. However, stem feeding below the soil by several weevil species (e.g., Ceutorhynchus cyanipennis and C. americanus) was associated with stand failures for fall-planted C. sativa [126]. Although aphids can colonize C. sativa under field conditions, they are not known to cause significant economic damage; however, such colonization can create a reservoir for different aphid species that can negatively impact other crops in the rotation [127].
5.3. Fungal, Bacterial, and Viral Pathogens
Similar to its apparent resistance to many insect pests, C. sativa shows good resistance to a variety of microbial pathogens that typically target crops within the Brassicaceae [18,128]. For example, C. sativa showed strong resistance to blackspot fungal disease caused by Alternaria brassicae (Berk.) Sacc. [129,130,131]. Such resistance is associated with the relative expression of defense-related genes [132]. Resistance to blackspot disease caused by Alternaria brassicicola [(Schw.) Wiltsh] varies among C. sativa genebank accessions [133]. However, such resistance can be transferred to hybrids of C. sativa and rapid-cycling Brassica oleracea in an effort to confer resistance of this disease to Brassica vegetable crops [134,135]. Resistance to the blackspot pathogen was associated with the production of phytoalexins, specifically to camalexin, an antimicrobial phytoalexin [129,131,134,136]. Resistance was also correlated with the higher expression of chitinase gene family members in C. sativa relative to B. juncea [137]. C. sativa showed strong resistance to blackleg fungal disease [39] assessed using 80 different isolates of Leptosphaeria maculans (Desmaz.) [138]. However, for those C. sativa genotypes that are susceptible to blackleg disease, the fungicides metoconazole and a mixture of tebuconazole and triadimenol provided very effective control against the fungus [61].
C. sativa genotypes show various, but low, degrees of susceptibility to common Brassicaceae diseases, such as stem rot (Sclerotinia sclerotiorum (Lib.) de Bary), brown girdling root rot (Fusarium spp. and R. solani), downy mildew (Peronospora sparsa, P. parasitica), powdery mildew (Erysiphe sp.), grey mold (Botrytis cinerea), sore shin and damping-off disease (Rhizoctonia solani), Verticillium wilt (Verticillium dahliae), and white rust (Albugo candida) [18,128,139,140]. Some C. sativa genotypes are susceptible to downy mildew (Hyaloperonospora camelinae) [141]. Thirty diverse C. sativa genotypes showed varying degrees of resistance to S. sclerotiorum [142]. Resistance to S. sclerotiorum was associated with cell wall strengthening due to monolignol biosynthesis in C. sativa [143]. However, while camalexin production was induced by S. sclerotiorum inoculation, the relative degree of disease resistance was not correlated with levels of camalexin production, suggesting that other antimicrobial activities might be responsible for the observed disease resistance [142]. C. sativa also shows resistance to the soil-born fungus Fusarium virguliforme [144]. However, C. sativa is susceptible to other fungal diseases common to the Brassicaceae, such as damping-off (caused by Rhizoctonia solani), clubroot (Plasmodiophora brassicae Woronin.), and white rust (Albugo candida Pers. Kuntze) [128]. While susceptible, C. sativa showed greater resistance to Rhizoctonia solani than B. napus did, due to the presence of the phytoalexins camalexin and methoxycamelexin and two additional antimicrobial compounds in its roots [145].
C. sativa is susceptible to some bacterial diseases, such as bacterial blight (Pseudomonas syringae pv. spec.), phytoplasma disease, which was reported for C. sativa grown in Germany and Canada [139,140,146], and aster yellows diseases [124]. C. sativa also shows susceptibility to the bacterial pathogen Xanthomonas campestris Dowson pv. campestris, which causes black rot of Brassica crops globally [133].
Lastly, although relatively less well studied for viral diseases, C. sativa is susceptible to the Turnip crinkle virus and Turnip rosette virus [147], which are viral diseases transmitted by flea beetles via infested seed [2,148]. The beet western yellows mosaic virus reportedly caused up to 34% yield reductions in C. sativa [149]. Aphid (Myzus persicae) vector behavior and host palatability are key factors in the transmission of the Turnip yellows virus in C. sativa [150].
6. Ecosystem Services Provided by C. sativa
In addition to its role in crop rotations, C. sativa may be used to provide a range of ecosystem services, including prevention of soil loss and erosion, habitat for pollinators, and phytoremediation. Regrowth of hardy winter C. sativa can help to limit soil erosion and nutrient run off in the early spring [35]. In general, the use of C. sativa as a cover crop, with its deep tap root, is expected to benefit soil structure, promote nutrient recycling, and enhance nutrient scavenging [2]. Although C. sativa produces relatively small amounts of biomass compared with larger crops, these crop residues can promote soil water absorption capacity, which is particularly important in areas with dry soils [37]. Nonetheless, no-till and low-till cultivation methods are recommended for C. sativa to avoid or reduce soil erosion [111]. Both winter and spring C. sativa can provide pollinators with early season nectar and pollen for honey bees and other pollinators, as its flowers open long before most other crops grown in many regions [151,152]. Furthermore, C. sativa nectar sugar produced throughout anthesis can exceed that of pennycress (Thlaspi arvense) or Canola (Brassica napus) [151]. Cocultivation of winter C. sativa with Persian clover was found to decrease the heavy metal (i.e., copper, lead, nickel, and zinc) content of soils [153]. Lastly, C. sativa can be used similarly to other cover crops, including those within the Brassicaceae, to promote atmospheric carbon sequestration, suppress weeds, provide erosion control, protect ground water quality, promote organic matter carbon and nitrogen accumulation in soils, and suppress the accumulation of fungal pathogens, particularly saprophytic fungal species, and nematodes [154,155].
7. Fatty Acid Synthesis and Seed Oil Profiling in C. sativa
C. sativa seed oil content varies from approximately 27 to 46% and has a high omega-3 fatty acid content [26,39,49,64]. C. sativa seed oil contains 90% unsaturated fatty acids (~60% polyunsaturated fatty acids (PUFAs), 30% monounsaturated fatty acids (MUFAs)), and 10% saturated fatty acids (SFAs). In C. sativa oil, palmitic (C16:0), oleic (C18:1), linoleic (C18:2), and α-linolenic (C18:3) acids are the predominant fatty acids [156]. C. sativa seed oil is unique in its oil composition and chemical characteristics because polyunsaturated α-linolenic acid (C18:3) is the major fatty acid, and the concentration of erucic acid (C22:1) is present in a comparatively low amount for a Brassicaceae species, whereas eicosenoic acid (C20:1) is produced instead as the major long-chain fatty acid [6].
The mean fatty acid composition of C. sativa oil includes α-linolenic acid (34.4%), linoleic acid (18.3%), oleic acid (14.7%), gadoleic acid (C20:1, 14.0%), and palmitic acid (5.8%) in descending order of relative abundance (Table 2). The mean erucic acid content was 2.9%, which is below the maximum threshold value allowed for biodiesel (5%) [157], and just above the 2% threshold required for food-grade oil in the USA [3]. Other fatty acids such as stearic acid (C18:0, 2.6%), docosaenoic acid (C22:0), nervonic acid (C24:1), ecosadienoic acid (C20:2), dihomo-gamma linolenic acid (C20:3), and docosadienoic acid (C22:3) are also present in trace amounts. The relative content of the major fatty acids varied widely depending on the study. For example, α-linolenic acid ranged from 22.8–38.4% [158] to 41% [157]. Linoleic acid ranged from 12.8–20.6% [159,160,161]. Oleic acid ranged from 6.9–22.1% [158]. The ranges of other fatty acids, such as gadoleic acid, palmitic acid, and erucic acid, are summarized (Table 2). The wide variations in fatty acid profiles of C. sativa likely arise from differences in genotypes, agronomic practices, growing environment (e.g., soil quality), weather and climatic conditions [2,90,162,163].
The biosynthesis of triacylglyceride (TAG) in C. sativa occurs in the plastid via a Type II fatty acid synthase complex [169]. In most oilseed crops, the fatty acid chain elongates to 16 or 18 carbons in length [162]. Oleic acid (C18:1) (MUFA) is synthesized in the plastid from stearic acid (SFA) through steroyl-acyl carrier protein desaturase (SAD). Further desaturation of oleic acid to linoleic acid (C18:2) is catalyzed by fatty acid desaturase-2 (FAD2), a key enzyme responsible for the biosynthesis of PUFA in non-photosynthetic tissues, for example, the roots and developing seeds of oilseed plants in the ER and FAD 6 (plastidial enzyme) in the plastid [170,171]. α-linolenic acid (C18:3) synthesis from linoleic acid is catalyzed by FAD3 (microsomal enzyme) in the ER and FAD7/FAD8 (plastidial enzymes) in the plastid [172]. Fatty acid composition can change during seed development and varies with C. sativa accession. Fatty acid production can be affected by location due to climactic differences [157,173]. In oilseed crops, α-linolenic acid content differs depending on the temperature conditions during seed development. For example, at temperatures greater than 25 °C, the synthesis of α-linolenic acid declines because the activity of FAD3 decreases [2], whereas the synthesis of oleic and α-linoleic acid increases [174,175]. Similarly, biosynthesis of α-linolenic acid declined in non-irrigated compared with irrigated C. sativa, likely due to the higher temperatures of the non-irrigated plants [81]. Additionally, increasing temperatures from 15 °C to 35 °C reduced the expression of the FAD2 gene, suggesting a reduction in the synthesis of linoleic acid from oleic acid. A shorter duration of grain filling and a greater number of days above 25 °C resulted in a decline in linoleic acid content [176]. More recently, water-deficit stress was shown to increase oleic and linoleic acid content in C. sativa seeds, whereas linolenic acid content decreases, being dependent upon genotype, as one accession showed an increase in a-linolenic content following drought treatment [177]. Importantly, various environmental stress factors including heat, drought, salinity, high light, low oxygen, and high nitrogen can lead to decreases in overall seed oil content and changes in fatty acid composition [162]. Thus, an important consideration for improving overall seed oil yield is the need to improve the abiotic stress tolerance of C. sativa, particularly in the face of increasingly hot and dry conditions brought about by global warming.
8. Uses of C. sativa
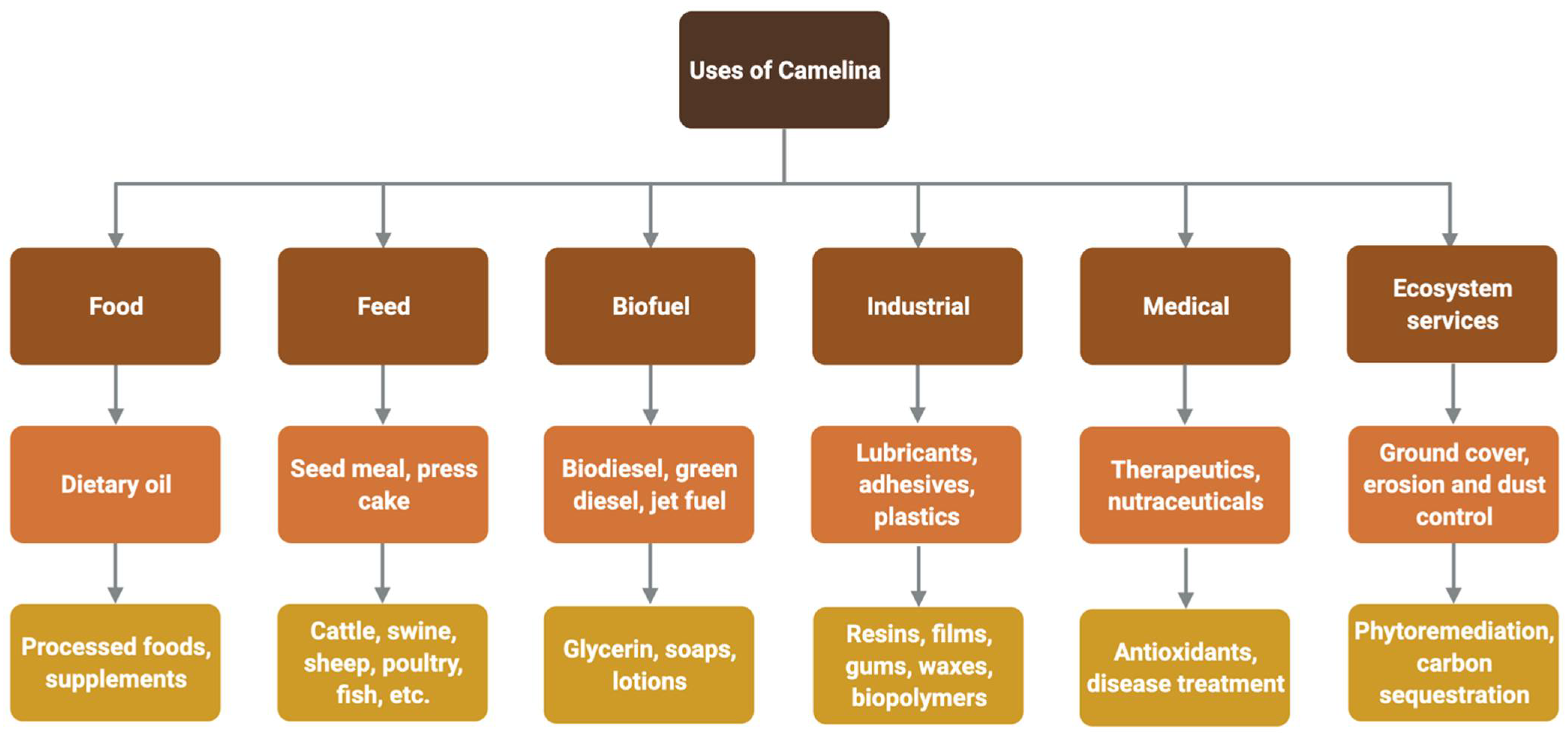
C. sativa is used in a wide array of products and applications ranging from industrial, biomedical and nutraceutical products to animal feed, erosion control as ground cover, phytoremediation and carbon sequestration. C. sativa is also used in processed foods for human consumption. C. sativa is widely used as a biofuel, including biodiesel, green diesel, and renewable jet fuel. Derived products include glycerin, soaps and lotions (Figure 2).
8.1. Food and Food Products for Humans
C. sativa cooking or salad oil has potential human health benefits due to its high levels of PUFAs, omega-3-fatty acids, and antioxidants [156,178]. The consumption of C. sativa oil can reduce blood serum cholesterol levels [179] and improve serum lipid profiles [180,181]. Furthermore, C. sativa oil consumption can attenuate inflammation in peripheral blood mononuclear cells [182]. Such metabolic changes are thought to not only protect against cardiovascular risk factors, but also improve mental health in patients suffering from non-alcoholic fatty liver disease [183]. C. sativa oil was also used in folk medicine to treat burns and wounds to the skin and eyes [184,185]. The high levels of tocopherols, phytosterols, and carotenoids in C. sativa oil protect it from oxidation, imparting extended shelf life [185,186]. C. sativa oil displays antioxidant activities similar to those found in sunflower oil, which can effectively limit oxidation in food products, such as salad dressings and mayonnaise [156,187]. For example, C. sativa oil and rapeseed meal prevented the oxidation of lipids and proteins in cooked pork patties [188].
8.2. Feed for Animal Nutrition
Following oil extraction, C. sativa seed meal is toasted, dried, and cooled ready to use as animal feed for cattle, dairy cows, sheep, swine, and poultry, as well as for aquaculture of various fish stocks [2,189]. C. sativa seed and seed meal are generally considered useful and beneficial as animal feed in limited quantities. C. sativa seed displays a balanced profile of essential and non-essential amino acids [159,190].
8.2.1. Beef Cattle
The exceptionally high level of omega-3 fatty acids, particularly α-linolenic acid (~32–40% of total oil content), protein (40%), oil (10–15%), fiber (0–15%), and phytate (1–6%), combined with relatively low glucosinolate levels (20–44 mmoles kg−1 dry weight), in C. sativa seed meal make it a suitable feed for animals [2,21,159,160,191,192]. C. sativa seed meal is generally comparable to Canola meal in terms of its amino acid profile, crude protein content, and digestibility, and was demonstrated to be a valuable feedstuff for ruminants [193,194,195]. However, some researchers report lower degradability and palatability of C. sativa seed meal compared with soybean or Canola meal [196,197]. Supplementation of the diet of beef cattle with C. sativa seed meal resulted in reduced forage and dry matter intake, but gave rise to greater serum PUFA concentrations [198]. In a related study, addition of 10% C. sativa seed meal on a dry weight basis did not affect dry matter intake and improved digestion of rumen organic matter, but did reduce body weights of dairy heifers [199]. Other studies report no negative effects on animal performance when C. sativa seed meal comprised up to 10% of the total diet [193]. C. sativa seed meal is highly degradable and shows comparable total digestibility and protein absorption to other cattle feed supplements [200]. Reduced intake or weight loss might be the result of the reduced palatability of C. sativa seed meal, as it contains anti-nutritive compounds, such as erucic acid, sinapine, and glucosinolates [196,201]. Due to these reasons, the U.S. Food and Drug Administration (FDA) permits only a maximum of 10% C. sativa seed meal in rations for livestock [202]. However, the sinapine content of C. sativa seed meal is lower than other Brassica species, and thus, only the glucosinolate content is relevant when assessing palatability [192]. Glucosinate content varies widely among accessions, and thus could be reduced through conventional breeding [192,203,204]. A high erucic acid content in seed meal can lead to fat deposition and reduced contractibility of the heart muscle and is thus limited to a maximum of <2% [205]. The erucic content in C. sativa seed meal ranges from 1.8–4.8% in the USDA National Genetic Resources Program collection [161]. Thus, similar to low glucosinolate accessions, accessions with low erucic acid content, such as C. sativa accession PI 650141, could be used for conventional breeding efforts or genome-editing approaches [161].
8.2.2. Dairy Cows
C. sativa meal has a reasonably high level of histidine content, which is a good supplement for silage and grain-fed lactating cows [196]. Inclusion of seed or seed meal in the diet of dairy cows decreased dry matter intake, but did not have a significant effect on milk production [206]. However, overall milk protein and fat yield declined and resulted in a modified fatty acid profile with increased MUFAs and PUFAs, with resultant increases in butter spreadability [206]. Addition of C. sativa seed meal or oil to dairy cow diets did not alter dry matter intake, digestibility, or milk yield, but resulted in decreased saturated fatty acids and increased milk MUFA and PUFA content [207]. In a related study, supplementation of up to 6% C. sativa oil decreased milk yields and saturated fatty acid content while increasing unsaturated fatty acid content. This study concluded that 2% C. sativa oil could be used for commercial production without major adverse effects on animal performance [208].
8.2.3. Sheep and Goat
Inclusion of 10–20% C. sativa seed meal in the diet of sheep increased the content of linoleic, oleic, and α-linolenic acid content in lamb muscle [209] and reduced blood triglycerides and glucose content, while increasing insulin levels [210]. Inclusion of C. sativa seed meal in sheep diets improved the total omega-3 fatty acid and the ratio of omega-6/omega-3 fatty acids in lamb and yearling meat, but resulted in reduced vitamin E content, which negatively impacted color and oxidative stability upon storage [211].
Supplementation of an ewe’s diet with 3–6% of C. sativa seed meal (or seed) in dietary dry matter increased the mono- and polyunsaturated fatty acid content of milk, resulting in milk with lower atherogenic and thrombogenic indices [212,213]. Similarly, inclusion of C. sativa seed meal in the diet of dairy goats increased total polyunsaturated fatty acids and decreased total saturated fatty acids in milk and kefir produced from the milk [214]. Inclusion of 10–20% C. sativa seed meal in the diet of dairy sheep altered the content and aroma of volatile compounds, primarily fatty acids, in raw and pasteurized ewe’s milk with an overall loss of dairy or freshness aroma [215]. Sheep or goat milk with increased fatty acid content resulted in increased fermentation time and alterations in the resulting kefir, and altered its aroma [216]. Additional studies validated these effects and showed that supplementation of sheep diets with varying amounts of C. sativa seed meal increased the mono- and/or poly-unsaturated fatty acids content and oxidative stability of the milk, resulting in a healthier milk for human consumption [217,218,219,220].
8.2.4. Swine
After cold or expeller pressing, a comparatively high amount of oil (100–150 g kg−1) remains in the meal of C. sativa, which provides a potential energy source for swine diets [2]. Inclusion of 5–10% C. sativa seed meal in swine diets resulted in increased plasma omega-3 fatty acids and reduced plasma omega-6 fatty acids in blood plasma, while reducing serum triglyceride levels [221]. During the finishing–fattening process, supplementation of the swine diet with C. sativa oil increased α-linolenic acid content and reduced cholesterol content in the meat, thereby improving meat quality [222], and increased n-3 polyunsaturated fatty acid and reduced cholesterol and triglyceride of the plasma, implicating improved animal health [223]. Compared with sunflower seed meal, the inclusion of 12% C. sativa seed meal in the swine diet did not affect growth performance, but elevated the expression of some antioxidant defense system components in the spleen [224]. Digestibility of C. sativa seed meal in swine was comparable to that of Canola meal [225]. However, the addition of 20% C. sativa seed meal lowered digestibility coefficients for amino acids and crude protein, which limits its utilization in the swine diet [226]. Inclusion of 12% C. sativa seed meal increased α-linolenic acid content in the heart and brain with little alteration in performance [227]. Inclusion of C. sativa seed meal in the diets of weaned piglets at up to 18% increased n-3 fatty acids in carcass fat depots, but did not elicit clinical signs of toxicity, although it did result in lower average daily feed intake and weight gain than controls, probably due to an aversion to the taste of the C. sativa seed meal [228].
Supplementation of C. sativa seed meal improved feed efficiency and liver weight in 28-day-old weaned piglets, but also increased the expression of selected liver enzymes [229]. Comparison of the effects of the addition of 5%, 10%, or 15% C. sativa seed meal to the diet of growing–finishing pigs showed that the two higher percentages reduced average daily weight gain and marketing weight, but no negative growth performance or carcass traits were observed at 5% addition to a corn–soybean meal-based diet [230]. Incorporation of up to 30% C. sativa seed meal in a mixed corn/soybean meal diet for growing swine did not alter the digestible, metabolizable, or net energy, suggesting that dietary glucosinolates from C. sativa seed meal did not affect these dietary parameters [231].
8.2.5. Poultry
Incorporation of C. sativa seed meal into poultry diets benefits from the provisioning of energy and protein, increased health-promoting PUFAs and tocopherol content of meat and liver, improved antioxidant activity and lipid stability, and increased market value of the meat and eggs [189,232]. Unlike other livestock animals, chickens appear somewhat less susceptible to the taste of C. sativa seed meal and generally do not show reduced feeding efficiency or weight gain when provided in moderation. For example, the addition of 2.5%, 5%, and 10% C. sativa seed meal to a basal corn–soybean diet had little effect on weight gain and feed efficiency [233]. However, C. sativa seed meal resulted in significant increases in a-linolenic acid and n-3 PUFAs in white and dark meat and tissues [233]. Inclusion of 10% C. sativa seed meal increased egg production and increased α-linolenic acid and total n-3 PUFAs in egg yolks, compared with a corn–soybean-based diet, but reduced crude protein digestibility [234]. Addition of 8%, 16%, and 24% C. sativa seed meal resulted in linear increases in α-linolenic acid content in various tissues including liver, breast, and thigh, and in the proportion of n-3 PUFAs in liver and brain [235]. The supplementation of cold-pressed C. sativa seed cake of broiler chicken diets with or without multienzyme supplementation to improve digestibility was found to improve the energy and amino acid content of the diets [236].
Related studies tested the effects of oil supplements on poultry diets. For example, the addition of 6% of C. sativa oil to feed rations for broiler chickens reduced the cholesterol content in the blood plasma and increased omega-3 PUFA content in the breast, without producing an unpleasant flavor [237,238]. The addition of C. sativa oil to broiler chicken diets resulted in increased α-linolenic acid content in muscle and abdominal fat to a greater extent than diets supplemented with either soybean or rapeseed oil [237]. Other studies showed that C. sativa oil and seed meal in broiler chicken diets improved the percentage of PUFAs, particularly omega-3, and lowered the percentage of MUFAs, such as oleic acid, in the lipids of the breast muscles [239].
In addition to chicken, a variety of other poultry species were evaluated for C. sativa seed meal use. As with chicken diets, the inclusion of 15% to 20% C. sativa seed meal in duck diets resulted in significant increases in α-linolenic acid and total omega-3 PUFA in breast and leg muscles [240]. The addition of up to 5% C. sativa seed meal to the diets of young turkeys showed similar weight gain and feed conversion as vegetable oil and showed no negative effects [241]. However, the addition of 5% and 10% C. sativa cake to turkey diets caused growth depression and reduction of feed intake [242,243]. In contrast, no negative effects of weight gain or feed intake were observed when 5%, 10%, or 15% C. sativa seed meal was added to quail diets [244].
8.2.6. Fish
C. sativa oil and seed meal can serve as good replacements for fish oil and fish meal in fish feed. Diets of 5% C. sativa seed meal are acceptable substitutes to reduce reliance on fish meal [245]. Substitution of C. sativa oil in fish feed improved the total lipid content in salmon (Salmo salar L.) and Atlantic cod (Gadus morhua L.) without negatively affecting the sensory quality of fish fillets [246]. C. sativa seed oil can be used as up to 100% replacement for fish oil without negative growth effects for Atlantic salmon raised in freshwater [245]. However, while replacing fish oil with C. sativa oil at ~80% had no effect on growth performance, PUFA content in the liver and muscle was reduced in Atlantic cod [247]. In another study with Atlantic cod, feeding C. sativa seed meal up to 24% of the diet did not affect growth in one experiment, while in a separate experiment, C. sativa seed diet greater than 30% resulted in depressed feed intake and growth due to reduced palatability [248]. Replacement of fish oil with oil from C. sativa genetically modified to express eicosapentaenoate (EPA; 20:5 n-3) and docosahexaenoate (DHA; 22:6 n-3) [249], in a feeding trial of European sea bass (Dicentrarchus labrax), showed no change in feed intake or growth [250]. However, the resulting fillets showed enhanced omega-3 PUFA content, indicating that C. sativa seed oil can effectively replace fish oil for the production of European sea bass without negative impacts on feed efficiency or growth rates [250]. C. sativa seed or seed meal of at least 10% can be used for feeding rainbow trout (Oncorhynchus mykiss) without reductions in feed conversion or weight gain [251].
8.3. Biodiesel/Renewable Diesel Fuel
One of the most important uses for C. sativa oil is the production of various biofuels. As with other oilseeds, the fatty acid profile of C. sativa oil varies depending upon the environmental conditions under which it is grown, thus resulting in variations in physical and chemical properties of the biodiesel produced. The major properties used to evaluate the quality and performance of biodiesel include cloud point (CP), cold-flow characteristics or viscosity, cetane number (CN), and oxidative stability [2,26]. Unformulated C. sativa B100 fails to meet the American Society for Testing and Materials (ASTM) standards for CP and oxidative stability due to exceptionally high (48–50%) polyunsaturated fatty acid content, but does meets the standard criteria for CN. However, with the addition of antioxidants, unformulated B100 can meet current ASTM standards while maintaining production costs at a level comparable with other biodiesel feedstocks [165,252]. Biodiesel produced from C. sativa oil generally resembles biodiesel from Canola oil (B. napus) [25]. The kinematic viscosity of methyl esters obtained from C. sativa varies from 2.9–6.4 mm2 s−1 at 40 °C, which lies within the acceptable 1.9–6.0 mm2 s−1 ASTM D651, and 3.5–5.0 mm2 s−1 EN14214 standard values [25,26,165,166,253,254,255]. The CN for C. sativa methyl ester ranged from 49.26 to 51.17, a value higher than ASTM biodiesel standards, resulting in good ignition quality of fuel. Furthermore, the pour point of C. sativa methyl ester was typically between −11 °C and −8 °C, suggesting a good quality fuel under cold season temperatures. Fuel consumption and vehicle operation from C. sativa and Canola methyl esters are similar [25]. In addition to traditional biodiesel, C. sativa can be used to produce hydrogenation-derived renewable diesel (HDRD) or green diesel, which generally has chemical properties more similar to that of petroleum diesel and improved cold-flow characteristics [256], yet is more sustainable than fossil diesel based upon emission parameters [257]. Life cycle analysis of C. sativa for biodiesel production showed that it produces lower greenhouse gas emissions than soybean- or Canola-derived biodiesel through reduced impacts on land-use changes [258].
8.4. Hydro-Processed Renewable Jet (HRJ) Fuel
Renewable jet fuel production from C. sativa oil follows a two-step standard process: (1) initial hydrodeoxygenation or hydrotreatment and (2) selective cracking or hydrocracking and isomerization, followed by product separation and formulation. The linear alkanes produced during step one can be used in renewable diesel mixtures [2,166]. Compared with JP-8 (typical jet fuel), C. sativa HRJ fuel had superior thermal-oxidative stability [259]. Engine tests with C. sativa fuel showed no obvious anomalies with engine operation, with the added advantage of lower carbon monoxide emissions than JP-8. However, C. sativa HRJ fuel contains elastomer seal swelling capability, which might cause fuel leaks in aircrafts, but this was lower than that of JP-8 [259]. C. sativa HRJ fuels have similar properties to conventional fuels used in turbine engines [259,260,261]. In 2009, the U.S. Air Force (USAF) tested the potential of C. sativa HRJ fuels as a replacement for jet fuels, with successful tests performed in fighter jets, private jets, and commercial airlines using a blend of JP-8 and C. sativa-based jet fuels [2,262]. C. sativa-based jet fuel with blending had a 75–80% lower carbon footprint compared with conventional fuel [263]. Overall, these results confirm the ability of renewable fuels derived from C. sativa to burn cleaner than conventional fossil fuel resources, while also removing carbon from the atmosphere during their production, resulting in overall lower carbon emissions. However, techno-economic analysis reveals that considerable investments in land area would be needed to grow C. sativa at large scale for economic viability [264].
8.5. Fast Pyrolysis Bio-Oil
In addition to the use of C. sativa for biodiesel and renewable diesel, the energetic value of the seed meal and lignocellulosic biomass was also considered. Thermochemical fast pyrolysis of the seed meal after oil extraction contains significant energetic value (29 MJ kg−1) when compared with whole seeds (34.7 MJ kg−1) and can provide additional volumes of high-energy, high-carbon liquid fuel intermediates for the production of renewable biofuels and jet fuel [265]. C. sativa residual biomass is comprised of 36.3–41.6% cellulose, 23.2–24.9% hemicellulose, and 25.0–26.3% lignin, and can be converted to bio-oil using thermal or catalytic fast pyrolysis [266]. However, the energetic value of C. sativa straw (15.1–15.5 MJ kg−1) or straw pellets (16.6–17.1 MJ kg−1) is lower than that of seeds or seed meal, but can produce gas, solid, and liquid fuel components with reduced nitrogen and sulfur contents relative to other woody or herbaceous feedstocks tested [267]. C. sativa straw was also used for ethanol production with an estimated total energy yield of 54.3 MJ L−1 ethanol [268]. Agronomic management can improve the overall energy potential of C. sativa biomass. For example, high rates of nitrogen and sulfur fertilization can increase the energy output of C. sativa seed and residual biomass by up to 186% and 155%, respectively [269]. The net energy efficiency and economic value of total C. sativa biomass (straw and seed) was higher than that of crambe (Crambe abyssinica) [270]. Thus, C. sativa biomass utilization, as part of an integrated biorefinery strategy where renewable diesel or jet fuel and thermal or catalytic fast pyrolysis bio-oil production are co-localized, can improve the overall energetic value of this bioenergy resource.
8.6. Industrial Applications
C. sativa oil has great potential for bio-based industrial applications. The high quantity of unsaturated fatty acids (~90%) present in C. sativa oil [6], enables rapid drying, and is useful for making dermatological products, cosmetics (lotions and soaps), polymers, paints, and varnishes [22,27,271]. A key limitation of using C. sativa oil for industrial application is its high content of PUFA, which promotes the oxidation of oils upon exposure to high temperatures [272,273]. C. sativa oil can be epoxidized and used for making adhesives, coatings, lubricants, and alkyd resins [274,275]. C. sativa meal has great potential for use in the paper industry [276]. The addition of C. sativa meal to recycled newspaper produced sustainable and bio-degradable green composite sheets and fibers [276]. Other applications include bio-herbicides [277], soil fungicides [278], and bio-oils [265]. C. sativa oil contains glucosinolates, which can be used to produce ionic thiocyanates that can serve as effective bio-herbicides against redroot pigweed (Amaranthus retroflexus L.) and wild oat (Avena fatua L.) [277]. C. sativa glucosinolate content ranged 9–31.4 µmol g−1 in seed [279,280], 14.5–23.4 µmol g−1 in seed meal [280], and 8.6–30.5 µmol g−1 in oil [281] depending upon the genotype and environmental growth conditions.
8.7. Biomedical Uses
In addition to the beneficial use of C. sativa seed meal for improving the quality of other foods such as meat and dairy products, the consumption of dietary oils, such as the oil from C. sativa, which is rich in essential omega-3 fatty acids (e.g., α-linolenic acid), was associated with reduced risks of coronary heart disease and inflammatory diseases [282,283]. Supplementation of the human diet with C. sativa oil can also reduce serum cholesterol in hypercholesterolemic subjects [179]. In addition, the oil contains high nutritional value due to the presence of tocopherols, phytosterols, and carotenoids [284,285].
Leaves of C. sativa contain flavonols such as quercetin [286]. Methanolic and ethanolic extracts of C. sativa seeds show significant antibacterial and antifungal activities and potential as natural preservatives [287]. Additional therapeutic properties are associated with phenolic compounds in C. sativa seeds and oil, such as chlorogenic, caffeic, sinapinic, and phytic acids [288]. However, as mentioned earlier, C. sativa seeds contain glucosinolates, which have antinutritive properties [201].
The ethanolic and methanolic extracts of defatted C. sativa seeds were shown to reverse short-term memory impairment and reduce anxiety and depression-like behaviors in a Swiss mouse model for irritable bowel syndrome exposed to various stress tests [289]. Furthermore, these extracts decreased superoxide dismutase, but increased glutathione peroxidase activity in brain and bowel tissues as well as potentially increasing lipid peroxidation in the bowel. The authors concluded that seed extracts could improve performance and mood while exhibiting antioxidant capacity in both brain and bowel tissues [289]. In a related study, the administration of cold-pressed C. sativa oil was shown to ameliorate impairment of short-term memory, anxiety, and depression in a Swiss mouse zymosan-induced model for irritable bowel syndrome exposed to various stress tests [290]. Furthermore, C. sativa oil treatment resulted in increases in superoxide dismutase and glutathione peroxidase activity in brain and bowel tissues, resulting in decreased malondialdehyde levels, a lipid peroxidation marker, in these tissues [290].
9. Strategic Innovations for Climate-Resilient C. sativa
The hotter and drier conditions brought about by climate change will require novel strategies for improving the heat and drought durability of C. sativa in the future. Recent research suggests that C. sativa is able to maintain a higher shoot/root ratio under water-deficit stress and is more drought tolerant than Canola (B. napus) [291]. Comparisons of C. sativa and Canola under deficit irrigation regimes suggest that C. sativa performed better and gave higher seed yields than Canola under field conditions [292]. To assess the relative tolerance to water-deficit and salinity stress, screening assays of different genotypes at germination were developed. Such germination screens suggested that seedlings of C. sativa cv. Calena exhibited germination up to 150 mM NaCl and thus might be suitable for cultivation in saline soils [293], but additional research is required to confirm this suggestion. Comparison of two different genotypes of C. sativa showed that increased water-deficit stress tolerance was associated with differences in stomatal conductance and increased accumulation of osmoprotectants such as proline, sugars, amino acids, and soluble proteins [294]. Screening of winter and spring genotypes revealed differences in germination under artificial osmotic stress imposed by different concentrations of polyethylene glycol (PEG) [295]. Application of sodium selenite (Na2SeO3) via seed priming or foliar application was shown to increase drought tolerance in C. sativa (and Canola) via the accumulation of osmoprotectants and antioxidants, and to improve crop yields [296,297]. Fertilization of C. sativa with thiourea at the seed filling stage improved heat tolerance [298]. While such selection or treatment efforts carry a certain degree of practical utility, more direct advanced breeding and biotechnological innovations will be needed to dramatically improve the yield potential and climate resiliency of C. sativa.
9.1. Molecular Breeding Approaches
Characterizing the genetic diversity and population structure of available genotypes is often the first step towards developing breeding programs for improving key traits such as yield potential, oil content, resistance to biotic and abiotic stresses, and adaptability to diverse environments. Sequencing complete allohexaploid C. sativa transcriptomes and genomes was a key first step for advanced molecular breeding efforts [34,53,299,300]. Such information is critically important for future gene identification using genomic selection, marker-assisted selection, and genome-wide associations studies (GWAS) to guide future C. sativa breeding programs. For example, a recombinant inbred population from two C. sativa accessions (Suneson and Pryzeth) with contrasting traits, especially seed size and oil content, was developed and used to assess phenotypic differences under two environmental conditions (dryland and irrigated) [301]. Using 189 lines from this population, a genetic map was created containing 2376 single nucleotide polymorphisms (SNPs) to identify quantitative trait loci (QTLs) associated with oil content, seed size, pod size, and seed number per pod as a first step towards isolating genes that control for seed development and oil accumulation [301]. In another example, a total of 213 spring C. sativa accessions were collected and genotyped to assess the genetic diversity available for future breeding efforts [302]. A total of 6192 SNPs were identified using genotyping-by-sequencing technology and used to reveal two distinct populations of C. sativa arising from Germany and eastern Europe [302].
Various screening efforts for wildtype and mutagenized C. sativa were used to identify lines with desirable traits of interest. For example, analysis of C. sativa seedlings from 10 different cultivars using germination screens under a gradient of NaCl conditions found that 100 mM NaCl was optimal for differentiating the phenotypic performances of seedlings [303]. Screening of a spring panel of 211 C. sativa accessions followed by GWAS revealed a total of 17 significant trait-associated SNPs for germination rates and dry weight with potential roles in root development through mediation of phosphate metabolism, signaling, and cell membrane activities [303]. In addition, mutational breeding using gamma-ray irradiation was successfully used to alter the fatty acid profile in C. sativa, resulting in the identification of M2 mutants with increased α linolenic acid content or decreased erucic acid content [304]. Screening of ethyl methanesulfonate (EMS) mutant populations resulted in the isolation of a C. sativa mutant with reduced seed coat mucilage, which could potentially improve the flow characteristics of the oil and reduce the washing requirements of the resultant biodiesel [305].
9.2. Improving Productivity Using Rhizosphere Microbes
In order to reduce reliance on fossil fuel-based fertilizers, interest into the use of plant growth-promoting rhizobacteria has increased in recent years. Early research indicated that the presence of nitrogen-fixing bacteria was necessary to stimulate root growth in flax (Linum usitatissimum L.) through aqueous washing of C. sativa leaves [306]. More recently, inoculation of C. sativa with a consortium of rhizosphere soil bacteria containing Bacillus species resulted in approximately a threefold increase in seed yield [307]. Similar enhancement in C. sativa winter hardiness and seed productivity were observed when C. sativa seeds were inoculated with nodule bacteria from crimson clover (Trifolium incarnatum L.) roots before sowing [308].
9.3. Improving Quality Traits
To complement molecular breeding and rhizosphere manipulation approaches, advances in the genetic transformation of C. sativa were developed [9,10,12,13]. The stable introduction of genes into C. sativa was used to modify or improve a wide range of agronomic traits with a focus on the manipulation of seed oil yield potential and characteristics [11]. For example, C. sativa was genetically modified using a set of heterologous genes to express eicosapentaenoate (EPA; 20:5 n-3) and docosahexaenoate (DHA; 22:6 n-3) as a replacement for fish oil [249]. Improving the oil content and seed yield in C. sativa by co-expressing A. thaliana diacylglycerol acyltransferase 1 (DGAT1) and yeast cytosolic glycerol-3 phosphate dehydrogenase (GPD1) genes under the control of seed-specific promoters resulted in up to 13% higher seed oil content and up to 52% higher seed mass compared with wild-type plants [309].
9.4. Improving Stress Tolerance Traits
Soil amendments containing PGPR (Pseudomonas migulae 8R6) that produce 1-aminocyclopropane-1-carboxylate deaminase (ACC deaminase) increased seed production in C. sativa by 30–50% under salinity stress by reducing stress ethylene [310]. Furthermore, plants expressing the bacterial version of the ACC deaminase (acdS) gene showed higher seed production, better seed quality, and higher levels of seed oil production under salinity stress than control lines [310]. The observed improvements in salinity tolerance were attributed to changes in the gene expression of multiple plant signaling pathway components and the modulation of genes involved in the production of reactive oxygen species (ROS) scavenging and signaling [311].
Many engineering strategies targeting improvement of abiotic stress tolerance begin with the characterization of stress-responsive genes. For C. sativa, stress-responsive gene discovery was performed by subjecting 15-day-old plants to 3 or 17 days of water-deficit stress and rehydration recovery for 3 days to identify hundreds of differentially expressed genes in diverse metabolic and signaling pathways based upon their relative, steady-state transcript abundance patterns [312]. Gene expression changes in C. sativa roots and vegetative tissues in response to salinity stress revealed hundreds of changes in relative, steady-state transcript abundance, the relative proportion of which could be assigned to each of the three C. sativa subgenomes [313]. Ectopic overexpression of a MYB96 transcription factor from A. thaliana in C. sativa resulted in improved water-deficit stress tolerance, due to the activation of genes involved in epicuticular wax biosynthesis [314]. Similarly, the ectopic overexpression of the CsMYB96A gene resulted in increased water-deficit stress tolerance due to the increased expression of epicuticular wax biosynthetic genes [315]. Such an approach might be useful for growing C. sativa on semi-arid and arid lands; however, field trials would be necessary to assess the efficacy of these modifications.
9.5. Genome-Editing Approaches
To complement molecular breeding approaches, advances in genetic transformation and genome editing using clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein (CRISPR/Cas) are used to improve the oil contents and fatty acid profiles in oilseed crops, including C. sativa [316,317,318]. An early demonstration of this technology in C. sativa was the simultaneous mutation of the diacylglycerol acyltransferase (DGAT1) and phospholipid:diacylglycerol acyltransferase (PDAT1) genes, which resulted in decreased oil content and altered fatty acid composition [319]. CRISPR/Cas9 disruption of all three homoeologous fatty acid desaturase 2 (FAD2) genes in C. sativa resulted in a substantial increase in oleic acid content (15% to >50%) with associated decreases in linoleic (~16% to <4%) and α-linolenic acid (~35% to <10%) content in C. sativa seeds [16]. In a related study, disruption of all three pairs of FAD2 homeologs in C. sativa resulted in an 80% enhancement in MUFA content in seeds, but this resulted in severely stunted plants [319]. However, if only two homeologous gene pairs were knocked out, up to a 60% increase in seed MUFAs was obtained and the plants showed normal phenotypes [319]. This alteration in the fatty acid profile resulted in a healthier oil profile while also improving the oxidative stability of the oil, which improves its utility for biodiesel production [16]. A related study used CRISPR/Cas9 to knock out all three FAD2 homeologs in the allohexaploid C. sativa, resulting in increased accumulation of oleic acid in the seed oil [15]. CRISPR/Cas9-mediated knockout mutagenesis of all three fatty acid elongase 1 (FAE1) genes reduced the total content of C20-C24 very long-chain fatty acids (VLCFAs), which normally constitute 22% of fatty acids in wild-type plants, to less than 2% of total fatty acids, with a corresponding increase in desirable C18 unsaturated fatty acids for dietary or fuel uses [14]. CRISPR/Cas9 gene editing was also used to disrupt the three homeologous genes encoding CRUCIFERIN C, which resulted in an alteration in the relative abundance of other cruciferin isoforms, but did not alter total seed protein content [320]. However, the overall amino acid composition of the seed was altered along with an increase in the relative abundance of all saturated fatty acids. Other attractive targets of genome editing to improve production traits include strategies to reduce silicle shattering and to prevent uneven seed maturation.
10. Conclusions and Future Directions
C. sativa is a highly attractive oilseed due to its low input requirements and costs and adaptability to diverse environments and soil conditions, including abandoned, marginalized, and semi-arid and arid regions. While genetic engineering and genome-editing approaches have yielded great improvements in its oil content and fatty acid profiles, C. sativa can also be outcrossed with related species [103], raising the possibility of targeted breeding programs to improve its yield and trait diversity. Improvements in agronomic management and commercial investments are also needed to reduce production costs and inputs and improve its overall seed yield and seed oil content relative to competing oilseeds [1,258]. In addition, improved valorization of the C. sativa value chain, from seed processing, oil extraction and refinement methods [321] to its co-products and lignocellulosic crop residues, is needed to better understand its integrated economic value [322] within the framework of the biorefinery concept and the bio-based economy for C. sativa [270,323]. More detailed examination of phenotypes is also necessary to discover natural variation in quality traits, such as the low erucic acid content of its oil [161], and production traits, such as improved heat and drought tolerance [324]. Lastly, increasing market demands for bio-based products for the renewable fuels market and improvements in value-chain cost efficiencies will likely increase future demands for this versatile crop, with great potential for agricultural production systems of the future that will increasingly rely on more climate-resilient crops such as C. sativa.
Author Contributions
Conceptualization, D.N. and J.C.C.; methodology, D.N. and R.H.L.; validation, D.N. and R.H.L.; formal analysis, D.N. and R.H.L.; investigation, D.N. and R.H.L.; resources, J.K.Q.S. and J.C.C.; data curation, D.N. and R.H.L.; writing—original draft preparation, D.N.; writing—review and editing, D.N. and J.C.C.; visualization, D.N. and J.C.C.; supervision, J.K.Q.S. and J.C.C. project administration, J.K.Q.S. and J.C.C.; funding acquisition, J.K.Q.S. and J.C.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Hatch grant funding from the Nevada Agricultural Experiment Station under projects NEV-00372 and NEV-00384.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Lisa Petrusa for critical reading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Bansal, S.; Durrett, T.P. Camelina sativa: An ideal platform for the metabolic engineering and field production of industrial lipids. Biochimie 2016, 120, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berti, M.; Gesch, R.; Eynck, C.; Anderson, J.; Cermak, S. Camelina uses, genetics, genomics, production, and management. Ind. Crops Prod. 2016, 94, 690–710. [Google Scholar] [CrossRef]
- Gesch, R.W.; Matthees, H.L.; Alvarez, A.L.; Gardner, R.D. Winter Camelina: Crop growth, seed yield, and quality response to cultivar and seeding rate. Crop Sci. 2018, 58, 2089–2098. [Google Scholar] [CrossRef]
- Putnam, D.H.; Budin, J.T.; Field, L.A.; Breene, W.M. Camelina: A promising low-input oilseed. In New Crops; Janick, J., Simon, J.E., Eds.; Wiley: New York, NY, USA, 1993; pp. 314–322. [Google Scholar]
- Schillinger, W.F. Camelina: Long-term cropping systems research in a dry Mediterranean climate. Field Crops Res. 2019, 235, 87–94. [Google Scholar] [CrossRef]
- Vollmann, J.; Eynck, C. Camelina as a sustainable oilseed crop: Contributions of plant breeding and genetic engineering. Biotechnol. J. 2015, 10, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Masella, P.; Martinelli, T.; Galasso, I. Agronomic evaluation and phenotypic plasticity of Camelina sativa growing in Lombardia, Italy. Crop Pasture Sci. 2014, 65, 453–460. [Google Scholar] [CrossRef]
- Iskandarov, U.; Kim, H.J.; Cahoon, E.B. Camelina: An emerging oilseed platform for advanced biofuels and bio-based materials. In Plants and Bioenergy; Advances in Plant Biology; McCann, M., Buckeridge, M., Carpita, N., Eds.; Springer: New York, NY, USA, 2014; Volume 4, pp. 131–140. [Google Scholar]
- Liu, X.; Brost, J.; Hutcheon, C.; Guilfoil, R.; Wilson, A.K.; Leung, S.; Shewmaker, C.K.; Rooke, S.; Nguyen, T.; Kiser, J. Transformation of the oilseed crop Camelina sativa by Agrobacterium-mediated floral dip and simple large-scale screening of transformants. Vitr. Cell. Dev. Biol.-Plant 2012, 48, 462–468. [Google Scholar] [CrossRef]
- Lu, C.; Kang, J. Generation of transgenic plants of a potential oilseed crop Camelina sativa by Agrobacterium-mediated transformation. Plant Cell Rep. 2008, 27, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Sainger, M.; Jaiwal, A.; Sainger, P.A.; Chaudhary, D.; Jaiwal, R.; Jaiwal, P.K. Advances in genetic improvement of Camelina sativa for biofuel and industrial bio-products. Renew. Sustain. Energy Rev. 2017, 68, 623–637. [Google Scholar] [CrossRef]
- Sitther, V.; Tabatabai, B.; Enitan, O.; Dhekney, S. Agrobacterium-mediated transformation of Camelina sativa for production of transgenic plants. J. Biol. Methods 2018, e83. [Google Scholar] [CrossRef] [Green Version]
- Yemets, A.I.; Boychuk, Y.N.; Shysha, E.N.; Rakhmetov, D.B.; Blume, Y.B. Establishment of in vitro culture, plant regeneration, and genetic transformation of Camelina sativa. Cytol. Genet. 2013, 47, 138–144. [Google Scholar] [CrossRef] [Green Version]
- Ozseyhan, M.E.; Kang, J.; Mu, X.; Lu, C. Mutagenesis of the FAE1 genes significantly changes fatty acid composition in seeds of Camelina sativa. Plant Physiol. Biochem. 2018, 123, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Morineau, C.; Bellec, Y.; Tellier, F.; Gissot, L.; Kelemen, Z.; Nogué, F.; Faure, J.D. Selective gene dosage by CRISPR-Cas9 genome editing in hexaploid Camelina sativa. Plant Biotechnol. J. 2017, 15, 729–739. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.Z.; Henry, I.M.; Lynagh, P.G.; Comai, L.; Cahoon, E.B.; Weeks, D.P. Significant enhancement of fatty acid composition in seeds of the allohexaploid, Camelina sativa, using CRISPR/Cas9 gene editing. Plant Biotechnol. J. 2017, 15, 648–657. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, S.; Bhattacharya, A.; Khare, S.K.; Kaushik, G. Camelina sativa: An Emerging Biofuel Crop; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–38. [Google Scholar]
- Dharavath, R.N.; Singh, S.; Chaturvedi, S.; Luqman, S. Camelina sativa (L.) Crantz A mercantile crop with speckled pharmacological activities. Ann. Phytomed. 2016, 5, 6–26. [Google Scholar] [CrossRef]
- Malik, M.R.; Tang, J.; Sharma, N.; Burkitt, C.; Ji, Y.; Mykytyshyn, M.; Bohmert-Tatarev, K.; Peoples, O.; Snell, K.D. Camelina sativa, an oilseed at the nexus between model system and commercial crop. Plant Cell Rep. 2018, 37, 1367–1381. [Google Scholar] [CrossRef]
- Moser, B.R. Camelina (Camelina sativa L.) oil as a biofuels feedstock: Golden opportunity or false hope? Lipid Technol. 2010, 22, 270–273. [Google Scholar] [CrossRef]
- Murphy, E.J. Camelina (Camelina sativa). In Industrial Oil Crops; McKeon, T., Hayes, D., Hildebrand, D., Weselake, R., Eds.; Elevier: London, UK, 2016; pp. 207–230. [Google Scholar]
- Obour, K.A.; Sintim, H.Y.; O’bend, E.; Jeliazkov, V.D. Oilseed Camelina (Camelina sativa L. Crantz): Production systems, prospects and challenges in the USA Great Plains. Adv. Plants Agric. Res. 2015, 2, 00043. [Google Scholar] [CrossRef] [Green Version]
- Waraich, E.A.; Ahmed, Z.; Ahmad, R.; Ashraf, M.Y.; Naeem, M.S.; Rengel, Z. ‘Camelina sativa’, a climate proof crop, has high nutritive value and multiple-uses: A review. Aust. J. Crop Sci. 2013, 7, 1551–1559. [Google Scholar]
- Zubr, J. Oil-seed crop: Camelina sativa. Ind. Crops Prod. 1997, 6, 113–119. [Google Scholar] [CrossRef]
- Fröhlich, A.; Rice, B. Evaluation of Camelina sativa oil as a feedstock for biodiesel production. Ind. Crops Prod. 2005, 21, 25–31. [Google Scholar] [CrossRef]
- Ciubota-Rosie, C.; Ruiz, J.R.; Ramos, M.J.; Pérez, Á. Biodiesel from Camelina sativa: A comprehensive characterisation. Fuel 2013, 105, 572–577. [Google Scholar] [CrossRef]
- Zaleckas, E.; Makarevičienė, V.; Sendžikienė, E. Possibilities of using Camelina sativa oil for producing biodiesel fuel. Transport 2012, 27, 60–66. [Google Scholar] [CrossRef]
- Agarwal, A.; Prakash, O.; Bala, M. Camelina sativa, a short gestation oilseed crop with biofuel potential: Opportunities for Indian scenario. Oil Crop Sci. 2021, 6, 114–121. [Google Scholar] [CrossRef]
- Domestication of Plants in the Old World, 4th ed.; Oxford University Press: Oxford, UK, 2012.
- Larsson, M. Cultivation and processing of Linum usitatissimum and Camelina sativa in southern Scandinavia during the Roman Iron Age. Veg. Hist. Archaeobotany 2013, 22, 509–520. [Google Scholar] [CrossRef]
- Dönmez, E.O.; Belli, O. Urartian plant cultivation at Yoncatepe (Van), eastern Turkey. Econ. Bot. 2007, 61, 290–298. [Google Scholar] [CrossRef]
- Carciumaru, M. Cultivarea plantelor in Dacia (Plants cultivation in Dacia). Thraco-Dacica 2007, 8, 1–6. [Google Scholar]
- Knorzer, K.H. Evolution and spread of Gold of Pleasure (Camelina sativa S.L.). Ber. Dtsch. Bot. Ges. 1978, 91, 187–195. [Google Scholar]
- Hutcheon, C.; Ditt, R.F.; Beilstein, M.; Comai, L.; Schroeder, J.; Goldstein, E.; Shewmaker, C.K.; Nguyen, T.; De Rocher, J.; Kiser, J. Polyploid genome of Camelina sativa revealed by isolation of fatty acid synthesis genes. BMC Plant Biol. 2010, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Gesch, R.W. Influence of genotype and sowing date on camelina growth and yield in the north central U.S. Ind. Crops Prod. 2014, 54, 209–215. [Google Scholar] [CrossRef]
- Neupane, D.; Solomon, J.K.Q.; McLennon, E.; Davison, J.; Lawry, T. Camelina production parameters response to different irrigation regimes. Ind. Crops Prod. 2020, 148, 112286. [Google Scholar] [CrossRef]
- Lohaus, R.H.; Neupane, D.; Mengistu, M.A.; Solomon, J.K.Q.; Cushman, J.C. Five-year field trial of eight Camelina sativa cultivars for biomass to be used in biofuel under irrigated conditions in a semi-arid climate. Agronomy 2020, 10, 562. [Google Scholar] [CrossRef]
- Blackshaw, R.; Johnson, E.; Gan, Y.; May, W.; McAndrew, D.; Barthet, V.; McDonald, T.; Wispinski, D. Alternative oilseed crops for biodiesel feedstock on the Canadian prairies. Can. J. Plant Sci. 2011, 91, 889–896. [Google Scholar] [CrossRef]
- Gugel, R.K.; Falk, K.C. Agronomic and seed quality evaluation of Camelina sativa in western Canada. Can. J. Plant Sci. 2006, 86, 1047–1058. [Google Scholar] [CrossRef]
- Urbaniak, S.D.; Caldwell, C.D.; Zheljazkov, V.D.; Lada, R.; Luan, L. The effect of seeding rate, seeding date and seeder type on the performance of Camelina sativa L. in the Maritime Provinces of Canada. Can. J. Plant Sci. 2008, 88, 501–508. [Google Scholar] [CrossRef]
- Beilstein, M.A.; Al-Shehbaz, I.A.; Kellogg, E.A. Brassicaceae phylogeny and trichome evolution. Am. J. Bot. 2006, 93, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Beilstein, M.A.; Al-Shehbaz, I.A.; Mathews, S.; Kellogg, E.A. Brassicaceae phylogeny inferred from phytochrome A and ndhF sequence data: Tribes and trichomes revisited. Am. J. Bot. 2008, 95, 1307–1327. [Google Scholar] [CrossRef] [Green Version]
- Brock, J.R.; Donmez, A.A.; Beilstein, M.A.; Olsen, K.M. Phylogenetics of Camelina Crantz. (Brassicaceae) and insights on the origin of gold-of-pleasure (Camelina sativa). Mol. Phylogenetics Evol. 2018, 127, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Brock, J.R.; Mandakova, T.; Lysak, M.A.; Al-Shehbaz, I.A. Camelina neglecta (Brassicaceae, Camelineae), a new diploid species from Europe. PhytoKeys 2019, 115, 51–57. [Google Scholar] [CrossRef]
- Ghamkhar, K.; Croser, J.; Aryamanesh, N.; Campbell, M.; Konkova, N.; Francis, C. Camelina (Camelina sativa (L). Crantz) as an alternative oilseed: Molecular and ecogeographic analyses. Genome 2010, 53, 558–567. [Google Scholar] [CrossRef]
- Kagale, S.; Koh, C.; Nixon, J.; Bollina, V.; Clarke, W.E.; Tuteja, R.; Spillane, C.; Robinson, S.J.; Links, M.G.; Clarke, C.; et al. The emerging biofuel crop Camelina sativa retains a highly undifferentiated hexaploid genome structure. Nat. Commun. 2014, 5, 3706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galasso, I.A.; Braglia, M.L.; Ponzoni, E.; Breviario, D. Genomic fingerprinting of Camelina species using cTBP as molecular marker. Am. J. Plant Sci. 2015, 6, 1184–1200. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.L.; Lujan-Toro, B.E.; Sauder, C.A.; James, T.; Ohadi, S.; Hall, L.M. An update to the Canadian range, abundance, and ploidy of Camelina spp. (Brassicaceae) east of the Rocky Mountains. Botany 2017, 95, 405–417. [Google Scholar] [CrossRef]
- Francis, A.; Warwick, S.I. The biology of Canadian weeds. 142. Camelina alyssum (Mill.) thell. C. microcarpa andrz. ex DC.; C. sativa (L.) crantz. Can. J. Plant Sci. 2009, 89, 791–810. [Google Scholar] [CrossRef]
- Gehringer, A.; Friedt, W.; Luhs, W.; Snowdon, R.J. Genetic mapping of agronomic traits in false flax (Camelina sativa subsp. sativa). Genome 2006, 49, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Al-Shehbaz, I.A.; Beilstein, M.A. Flora of North America North of Mexico. N. Y. Oxf. 2010, 7, 451–453. [Google Scholar]
- Koch, M.A.; Kiefer, M.; German, D.A.; Al-Shehbaz, I.A.; Franzke, A.; Mummenhoff, K.; Schmickl, R. BrassiBase: Tools and biological resources to study characters and traits in the Brassicaceae version 1.1. Taxon 2012, 61, 1001–1009. [Google Scholar] [CrossRef]
- Chaudhary, R.; Koh, C.S.; Kagale, S.; Tang, L.; Wu, S.W.; Lv, Z.; Mason, A.S.; Sharpe, A.G.; Diederichsen, A.; Parkin, I.A.P. Assessing diversity in the Camelina genus provides insights into the genome structure of Camelina sativa. G3 Genes Genomes Genet. 2020, 10, 1297–1308. [Google Scholar] [CrossRef] [Green Version]
- Brock, J.R.; Mandáková, T.; McKain, M.; Lysak, M.A.; Olsen, K.M. Chloroplast phylogenomics in Camelina (Brassicaceae) reveals multiple origins of polyploid species and the maternal lineage of C. sativa. Hortic. Res. 2022, 9, uhab050. [Google Scholar] [CrossRef] [PubMed]
- Mirek, Z. Genus Camelina in Poland—Taxonomy, Distribution and Habitats. Fragm. Florist. Geobot. 1981, 27, 445–503. [Google Scholar]
- Gesch, R.W.; Archer, D.W. Double-cropping with winter camelina in the northern corn belt to produce fuel and food. Ind. Crops Prod. 2013, 44, 718–725. [Google Scholar] [CrossRef]
- Gesch, R.W.; Cermak, S. Sowing date and tillage effects on fall-seeded camelina in the northern Corn Belt. Agron. J. 2011, 103, 980–987. [Google Scholar] [CrossRef] [Green Version]
- Johnson, G.A.; Wells, M.S.; Anderson, K.; Gesch, R.W.; Forcella, F.; Wyse, D.L. Yield tradeoffs and nitrogen between pennycress, camelina, and soybean in relay- and double-crop systems. Agron. J. 2017, 109, 2128–2135. [Google Scholar] [CrossRef]
- Neupane, D.; Solomon, J.K.Q.; Davison, J.; Lawry, T. Nitrogen source and rate effects on grain and potential biodiesel production of camelina in the semiarid environment of northern Nevada. Glob. Change Biol. Bioenergy 2018, 10, 861–876. [Google Scholar] [CrossRef] [Green Version]
- Walia, M.K.; Wells, M.S.; Cubins, J.; Wyse, D.; Gardner, R.D.; Forcella, F.; Gesch, R. Winter camelina seed yield and quality responses to harvest time. Ind. Crops Prod. 2018, 124, 765–775. [Google Scholar] [CrossRef]
- Berti, M.; Wilckens, R.; Fischer, S.; Solis, A.; Johnson, B. Seeding date influence on camelina seed yield, yield components, and oil content in Chile. Ind. Crops Prod. 2011, 34, 1358–1365. [Google Scholar] [CrossRef]
- Neupane, D.; Solomon, J.K.Q.; McLennon, E.; Davison, J.; Lawry, T. Sowing date and sowing method influence on camelina cultivars grain yield, oil concentration, and biodiesel production. Food Energy Secur. 2019, 8, e00166. [Google Scholar] [CrossRef] [Green Version]
- Angelini, L.G.; Moscheni, E.; Colonna, G.; Belloni, P.; Bonari, E. Variation in agronomic characteristics and seed oil composition of new oilseed crops in central Italy. Ind. Crops Prod. 1997, 6, 313–323. [Google Scholar] [CrossRef]
- Vollmann, J.; Moritz, T.; Kargl, C.; Baumgartner, S.; Wagentristl, H. Agronomic evaluation of camelina genotypes selected for seed quality characteristics. Ind. Crops Prod. 2007, 26, 270–277. [Google Scholar] [CrossRef]
- Dai, A.; Zhao, T.; Chen, J. Climate Change and Drought: A Precipitation and Evaporation Perspective. Curr. Clim. Change Rep. 2018, 4, 301–312. [Google Scholar] [CrossRef]
- Naumann, G.; Alfieri, L.; Wyser, K.; Mentaschi, L.; Betts, R.A.; Carrao, H.; Spinoni, J.; Vogt, J.; Feyen, L. Global changes in drought conditions under different levels of warming. Geophys. Res. Lett. 2018, 45, 3285–3296. [Google Scholar] [CrossRef]
- McKenzie, B.A.; Smallfield, B.M.; Fasi, V.; Martin, R.J. Possible species for the production of biodiesel on marginal land. Agron. N. Z. 2011, 41, 97–108. [Google Scholar]
- Pavlista, A.D.; Isbell, T.A.; Baltensperger, D.D.; Hergert, G.W. Planting date and development of spring-seeded irrigated canola, brown mustard and camelina. Ind. Crops Prod. 2011, 33, 451–456. [Google Scholar] [CrossRef] [Green Version]
- Hergert, G.W.; Margheim, J.F.; Pavlista, A.D.; Martin, D.L.; Isbell, T.A.; Supalla, R.J. Irrigation response and water productivity of deficit to fully irrigated spring camelina. Agric. Water Manag. 2016, 177, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Hergert, G.W.; Margheim, J.F.; Pavlista, A.D.; Martin, D.L.; Supalla, R.J.; Isbell, T.A. Yield, irrigation response, and water productivity of deficit to fully irrigated spring canola. Agric. Water Manag. 2016, 168, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Matteo, R.; D’Avino, L.; Ramirez-Cando, L.J.; Pagnotta, E.; Angelini, L.G.; Spugnoli, P.; Tavarini, S.; Ugolini, L.; Foschi, L.; Lazzeri, L. Camelina (Camelina sativa L. Crantz) under low-input management systems in northern Italy: Yields, chemical characterization and environmental sustainability. Ital. J. Agron. 2020, 15, 132–143. [Google Scholar] [CrossRef]
- Załuski, D.; Tworkowski, J.; Krzyżaniak, M.; Stolarski, M.J.; Kwiatkowski, J. The characterization of 10 spring Camelina genotypes grown in environmental conditions in North-eastern Poland. Agronomy 2020, 10, 64. [Google Scholar] [CrossRef] [Green Version]
- Lošák, T.; Hlusek, J.; Martinec, J.; Vollmann, J.; Peterka, J.; Filipcik, R.; Varga, L.; Ducsay, L.; Martensson, A. Effect of combined nitrogen and sulphur fertilization on yield and qualitative parameters of Camelina sativa [L.] Crtz. (false flax). Acta Agric. Scand. Sect. B-Soil Plant Sci. 2011, 61, 313–321. [Google Scholar] [CrossRef]
- Manore, D.; Yohanns, A. Evaluating growth, seed yield and yield attributes of Camelina (Camelina sativa L) in response to seeding rate and nitrogen fertilizer levels under irrigation condition, southern Ethiopia. Agric. For. Fish. 2019, 8, 31–35. [Google Scholar] [CrossRef]
- Obeng, E.; Obour, A.K.; Nelson, N.O.; Moreno, J.A.; Ciampitti, I.A.; Wang, D.; Durrett, T.P. Seed yield and oil quality as affected by Camelina cultivar and planting date. J. Crop Improv. 2019, 33, 202–222. [Google Scholar] [CrossRef]
- Amiri-Darban, N.; Nourmohammadi, G.; Rad, A.H.S.; Mirhadi, S.M.J.; Heravan, I.M. Potassium sulfate and ammonium sulfate affect quality and quantity of camelina oil grown with different irrigation regimes. Ind. Crops Prod. 2020, 148, 112308. [Google Scholar] [CrossRef]
- Zanetti, F.; Eynck, C.; Christou, M.; Krzyżaniak, M.; Righini, D.; Alexopoulou, E.; Stolarski, M.J.; Van Loo, E.N.; Puttick, D.; Monti, A. Agronomic performance and seed quality attributes of Camelina (Camelina sativa L. crantz) in multi-environment trials across Europe and Canada. Ind. Crops Prod. 2017, 107, 602–608. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, Y.A.; Chen, C.; Lamb, P.; Afshar, R.K. Agronomic Evaluation of Camelina (Camelina sativa L. Crantz) Cultivars for Biodiesel Feedstock. BioEnergy Res. 2017, 10, 792–799. [Google Scholar] [CrossRef]
- Obour, A.K.; Obeng, E.; Mohammed, Y.A.; Ciampitti, I.A.; Durrett, T.P.; Aznar-Moreno, J.A.; Chen, C.C. Camelina seed yield and fatty acids as influenced by genotype and wnvironment. Agron. J. 2017, 109, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Obour, A.K.; Chen, C.C.; Sintim, H.Y.; McVay, K.; Lamb, P.; Obeng, E.; Mohammed, Y.A.; Khan, Q.; Afshar, R.K.; Zheljazkov, V.D. Camelina sativa as a fallow replacement crop in wheat-based crop production systems in the US Great Plains. Ind. Crops Prod. 2018, 111, 22–29. [Google Scholar] [CrossRef]
- Pavlista, A.D.; Hergert, G.W.; Margheim, J.M.; Isbell, T.A. Growth of spring camelina (Camelina sativa) under deficit irrigation in western Nebraska. Ind. Crops Prod. 2016, 83, 118–123. [Google Scholar] [CrossRef] [Green Version]
- French, A.N.; Hunsaker, D.; Thorp, K.; Clarke, T. Evapotranspiration over a camelina crop at Maricopa, Arizona. Ind. Crops Prod. 2009, 29, 289–300. [Google Scholar] [CrossRef]
- Hunsaker, D.J.; French, A.N.; Clarke, T.R.; El-Shikha, D.M. Water use, crop coefficients, and irrigation management criteria for camelina production in arid regions. Irrig. Sci. 2011, 29, 27–43. [Google Scholar] [CrossRef]
- Hunsaker, D.J.; French, A.N.; Thorp, K.R. Camelina water use and seed yield response to irrigation scheduling in an arid environment. Irrig. Sci. 2012, 31, 911–929. [Google Scholar] [CrossRef] [Green Version]
- George, N.; Thompson, S.E.; Hollingsworth, J.; Orloff, S.; Kaffka, S. Measurement and simulation of water-use by canola and camelina under cool-season conditions in California. Agric. Water Manag. 2018, 196, 15–23. [Google Scholar] [CrossRef]
- Bronson, K.F.; Hunsaker, D.J.; Thorp, K.R. Nitrogen fertilizer and irrigation effects on seed yield and oil in Camelina. Agron. J. 2019, 111, 1712–1719. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Caldwell, C.D.; Falk, K.C.; Lada, R.R.; MacDonald, D. Camelina yield and quality response to combined nitrogen and sulfur. Agron. J. 2013, 105, 1847–1852. [Google Scholar] [CrossRef]
- Malhi, S.S.; Johnson, E.N.; Hall, L.M.; May, W.E.; Phelps, S.; Nybo, B. Effect of nitrogen fertilizer application on seed yield, N uptake, and seed quality of Camelina sativa. Can. J. Soil Sci. 2014, 94, 35–47. [Google Scholar] [CrossRef]
- Martinez, S.; Alvarez, S.; Capuano, A.; del Mar Delgado, M. Environmental performance of animal feed production from Camelina sativa (L.) Crantz: Influence of crop management practices under Mediterranean conditions. Agric. Syst. 2020, 177, 102717. [Google Scholar] [CrossRef]
- Krzyżaniak, M.; Stolarski, M.J. Life cycle assessment of camelina and crambe production for biorefinery and energy purposes. J. Clean. Prod. 2019, 237, 117755. [Google Scholar] [CrossRef]
- Sintim, H.Y.; Zheljazkov, V.D.; Obour, A.K.; Garcia y Garcia, A.; Foulke, T.K. Influence of nitrogen and sulfur application on camelina performance under dryland conditions. Ind. Crops Prod. 2015, 70, 253–259. [Google Scholar] [CrossRef]
- Kemp, W.H. Biodiesel: Basics and Beyond, a Comprehensive Guide to Production and Use for the Home and Farm; Aztext Press: Tamworth, ON, Canada, 2006. [Google Scholar]
- Bobrecka-Jamro, M.C. The effects of varied plant density and nitrogen fertilization on quantity and quality yield of Camelina sativa L. Emir. J. Food Agric. 2018, 29, 988–993. [Google Scholar] [CrossRef]
- Keshavarz-Afshar, R.; Mohammed, Y.A.; Chen, C. Energy balance and greenhouse gas emissions of dryland camelina as influenced by tillage and nitrogen. Energy 2015, 91, 1057–1063. [Google Scholar] [CrossRef] [Green Version]
- Rathke, G.-W.; Christen, O.; Diepenbrock, W. Effects of N source and rate on productivity and quality of winter oilseed rape (Brassica napus L.) grown in different crop rotations. Field Crops Res. 2005, 94, 103–113. [Google Scholar] [CrossRef]
- Leclère, M.; Jeuffroy, M.-H.; Butier, A.; Chatain, C.; Loyce, C. Controlling weeds in camelina with innovative herbicide-free crop management routes across various environments. Ind. Crops Prod. 2019, 140, 111605. [Google Scholar] [CrossRef]
- Lovett, J.V.; Jackson, H.F. Allelopathic activity of Camelina sativa (L.) Crantz inrelation to its phyllosphere bacteria. New Phytol. 1980, 86, 273–277. [Google Scholar] [CrossRef]
- Walsh, K.D.; Sanderson, D.; Hall, L.M.; Mugo, S.; Hills, M.J. Allelopathic effects of camelina (Camelina sativa) and canola (Brassica napus) on wild oat, flax and radish. Allelopath. J. 2014, 33, 83–96. [Google Scholar]
- Abbas, A.; Huang, P.; Hussain, S.; Saqib, M.; He, L.; Shen, F.; Du, D. Application of allelopathic phenomemna to enhance growth and production of camelina (Camelina sativa (L.). Appl. Ecol. Environ. Res. 2021, 19, 453–469. [Google Scholar] [CrossRef]
- Davis, P.B.; Maxwell, B.; Menalled, F.D. Impact of growing conditions on the competitive ability of Camelina sativa (L.) Crantz (Camelina). Can. J. Plant Sci. 2013, 93, 243–247. [Google Scholar] [CrossRef]
- Lenssen, A.W.; Iversen, W.M.; Sainju, U.M.; Caesar-TonThat, T.C.; Blodgett, S.L.; Allen, B.L.; Evans, R.G. Yield, pests, and water use of durum and selected crucifer oilseeds in two-year rotations. Agron. J. 2012, 104, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- Julié-Galau, S.; Bellec, Y.; Faure, J.-D.; Tepfer, M. Evaluation of the potential for interspecific hybridization between Camelina sativa and related wild Brassicaceae in anticipation of field trials of GM camelina. Transgenic Res. 2014, 23, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-J.; Auer, C. Hybridization between Camelina sativa (L.) Crantz and common Brassica weeds. Ind. Crops Prod. 2020, 147, 112240. [Google Scholar] [CrossRef]
- Davis, P.B. The Invasion Potential and Competitive Ability of Camelina sativa (L.) Crantz (Camelina) in Rangeland Ecosystems; Montana State University-Bozeman, College of Agriculture: Bozeman, MT, USA, 2010. [Google Scholar]
- Enjalbert, J.; Johnson, J. Guide for Producing Dryland Camelina in Eastern Colorado; Colorado State University Extension: Fort Collins, CO, USA, 2011. [Google Scholar]
- WAPMC. Biomass Crop Assistance Program: Camelina Agronomy Sheet; NRCS: Washington, DC, USA, 2012. [Google Scholar]
- Sobiech, Ł.; Grzanka, M.; Kurasiak-Popowska, D.; Radzikowska, D. Phytotoxic effect of herbicides on various camelina [Camelina sativa (L.) Crantz] genotypes and plant chlorophyll fluorescence. Agriculture 2020, 10, 185. [Google Scholar] [CrossRef]
- Scheliga, M.; Petersen, J. Selectivity of herbicides in camelina (Camelina sativa (L.) Crtz.). Jul.-Kühn-Arch. 2016, 452, 154–162. [Google Scholar] [CrossRef]
- Jha, P.; Stougaard, R.N. Camelina (Camelina sativa) tolerance to selected preemergence herbicides. Weed Technol. 2013, 27, 712–717. [Google Scholar] [CrossRef]
- Walsh, D.T.; Babiker, E.M.; Burke, I.C.; Hulbert, S.H. Camelina mutants resistant to acetolactate synthase inhibitor herbicides. Mol. Breed. 2012, 30, 1053–1063. [Google Scholar] [CrossRef]
- Schillinger, W.F.; Wysocki, D.J.; Chastain, T.G.; Guy, S.O.; Karow, R.S. Camelina: Planting date and method effects on stand establishment and seed yield. Field Crops Res. 2012, 130, 138–144. [Google Scholar] [CrossRef] [Green Version]
- Saucke, H.; Ackermann, K. Weed suppression in mixed cropped grain peas and false flax (Camelina sativa). Weed Res. 2006, 46, 453–461. [Google Scholar] [CrossRef]
- Almoussawi, A.; Lenoir, J.; Spicher, F.; Dupont, F.; Chabrerie, O.; Closset-Kopp, D.; Brasseur, B.; Kobaissi, A.; Dubois, F.; Decocq, G. Direct seeding associated with a mixture of winter cover crops decreases weed abundance while increasing cash-crop yields. Soil Tillage Res. 2020, 200, 104622. [Google Scholar] [CrossRef]
- Royo-Esnal, A.; Valencia-Gredilla, F. Camelina as a rotation crop for weed control in organic farming in a semiarid Mediterranean climate. Agriculture 2018, 8, 156. [Google Scholar] [CrossRef] [Green Version]
- Berti, M.; Gesch, R.; Johnson, B.; Ji, Y.; Seames, W.; Aponte, A. Double-and relay-cropping of energy crops in the northern Great Plains, USA. Ind. Crops Prod. 2015, 75, 26–34. [Google Scholar] [CrossRef]
- Berti, M.; Samarappuli, D.; Johnson, B.L.; Gesch, R.W. Integrating winter camelina into maize and soybean cropping systems. Ind. Crops Prod. 2017, 107, 595–601. [Google Scholar] [CrossRef]
- Chandra, K.; Sharma, D.K.; Meher, L.C.; Kulkarni, A.V.; Nasim, M. Studies of feasibility of intercropping of Camelina sativa in jatropha plantation in semi–arid climate in Andhra Pradesh, India. Res. J. Agric. For. Sci. 2014, 2, 23–26. [Google Scholar]
- Robinson, R.G. Camelina: A Useful Research Crop and a Potential Oilseed Crop. 1987, 579. Available online: https://conservancy.umn.edu/bitstream/handle/11299/141546/SB579.pdf?sequence=1 (accessed on 10 February 2022).
- Soroka, J.J.; Olivier, C.; Wist, T.J.; Grenkow, L. Present and potential impacts of insects on Camelina and Crambe. In Integrated Management of Insect Pests on Canola and Other Brassica Oilseed Crops; CABI: Boston, MA, USA, 2017; pp. 316–340. [Google Scholar]
- Nielsen, J.K.; Hansen, M.L.; Agerbirk, N.; Petersen, B.L.; Halkier, B.A. Responses of the flea beetles Phyllotreta nemorum and P. cruciferae to metabolically engineered Arabidopsis thaliana with an altered glucosinolate profile. Chemoecology 2001, 11, 75–83. [Google Scholar] [CrossRef]
- Pachagounder, P.; Lamb, R.J.; Bodnaryk, R.P. Resistance to the flea beetle Phyllotreta cruciferae (Coleoptera: Chrysomelidae) in false flax, Camelina sativa (Brassicaceae). Can. Entomol. 1998, 130, 235–240. [Google Scholar] [CrossRef]
- Henderson, A.E.; Hallett, R.H.; Soroka, J.J. Prefeeding behavior of the crucifer flea beetle, Phyllotreta cruciferae, on host and nonhost crucifers. J. Insect Behav. 2004, 17, 17–39. [Google Scholar] [CrossRef]
- Onyilagha, J.C.; Gruber, M.Y.; Hallett, R.H.; Holowachuk, J.; Buckner, A.; Soroka, J.J. Constitutive flavonoids deter flea beetle insect feeding in Camelina sativa L. Biochem. Syst. Ecol. 2012, 42, 128–133. [Google Scholar] [CrossRef]
- Soroka, J.; Olivier, C.; Grenkow, L.; Séguin-Swartz, G. Interactions between Camelina sativa (Brassicaceae) and insect pests of canola. Can. Entomol. 2015, 147, 193–214. [Google Scholar] [CrossRef]
- Cárcamo, H.; Olfert, O.; Dosdall, L.; Herle, C.; Beres, B.; Soroka, J. Resistance to cabbage seedpod weevil among selected Brassicaceae germplasm. Can. Entomol. 2007, 139, 658–669. [Google Scholar] [CrossRef]
- Jewett, F.G. Camelina sativa: For biofuels and bioproducts. Ind. Crops 2015, 9, 157–170. [Google Scholar] [CrossRef]
- Chesnais, Q.; Verzeaux, J.; Couty, A.; Le Roux, V.; Ameline, A. Is the oilseedcrop Camelina sativa a potential host for aphid pests? BioEnergy Res. 2015, 8, 91–99. [Google Scholar] [CrossRef]
- Séguin-Swartz, G.; Eynck, C.; Gugel, R.K.; Strelkov, S.E.; Olivier, C.Y.; Li, J.L.; Klein-Gebbinck, H.; Borhan, H.; Caldwell, C.D.; Falk, K.C. Diseases of Camelina sativa (false flax). Can. J. Plant Pathol. 2009, 31, 375–386. [Google Scholar] [CrossRef]
- Conn, K.L.; Tewari, J.P.; Dahiya, J.S. Resistance to Alternaria brassicae and phytoalexin-elicitation in rapeseed and other crucifers. Plant Sci. 1988, 56, 21–25. [Google Scholar] [CrossRef]
- Sharma, G.; Kumar, V.D.; Haque, A.; Bhat, S.R.; Prakash, S.; Chopra, V.L. Brassica coenospecies: A rich reservoir for genetic resistance to leaf spot caused by Alternaria brassicae. Euphytica 2002, 125, 411–417. [Google Scholar] [CrossRef]
- Jejelowo, O.A.; Conn, K.L.; Tewari, J.P. Relationship between conidial concentration, germling growth, and phytoalexin production by Camelina sativa leaves inoculated with Alternaria brassicae. Mycol. Res. 1991, 95, 928–934. [Google Scholar] [CrossRef]
- Dixit, S.; Jangid, V.K.; Grover. Evaluation of physiological and molecular effect of variable virulence of Alternaria brassicae isolates in Brassica juncea, Sinapis alba and Camelina sativa. Plant Physiol. Biochem. 2020, 155, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Westman, A.L.; Kresovich, S.; Dickson, M.H. Regional variation in Brassica nigra and other weedy crucifers for disease reaction to Alternaria brassicicola and Xanthomonas campestris pv. campestris. Euphytica 1999, 106, 253–259. [Google Scholar] [CrossRef]
- Sigareva, M.A.; Earle, E.D. Camalexin induction in intertribal somatic hybrids between Camelina sativa and rapid-cycling Brassica oleracea. Theor. Appl. Genet. 1999, 98, 164–170. [Google Scholar] [CrossRef]
- Hansen, L.N. Intertribal somatic hybridization between rapid cycling Brassica oleracea L. and Camelina sativa (L.) Crantz. Euphytica 1998, 104, 173–179. [Google Scholar] [CrossRef]
- Browne, L.M.; Conn, K.L.; Ayer, W.A.; Tewari, J.P. The camalexins: New phytoalexins produced in the leaves of Camelina sativa (Cruciferae). Tetrahedron 1991, 47, 3909–3914. [Google Scholar] [CrossRef]
- Mir, Z.A.; Ali, S.; Shivaraj, S.M.; Bhat, J.A.; Singh, A.; Yadav, P.; Rawat, S.; Paplao, P.K.; Grover, A. Genome-wide identification and characterization of chitinase gene family in Brassica juncea and Camelina sativa in response to Alternaria brassicae. Genomics 2020, 112, 749–763. [Google Scholar] [CrossRef]
- Li, H.; Barbetti, M.J.; Sivasithamparam, K. Hazard from reliance on cruciferous hosts as sources of major gene-based resistance for managing blackleg (Leptosphaeria maculans) disease. Field Crops Res. 2005, 91, 185–198. [Google Scholar] [CrossRef]
- Paul, V.H.; Tewari, J.P.; Tewari, I.; Foller, I.; Henneken, M. Joint field experiment on cultivation, yield, and diseases of false flax (Camelina sativa) in Germany and Canada. IOBC WPRS BULLETIN 2000, 23, 205–208. [Google Scholar]
- Föller, I.; Henneken, M.; Paul, V.H. Occurrence of false flax diseases (Camelina sativa (L.) CRTZ.) in field trials in Germany in 1995 and 1996. Bull. OILB/SROP 1998, 21, 65–76. [Google Scholar]
- Putnam, M.L.; Serdani, M.; Ehrensing, D.; Curtis, M. Camelina infected by downy mildew (Hyaloperonospora camelinae) in the western United States: A first report. Plant Health Prog. 2009, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Purnamasari, M.; Cawthray, G.R.; Barbetti, M.J.; Erskine, W.; Croser, J.S. Camalexin production in Camelina sativa is independent of cotyledon resistance to Sclerotinia sclerotiorum. Plant Dis. 2015, 99, 1544–1549. [Google Scholar] [CrossRef] [Green Version]
- Eynck, C.; Séguin-Swartz, G.; Clarke, W.E.; Parkin, I.A.P. Monolignol biosynthesis is associated with resistance to Sclerotinia sclerotiorum in Camelina sativa. Mol. Plant Pathol. 2012, 13, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi-Leonel, R.; Mueller, D.; Harbach, C.; Tylka, G.; Leandro, L. Susceptibility of cover crop plants to Fusarium virguliforme, causal agent of soybean sudden death syndrome, and Heterodera glycines, the soybean cyst nematode. J. Soil Water Conserv. 2017, 72, 575–583. [Google Scholar] [CrossRef]
- Conn, K.L.; Browne, L.M.; Tewari, J.P.; Ayer, W.A. Resistance to Rhizoctonia solani and presence of antimicrobial compounds in Camelina sativa roots. J. Plant Biochem. Biotechnol. 1994, 3, 125–130. [Google Scholar] [CrossRef]
- Paul, V.H.; Badent, S.; Henneken, M.; Foller, I. Results on the occurrence and seed-borne nature of the new disease bacterial blight (Pseudomonas syringae pv. spec) on false flax (Camelina sativa). IOBC WPRS BULLETIN 2000, 23, 43–50. [Google Scholar]
- Broadbent, L.; Heathcote, G.D. Properties and host range of turnip crinkle, rosette and yellow mosaic viruses. Ann. Appl. Biol. 1958, 46, 585–592. [Google Scholar] [CrossRef]
- Hein, A. Transmission of turnip yellow mosaic virus through seed of Camelina sativa (gold of pleasure). Z. Pflanzenkrankh. Pflanzenschutz/J. Plant Dis. Prot. 1984, 91, 549–551. [Google Scholar]
- Graichen, K. Zur Bedeutung von Virusbefall fuer denAnbau von Winterraps und Leindotter. Mitt. Biol. Bundesanst. Land-Forstwirtsch 1995, 310, 102–108. [Google Scholar]
- Chesnais, Q.; Mauck, K.E.; Bogaert, F.; Bamière, A.; Catterou, M.; Spicher, F.; Brault, V.; Tepfer, M.; Ameline, A. Virus effects on plant quality and vector behavior are species specific and do not depend on host physiological phenotype. J. Pest Sci. 2019, 92, 791–804. [Google Scholar] [CrossRef] [Green Version]
- Eberle, C.A.; Thom, M.D.; Nemec, K.T.; Forcella, F.; Lundgren, J.G.; Gesch, R.W.; Riedell, W.E.; Papiernik, S.K.; Wagner, A.; Peterson, D.H. Using pennycress, camelina, and canola cash cover crops to provision pollinators. Ind. Crops Prod. 2015, 75, 20–25. [Google Scholar] [CrossRef]
- Thom, M.D.; Eberle, C.A.; Forcella, F.; Gesch, R.; Weyers, S.; Lundgren, J.G. Nectar production in oilseeds: Food for pollinators in an agricultural landscape. Crop Sci. 2016, 56, 727–739. [Google Scholar] [CrossRef]
- Bekuzarova, S.A.; Basiev, S.S.; Buyankin, V.I.; Kozirev, A.K.; Dulaev, T.A. Ecological significance of winter camelina in biological agriculture. J. Pharm. Sci. Res. 2018, 10, 893–895. [Google Scholar]
- Wagg, C.; van Erk, A.; Fava, E.; Comeau, L.-P.; Mitterboeck, T.F.; Goyer, C.; Li, S.; McKenzie-Gopsil, A.; Mills, A. Full-season cover crops and their traits that promote agroecosystem services. Agriculture 2021, 11, 830. [Google Scholar] [CrossRef]
- Ugrenović, V.; Filipović, V.; Jevremović, S.; Marjanović-Jeromela, A.; Popović, V.; Buntić, A.; Delić, D.I. Effect of Brassicaceae as cover crops. Sel. I Semen. 2019, 25, 1–8. [Google Scholar] [CrossRef]
- Abramovič, H.; Abram, V. Physico-chemical properties, composition and oxidative stability of Camelina sativa oil. Food Technol. Biotechnol. 2005, 43, 63–70. [Google Scholar]
- Rodríguez-Rodríguez, M.F.; Sánchez-García, A.; Salas, J.J.; Garcés, R.; Martínez-Force, E. Characterization of the morphological changes and fatty acid profile of developing Camelina sativa seeds. Ind. Crops Prod. 2013, 50, 673–679. [Google Scholar] [CrossRef]
- Anderson, J.V.; Wittenberg, A.; Li, H.; Berti, M.T. High throughput phenotyping of Camelina sativa seeds for crude protein, total oil, and fatty acids profile by near infrared spectroscopy. Ind. Crops Prod. 2019, 137, 501–507. [Google Scholar] [CrossRef]
- Zubr, J. Dietary fatty acids and amino acids of Camelina sativa seed. J. Food Qual. 2003, 26, 451–462. [Google Scholar] [CrossRef]
- Zubr, J. Qualitative variation of Camelina sativa seed from different locations. Ind. Crops Prod. 2003, 17, 161–169. [Google Scholar] [CrossRef]
- Hotton, S.K.; Kammerzell, M.; Chan, R.; Hernandez, B.T.; Young, H.A.; Tobias, C.; McKeon, T.; Brichta, J.; Thomson, N.J.; Thomson, J.G. Phenotypic examination of Camelina sativa (L.) Crantz accessions from the USDA-ARS National Genetics Resource Program. Plants 2020, 9, 642. [Google Scholar] [CrossRef]
- Singer, S.D.; Zou, J.; Weselake, R.J. Abiotic factors influence plant storage lipid accumulation and composition. Plant Sci. 2016, 243, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zubr, J.; Matthaus, B. Effects of growth conditions on fatty acids and tocopherols in Camelina sativa oil. Ind. Crops Prod. 2002, 15, 155–162. [Google Scholar] [CrossRef]
- Krzyżaniak, M.; Stolarski, M.J.; Tworkowski, J.; Puttick, D.; Eynck, C.; Załuski, D.; Kwiatkowski, J. Yield and seed composition of 10 spring camelina genotypes cultivated in the temperate climate of Central Europe. Ind. Crops Prod. 2019, 138, 111443. [Google Scholar] [CrossRef]
- Yang, J.; Caldwell, C.; Corscadden, K.; He, Q.S.; Li, J. An evaluation of biodiesel production from Camelina sativa grown in Nova Scotia. Ind. Crops Prod. 2016, 81, 162–168. [Google Scholar] [CrossRef]
- Moser, B.R.; Vaughn, S.F. Evaluation of alkyl esters from Camelina sativa oil as biodiesel and as blend components in ultra low-sulfur diesel fuel. Bioresour. Technol. 2010, 101, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Leung, D.Y.C. Optimization of biodiesel production from camelina oil using orthogonal experiment. Appl. Energy 2011, 88, 3615–3624. [Google Scholar] [CrossRef]
- Hrastar, R.; Petrišič, M.G.; Ogrinc, N.; Košir, I.J. Fatty acid and stable carbon isotope characterization of Camelina sativa oil: Implications for authentication. J. Agric. Food Chem. 2009, 57, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Voelker, T.; Kinney, A.J. Variations in the biosynthesis of seed-storage lipids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 335–361. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, H.; Sun, J.; Li, B.; Zhu, Q.; Chen, S.; Zhang, H. Arabidopsis fatty acid desaturase FAD2 is required for salt tolerance during seed germination and early seedling growth. PLoS ONE 2012, 7, e30355. [Google Scholar] [CrossRef] [PubMed]
- Dar, A.A.; Choudhury, A.R.; Kancharla, P.K.; Arumugam, N. The FAD2 Gene in Plants: Occurrence, Regulation, and Role. Front Plant Sci. 2017, 8, 1789. [Google Scholar] [CrossRef] [Green Version]
- Bhunia, R.K.; Kaur, R.; Maiti, M.K. Metabolic engineering of fatty acid biosynthetic pathway in sesame (Sesamum indicum L.): Assembling tools to develop nutritionally desirable sesame seed oil. Phytochem. Rev. 2015, 15, 799–811. [Google Scholar] [CrossRef]
- Raziei, Z.; Kahrizi, D.; Rostami-Ahmadvandi, H. Effects of climate on fatty acid profile in Camelina sativa. Cell. Mol. Biol. 2018, 64, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Berti, M.T.; Wilckens, R.; Fischer, S.; Araos, R. Borage: A new crop for southern Chile. In Trends in New Crops and New Uses: Strengthin Diversity; Janick, J., Whipkey, A., Eds.; ASHS Press: Alexandria, VA, USA, 2002; pp. 501–505. [Google Scholar]
- Gilbertson, P.K.; Berti, M.T.; Johnson, B.L. Borage cardinal germination temperatures and seed development. Ind. Crops Prod. 2014, 59, 202–209. [Google Scholar] [CrossRef]
- Leclère, M.; Lorent, A.-R.; Jeuffroy, M.-H.; Butier, A.; Chatain, C.; Loyce, C. Diagnosis of camelina seed yield and quality across an on-farm experimental network. Eur. J. Agron. 2021, 122, 126190. [Google Scholar] [CrossRef]
- Borzoo, S.; Mohsenzadeh, S.; Moradshahi, A.; Kahrizi, D.; Zamani, H.; Zarei, M. Characterization of physiological responses and fatty acid compositions of Camelina sativa genotypes under water deficit stress and symbiosis with Micrococcus yunnanensis. Symbiosis 2021, 83, 79–90. [Google Scholar] [CrossRef]
- Singh, S.K.; Rajpurohit, B.; Singha, P. Camelina (Camelina sativa) seed. In Oilseeds: Health Attributes and Food Applications; Tanwar, B., Goyal, A., Eds.; Springer: Singapore, 2021; pp. 455–471. [Google Scholar]
- Karvonen, H.M.; Aro, A.; Tapola, N.S.; Salminen, I.; Uusitupa, M.I.; Sarkkinen, E.S. Effect of alpha-linolenic acid-rich Camelina sativa oil on serum fatty acid composition and serum lipids in hypercholesterolemic subjects. Metabolism 2002, 51, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Schwab, U.S.; Lankinen, M.A.; de Mello, V.D.; Manninen, S.M.; Kurl, S.; Pulkki, K.J.; Laaksonen, D.E.; Erkkilä, A.T. Camelina sativa oil, but not fatty fish or lean fish, improves serum lipid profile in subjects with impaired glucose metabolism—a randomized controlled trial. Mol. Nutr. Food Res. 2018, 62, 1700503. [Google Scholar] [CrossRef]
- Manninen, S.; Lankinen, M.; Erkkilä, A.; Nguyen, S.D.; Ruuth, M.; de Mello, V.; Öörni, K.; Schwab, U. The effect of intakes of fish and Camelina sativa oil on atherogenic and anti-atherogenic functions of LDL and HDL particles: A randomized controlled trial. Atherosclerosis 2019, 281, 56–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Mello, V.D.; Dahlman, I.; Lankinen, M.; Kurl, S.; Pitkänen, L.; Laaksonen, D.E.; Schwab, U.; Erkkilä, A.T. The effect of different sources of fish and camelina sativa oil on immune cell and adipose tissue mRNA expression in subjects with abnormal fasting glucose metabolism: A randomized controlled trial. Nutr. Diabetes 2019, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Saleh-Ghadimi, S.; Dehghan, P.; Farhangi, M.A.; Khoshbaten, M. Co-Supplementation of Camelina Oil and Prebiotic is more Effective in Improvement of Cardiometabolic Risk Factors and Mental Health in Patients with NAFLD: A Randomized Clinical Trial. Food Funct. 2021, in press. [Google Scholar] [CrossRef]
- Ergönül, P.G.; Özbek, Z.A. Cold pressed camelina (Camelina sativa L.) seed oil. In Cold Pressed Oils; Elsevier: Amsterdam, The Netherlands, 2020; pp. 255–266. [Google Scholar]
- Ibrahim, F.M.; El Habbasha, S. Chemical composition, medicinal impacts andcultivation of camelina (Camelina sativa): Review. Int. J. PharmTech Res. 2015, 8, 114–122. [Google Scholar]
- Ratusz, K.; Symoniuk, E.; Wroniak, M.; Rudzińska, M. Bioactive compounds, nutritional quality and oxidative stability of cold-pressed camelina (Camelina sativa L.) oils. Appl. Sci. 2018, 8, 2606. [Google Scholar] [CrossRef] [Green Version]
- Eidhin, D.N.; O’Beirne, D. Oxidative stability of camelina oil in salad dressings, mayonnaises and during frying. Int. J. Food Sci. + Technol. 2010, 45, 444–452. [Google Scholar] [CrossRef]
- Salminen, H.; Estévez, M.; Kivikari, R.; Heinonen, M. Inhibition of protein and lipid oxidation by rapeseed, camelina and soy meal in cooked pork meat patties. Eur. Food Res. Technol. Vol. 2006, 223, 461–468. [Google Scholar] [CrossRef]
- Cherian, G. Camelina sativa in poultry diets: Opportunities and challenges. In Biofuel Co-Products as Livestock Feed: Opportunities and Challenges; Harinder, P.S., Ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012; pp. 303–310. [Google Scholar]
- Bătrîna, Ș.L.; Jurcoane, Ș.; Popescu, I.; Marin, F.; Imbrea, I.M.; Crista, F.; Pop, G.; Imbrea, F. Camelina sativa: A study on amino acid content. Rom. Biotechnol Lett. 2020, 25, 1136–1142. [Google Scholar] [CrossRef]
- Singh, B.K.; Bala, M.; Rai, P.K. Fatty acid composition and seed meal characteristics of Brassica and allied genera. Natl. Acad. Sci. Lett. 2014, 37, 219–226. [Google Scholar] [CrossRef]
- Russo, R.; Reggiani, R. Glucosinolates and sinapine in camelina meal. Food Nutr. Sci. 2017, 8, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Paula, E.M.; da Silva, L.G.; Brandao, V.L.N.; Dai, X.; Faciola, A.P. Feeding canola, camelina, and carinata meals to ruminants. Animals 2019, 9, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas, H.; Castillejos, L.; López-Suárez, M.; Ferret, A. In vitro digestibility, in situ degradability, rumen fermentation and N metabolism of camelina co-products for beef cattle studied with a dual flow continuous culture system. Animals 2019, 9, 1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriel, P.; Nayigihugu, V.; Cappellozza, B.I.; Gonçalves, E.P.; Krall, J.M.; Foulke, T.; Cammack, K.M.; Hess, B.W. Camelina meal and crude glycerin as feed supplements for developing replacement beef heifers. J. Anim. Sci. 2011, 89, 4314–4324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombini, S.; Broderick, G.A.; Galasso, I.; Martinelli, T.; Rapetti, L.; Russo, R.; Reggiani, R. Evaluation of Camelina sativa (L.) Crantz meal as an alternative protein source in ruminant rations. J. Sci. Food Agric. 2014, 94, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, D.A.; Rohwer, G.; Norberg, O.S.; Kimura, E.; Neibergs, J.S.; Fransen, S.C. Case study: Evaluating farm processed canola and Camelina meals as protein supplements for beef cattle. J. NACAA 2015, 8. Available online: https://www.nacaa.com/journal (accessed on 10 February 2022).
- Cappellozza, B.I.; Cooke, R.F.; Bohnert, D.W.; Cherian, G.; Carroll, J.A. Effects of camelina meal supplementation on ruminal forage degradability, performance, and physiological responses of beef cattle. J. Anim. Sci. 2012, 90, 4042–4054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, R.D.; Anderson, J.L.; Clapper, J.A. Evaluation of camelina meal as a feedstuff for growing dairy heifers. J. Dairy Sci. 2016, 99, 6215–6228. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.D.; Anderson, J.L. Ruminal degradation and intestinal digestibility of camelina meal and carinata meal compared with other protein sources. Prof. Anim. Sci. 2018, 34, 10–18. [Google Scholar] [CrossRef]
- Russo, R.; Reggiani, R. Antinutritive compounds in twelve Camelina sativa genotypes. Am. J. Plant Sci. 2012, 3, 1408–1412. [Google Scholar] [CrossRef] [Green Version]
- Schill, S.R. Camelina meal approved for feedlot cattle. Biodiesel Mag. 2009. Available online: http://www.biodieselmagazine.com/articles/3837/camelinameal-approved-for-feedlot-cattle (accessed on 10 February 2022).
- Berhow, M.A.; Polat, U.; Glinski, J.A.; Glensk, M.; Vaughn, S.F.; Isbell, T.; Ayala-Diaz, I.; Marek, L.; Gardner, C. Optimized analysis and quantification of glucosinolates from Camelina sativa seeds by reverse-phase liquid chromatography. Ind. Crops Prod. 2013, 43, 119–125. [Google Scholar] [CrossRef]
- Russo, R.; Galasso, I.; Reggiani, R. Variability in glucosinolate content among Camelina species. Am. J. Plant Sci. 2014, 5, 294–298. [Google Scholar] [CrossRef] [Green Version]
- Vetter, W.; Darwisch, V.; Lehnert, K. Erucic acid in Brassicaceae and salmon–An evaluation of the new proposed limits of erucic acid in food. NFS J. 2020, 19, 9–15. [Google Scholar] [CrossRef]
- Hurtaud, C.; Peyraud, J.L. Effects of feeding camelina (seeds or meal) on milk fatty acid composition and butter spreadability. J. Dairy Sci. 2007, 90, 5134–5145. [Google Scholar] [CrossRef]
- Halmemies-Beauchet-Filleau, A.; Kokkonen, T.; Lampi, A.M.; Toivonen, V.; Shingfield, K.J.; Vanhatalo, A. Effect of plant oils and camelina expeller on milk fatty acid composition in lactating cows fed diets based on red clover silage. J. Dairy Sci. 2011, 94, 4413–4430. [Google Scholar] [CrossRef] [PubMed]
- Halmemies-Beauchet-Filleau, A.; Shingfield, K.J.; Simpura, I.; Kokkonen, T.; Jaakkola, S.; Toivonen, V.; Vanhatalo, A. Effect of incremental amounts of camelina oil on milk fatty acid composition in lactating cows fed diets based on a mixture of grass and red clover silage and concentrates containing camelina expeller. J. Dairy Sci. 2017, 100, 305–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieslak, A.; Stanisz, M.; Wojtowski, J.; Pers-Kamczyc, E.; Szczechowiak, J.; El-Sherbiny, M.; Szumacher-Strabel, M. Camelina sativa affects the fattyacid contents in M. longissimus muscle of lambs. Eur. J. Lipid Sci. Technol. 2013, 115, 1258–1265. [Google Scholar] [CrossRef]
- Steppa, R.; Cieślak, A.; Szumacher-Strabel, M.; Bielińska-Nowak, S.; Bryszak, M.; Stanisz, M.; Szkudelska, K. Blood serum metabolic profile and fatty acid composition in sheep fed concentrates with Camelina sativa cake and distillers dried grains with solubles. Small Rumin. Res. 2017, 156, 20–26. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Butler, K.L.; Muir, S.K.; Plozza, T.E.; Kerr, M.G.; Brown, W.G.; Jacobs, J.L.; Knight, M.I. Lipid oxidation and colour stability of lamb and yearling meat (muscle longissimus lumborum) from sheep supplemented with Camelina-based diets after short-, medium-, and long-term storage. Antioxidants 2021, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Szumacher-Strabel, M.; Cieślak, A.; Zmora, P.; Pers-Kamczyc, E.; Bielińska, S.; Stanisz, M.; Wójtowski, J. Camelina sativa cake improved unsaturated fatty acids in ewe’s milk. J. Sci. Food Agric. 2011, 91, 2031–2037. [Google Scholar] [CrossRef]
- Mierlita, D.; Daraban, S.; Lup, F.; Chereji, A. The effect of grazing management and camelina seed supplementation in the diet on milk performance and milk fatty acid composition of dairy ewes. J. Food Agric. Environ. 2011, 9, 368–373. [Google Scholar]
- Pikul, J.; Wójtowski, J.; Danków, R.; Teichert, J.; Czyżak-Runowska, G.; Cais-Sokolińska, D.; Cieślak, A.; Szumacher-Strabel, M.; Bagnicka, E. The effect of false flax (Camelina sativa) cake dietary supplementation in dairy goats on fatty acid profile of kefir. Small Rumin. Res. 2014, 122, 44–49. [Google Scholar] [CrossRef]
- Cais-Sokolińska, D.; Majcher, M.; Pikul, J.; Bielińska, S.; Czauderna, M.; Wójtowski, J. The effect of Camelina sativa cake diet supplementation on sensory and volatile profiles of ewe’s milk. Afr. J. Biotechnol. 2011, 10, 7245–7252. [Google Scholar] [CrossRef]
- Cais-Sokolińska, D.; Wójtowski, J.; Pikul, J.; Danków, R.; Majcher, M.; Teichert, J.; Bagnicka, E. Formation of volatile compounds in kefir made of goat and sheep milk with high polyunsaturated fatty acid content. J. Dairy Sci. 2015, 98, 6692–6705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mierlita, D.; Vicas, S. Dietary effect of silage type and combination with camelina seed on milk fatty acid profile and antioxidant capacity of sheep milk. S. Afr. J. Anim. Sci. 2015, 45, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Christodoulou, C.; Mavrommatis, A.; Mitsiopoulou, C.; Symeon, G.; Dotas, V.; Sotirakoglou, K.; Kotsampasi, B.; Tsiplakou, E. Assessing the optimum level of supplementation with Camelina seeds in ewes’ diets to improve milk quality. Foods 2021, 10, 2076. [Google Scholar] [CrossRef] [PubMed]
- Danków, R.; Pikul, J.; Wójtowski, J.; Cais-Sokolińska, D.; Teichert, J.; Bagnicka, E.; Cieślak, A.; Szumacher-Strabel, M. The effect of supplementation with gold of pleasure (Camelina sativa) cake on the fatty acid profile of ewe milk and yoghurt produced from it. J. Anim. Feed Sci. 2015, 24, 193–202. [Google Scholar] [CrossRef]
- Mierlita, D. Conjugated linoleic acid and ω-3 fatty acid in sheep milk was increased by part-time grazing and camelina seed diet. J. Food Agric. Environ. 2015, 13, 76–81. [Google Scholar]
- Eidhin, D.N.; Burke, J.; Lynch, B.; O’Beirne, D. Effects of dietary supplementation with camelina oil on porcine blood lipids. J. Food Sci. 2003, 68, 671–679. [Google Scholar] [CrossRef]
- Hăbeanu, M.; Hebean, V.; Țăranu, I.; Marin, D.; Ropotă, M.; Tamaș, V. Effects of the dietary Camelina oil on large white pigs’ meat quality. Arch. Zootech. 2009, 12, 31–45. [Google Scholar]
- Hăbeanu, M.; Hebean, V.; Taranu, I.; Ropota, M.; Lefter, N.; Marin, D. Dietary ecologic Camelina oil: A beneficial aource of n-3 PUFA in muscle tissue and health status in finishing pigs. Rom. Biotechnol. Lett. 2011, 16, 5564–5571. [Google Scholar]
- Taranu, I.; Gras, M.; Pistol, G.C.; Motiu, M.; Marin, D.E.; Lefter, N.; Ropota, M.; Habeanu, M. ω-3 PUFA rich Camelina oil by-products improve the systemic metabolism and spleen cell functions in fattening pigs. PLoS ONE 2014, 9, e110186. [Google Scholar] [CrossRef] [PubMed]
- Almeida, F.N.; Htoo, J.K.; Thomson, J.; Stein, H.-H. Amino acid digestibility in camelina products fed to growing pigs. Can. J. Anim. Sci. 2013, 93, 335–343. [Google Scholar] [CrossRef]
- Kahindi, R.K.; Woyengo, T.A.; Thacker, P.A.; Nyachoti, C.M. Energy and amino acid digestibility of camelina cake fed to growing pigs. Anim. Feed Sci. Technol. 2014, 193, 93–101. [Google Scholar] [CrossRef]
- Hăbeanu, M.; Lefter, A.N.; Ropotă, M. Effects of the camelina cake on fatty acids composition in different organs of the pigs. Arch. Zootech. 2016, 19, 5–16. [Google Scholar]
- Smit, M.N.; Beltranena, E. Effects of feeding camelina cake to weaned pigs on safety, growth performance, and fatty acid composition of pork. J. Anim. Sci. 2017, 95, 2496–2508. [Google Scholar] [CrossRef] [PubMed]
- Meadus, W.J.; Duff, P.; McDonald, T.; Caine, W.R. Pigs fed camelina meal increase hepatic gene expression of cytochrome 8b1, aldehyde dehydrogenase, and thiosulfate transferase. J. Anim. Sci. Biotechnol. 2014, 5, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilbrands, A.M.; Johnston, L.J.; Cox, R.B.; Forcella, F.; Gesch, R.; Li, Y.Z. Effects of increasing dietary inclusion of camelina cake on growth performance of growing-finishing pigs. Transl. Anim. Sci. 2021, 5, 1–10. [Google Scholar] [CrossRef]
- Kim, J.W.; Koo, B.; Nyachoti, C.M. Digestible, metabolizable, and net energy of camelina cake fed to growing pigs and additivity of energy in mixed diets. J. Anim. Sci. 2017, 95, 4037–4044. [Google Scholar] [CrossRef]
- Juodka, R.; Nainienė, R.; Juškienė, V.; Juška, R.; Leikus, R.; Kadžienė, G.; Stankevičienė, D. Camelina (Camelina sativa (L.) Crantz) as feedstuffs in meat type poultry diet: A source of protein and n-3 fatty acids. Animals 2022, 12, 295. [Google Scholar] [CrossRef]
- Aziza, A.E.; Quezada, N.; Cherian, G. Feeding Camelina sativa meal to meat-type chickens: Effect on production performance and tissue fatty acid composition. J. Appl. Poult. Res. 2010, 19, 157–168. [Google Scholar] [CrossRef]
- Aziza, A.; Panda, A.; Quezada, N.; Cherian, G. Nutrient digestibility, egg quality, and fatty acid composition of brown laying hens fed camelina or flaxseed meal. J. Appl. Poult. Res. 2013, 22, 832–841. [Google Scholar] [CrossRef]
- Nain, S.; Oryschak, M.A.; Betti, M.; Beltranena, E. Camelina sativa cake for broilers: Effects of increasing dietary inclusion from 0 to 24% on tissue fatty acid proportions at 14, 28, and 42 d of age. Poult. Sci. 2015, 94, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Woyengo, T.A.; Patterson, R.; Slominski, B.A.; Beltranena, E.; Zijlstra, R.T. Nutritive value of cold-pressed camelina cake with or without supplementation of multi-enzyme in broiler chickens. Poult. Sci. 2016, 95, 2314–2321. [Google Scholar] [CrossRef] [PubMed]
- Jáskiewicz, T.; Sagan, A.; Puzio, I. Effect of the Camelina sativa oil on theperformance, essential fatty acid level in tissues and fat-soluble vitamins content in the livers of broiler chickens. Livest. Sci. 2014, 165, 74–79. [Google Scholar] [CrossRef]
- Pietras, M.P.; Orczewska-Dudek, S. The effect of dietary Camelina sativa oil on quality of broiler chicken meat. Ann. Anim. Sci. 2013, 13, 869–882. [Google Scholar] [CrossRef]
- Orczewska-Dudek, S.; Pietras, M. The effect of dietary Camelina sativa oil or cake in the diets of broiler chickens on growth performance, fatty acid profile, and sensory quality of meat. Animals 2019, 9, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juodka, R.; Juska, R.; Juskiene, V.; Leikus, R.; Stankeviciene, D.; Nainiene, R. The effect of feeding with hemp and Camelina cakes on the fatty acid profile of duck muscles. Arch. Anim. Breed. 2018, 61, 293–303. [Google Scholar] [CrossRef]
- Frame, D.D.; Palmer, M.; Peterson, B. Use of Camelina sativa in the diets of young turkeys. J. Appl. Poult. Res. 2007, 16, 381–386. [Google Scholar] [CrossRef]
- Pekel, A.Y.; Kim, J.I.; Chapple, C.; Adeola, O. Nutritional characteristics of camelina meal for 3-week-old broiler chickens. Poult Sci 2015, 94, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Ryhänen, E.L.; Perttilä, S.; Tupasela, T.; Valaja, J.; Eriksson, C.; Larkka, K.E. Effect of Camelina sativa expeller cake on performance and meat quality of broilers. J. Sci. Food Agric. 2007, 87, 1489–1494. [Google Scholar] [CrossRef]
- Bulbul, T.; Rahman, A.; Ozdemir, V. Effect of false flax meal on certain growth, serum and meat parameters of japanese quails. J. Anim. Plant Sci. 2015, 25, 1245–1250. [Google Scholar]
- Ye, C.L.; Anderson, D.M.; Lall, S.P. The effects of camelina oil and solvent extracted camelina meal on the growth: Carcass composition and hindguthistology of Atlantic salmon (Salmo salar) parr in freshwater. Aquaculture 2016, 450, 397–404. [Google Scholar] [CrossRef]
- Hixson, S.M.; Parrish, C.C.; Anderson, D.M. Full substitution of fish oil with camelina (Camelina sativa) oil, with partial substitution of fish meal with camelina meal, in diets for farmed Atlantic salmon (Salmo salar) and its effect on tissue lipids and sensory quality. Food Chem. 2014, 157, 51–61. [Google Scholar] [CrossRef]
- Hixson, S.M.; Parrish, C.C.; Anderson, D.M. Effect of replacement of fish oil with camelina (Camelina sativa) oil on growth, lipid class and fatty acid composition of farmed juvenile Atlantic cod (Gadus morhua). Fish Physiol. Biochem. 2013, 39, 1441–1456. [Google Scholar] [CrossRef] [PubMed]
- Hixson, S.M.; Parrish, C.C.; Wells, J.S.; Marie Winkowski, E.; Anderson, D.M. Inclusion of camelina meal as a protein source in diets for farmed Atlantic cod Gadus morhua. Aquac. Res. 2016, 47, 2607–2622. [Google Scholar] [CrossRef]
- Ruiz-Lopez, N.; Haslam, R.P.; Napier, J.A.; Sayanova, O. Successful high-level accumulation of fish oil omega-3 long-chain polyunsaturated fatty acids in a transgenic oilseed crop. Plant J. 2014, 77, 198–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betancor, M.B.; MacEwan, A.; Sprague, M.; Gong, X.; Montero, D.; Han, L.; Napier, J.A.; Norambuena, F.; Izquierdo, M.; Tocher, D.R. Oil from transgenic Camelina sativa as a source of EPA and DHA in feed for European sea bass (Dicentrarchus labrax L.). Aquaculture 2021, 530, 735759. [Google Scholar] [CrossRef] [PubMed]
- Bullerwell, C.N.; Collins, S.A.; Lall, S.P.; Anderson, D.M. Growth performance, proximate and histological analysis of rainbow trout fed diets containing Camelina sativa seeds, meal (high-oil and solvent-extracted) and oil. Aquaculture 2016, 452, 342–350. [Google Scholar] [CrossRef]
- Liang, Y.; May, C.; Foon, C.; Ngan, M.; Hock, C.; Basiron, Y. The effect of natural and synthetic antioxidants on the oxidative stability of palm diesel. Fuel 2006, 85, 867–870. [Google Scholar] [CrossRef]
- ASTM. ASTM (D6751–15ce1) Standard Specification for Biodiesel Fuel BlendStock (B100) for Middle Distillate Fuels. Am. Soc. Test. Mater. 2015. Available online: https://dieselnet.com/tech/fuel_biodiesel_std.php (accessed on 10 February 2022).
- European Standard EN 14214. EN (14214) European Specifications for Biodiesel Fuels. Eur. Comm. Stand. 2012. Available online: https://en.wikipedia.org/wiki/EN_14214 (accessed on 10 February 2022).
- Soriano, N.U.; Narani, A. Evaluation of biodiesel rerived from Camelina sativa oil. J. Am. Oil Chem. Soc. 2011, 89, 917–923. [Google Scholar] [CrossRef]
- Miller, P.; Kumar, A. Techno-economic assessment of hydrogenation-derived renewable diesel production from canola and camelina. Sustain. Energy Technol. Assess. 2014, 6, 105–115. [Google Scholar] [CrossRef]
- Miller, P.; Kumar, A. Development of emission parameters and net energy ratio for renewable diesel from Canola and Camelina. Energy 2013, 58, 426–437. [Google Scholar] [CrossRef]
- Krohn, B.J.; Fripp, M. A life cycle assessment of biodiesel derived from the “niche filling” energy crop camelina in the USA. Appl. Energy 2012, 92, 92–98. [Google Scholar] [CrossRef]
- Corporan, E.; Edwards, T.; Shafer, L.; DeWitt, M.J.; Klingshirn, C.; Zabarnick, S.; West, Z.; Striebich, R.; Graham, J.; Klein, J. Chemical, thermal stability, sealswell, and emissions studies of alternative jet fuels. Energy Fuels 2011, 25, 955–966. [Google Scholar] [CrossRef]
- Sivakumar, D.; Vankeswaram, S.; Sakthikumar, R.; Raghunandan, B. Analysis on the atomization characteristics of aviation biofuel discharging from simplex swirl atomizer. Int. J. Multiph. Flow 2015, 72, 88–96. [Google Scholar] [CrossRef]
- Sivakumar, G.; Vail, D.R.; Xu, J.; Burner, D.M.; Lay, J.O.; Ge, X.; Weathers, P.J. Bioethanol and biodiesel: Alternative liquid fuels for future generations. Eng. Life Sci. 2010, 10, 8–18. [Google Scholar] [CrossRef]
- Agusdinata, D.B.; Zhao, F.; Ileleji, K.; DeLaurentis, D. Life cycle assessment of potential biojet fuel production in the United States. Environ. Sci. Technol. 2011, 45, 9133–9143. [Google Scholar] [CrossRef] [PubMed]
- Shonnard, D.R.; Williams, L.; Kalnes, T.N. Camelina-derived jet fuel and diesel: Sustainable advanced biofuels. Env. Prog Sustain Energy 2010, 29, 382–392. [Google Scholar] [CrossRef]
- Shila, J.; Johnson, M.E. Techno-economic analysis of Camelina-derived hydroprocessed renewable jet fuel within the US context. Appl. Energy 2021, 287, 116525. [Google Scholar] [CrossRef]
- Boateng, A.A.; Mullen, C.A.; Goldberg, N.M. Producing stable pyrolysis liquids from the oil-seed presscakes of mustard family plants: Pennycress (Thlaspi arvense L.) and Camelina (Camelina sativa). Energy Fuels 2010, 24, 6624–6632. [Google Scholar] [CrossRef]
- Hernando, H.; Fermoso, J.; Moreno, I.; Coronado, J.M.; Serrano, D.P.; Pizarro, P. Thermochemical valorization of camelina straw waste via fast pyrolysis. Biomass Convers. Biorefinery 2017, 7, 277–287. [Google Scholar] [CrossRef]
- Gómez-Monedero, B.; Bimbela, F.; Arauzo, J.; Faria, J.; Ruiz, M.P. Pyrolysis of red eucalyptus, camelina straw, and wheat straw in an ablative reactor. Energy Fuels 2015, 29, 1766–1775. [Google Scholar] [CrossRef]
- Mohammad, B.T.; Al-Shannag, M.; Alnaief, M.; Singh, L.; Singsaas, E.; Alkasrawi, M. Production of multiple biofuels from whole Camelina material: A renewable energy crop. BioResources 2018, 13, 4870–4883. [Google Scholar]
- Jankowski, K.J.; Sokólski, M. Spring camelina: Effect of mineral fertilization on the energy efficiency of biomass production. Energy 2021, 220, 119731. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Krzyżaniak, M.; Kwiatkowski, J.; Tworkowski, J.; Szczukowski, S. Energy and economic efficiency of camelina and crambe biomass production on a large-scale farm in north-eastern Poland. Energy 2018, 150, 770–780. [Google Scholar] [CrossRef]
- Kasetaite, S.; Ostrauskaite, J.; Grazuleviciene, V.; Svediene, J.; Bridziuviene, D. Camelina oil- and linseed oil-based polymers with bisphosphonate crosslinks. Appl. Polym. 2014, 131, 8536–8543. [Google Scholar] [CrossRef]
- Fox, N.J.; Stachowiak, G.W. Vegetable oil-based lubricants- a review of oxidation. Tribol. Int. 2007, 40, 1035–1046. [Google Scholar] [CrossRef]
- Knothe, G. Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Processing Technol. 2005, 86, 1059–1070. [Google Scholar] [CrossRef]
- Kim, N.; Li, Y.; Sun, X.S. Epoxidation of Camelina sativa oil and peel adhesion properties. Ind. Crops Prod. 2015, 64, 1–8. [Google Scholar] [CrossRef]
- Nosal, H.; Nowicki, J.; Warzała, M.; Nowakowska-Bogdan, E.; Zarębska, M. Synthesis and characterization of alkyd resins based on Camelina sativa oil and polyglycerol. Prog. Org. Coat. 2015, 86, 59–70. [Google Scholar] [CrossRef]
- Kim, J.T.; Netravali, A.N. Non-food application of camelina meal: Development of sustainable and green biodegradable paper-camelina composite sheets and fibers. Polym. Compos. 2012, 33, 1969–1976. [Google Scholar] [CrossRef]
- Cao, Y.; Gu, Z.; Muthukumarappan, K.; Gibbons, W. Separation of glucosinolates from Camelina seed meal via membrane and acidic aluminum oxide column. J. Liq. Chromatogr. Relat. Technol. 2015, 38, 1273–1278. [Google Scholar] [CrossRef]
- Ma, Y.; Gentry, T.; Hu, P.; Pierson, E.; Gu, M.; Yin, S. Impact of brassicaceous seed meals on the composition of the soil fungal community and the incidence of Fusarium wilt on chili pepper. Appl. Soil Ecol. 2015, 90, 41–48. [Google Scholar] [CrossRef]
- Schuster, A.; Friedt, W. Glucosinolate content and composition as parameters of quality of Camelina seed. Ind. Crops Prod. 1998, 7, 297–302. [Google Scholar] [CrossRef]
- Matthäus, B.; Zubr, J. Variability of specific components in Camelina sativa oilseed cakes. Ind. Crops Prod. 2000, 12, 9–18. [Google Scholar] [CrossRef]
- Maršalkienė, N.; Žilėnaitė, L.; Karpavičienė, B. Oil content and composition in seeds of Camelina sativa and Crambe abyssinica cultivars. J. Ementol. 2020, 25, 1399–1412. [Google Scholar] [CrossRef]
- Zubr, J. Unique dietary oil from Camelina sativa seed. Agrafood Ind. Hi-Tech 2009, 20, 42–46. [Google Scholar]
- De Lorgeril, M.; Renaud, S.; Salen, P.; Monjaud, I.; Mamelle, N.; Martin, J.; Guidollet, J.; Touboul, P.; Delaye, J. Mediterranean alpha-linolenic acid-rich diet in secondary prevention of coronary heart disease. Lancet 1994, 343, 1454–1459. [Google Scholar] [CrossRef]
- Budin, J.T.; Breene, W.M.; Putnam, D.H. Some compositional properties of camelina (Camelina sativa L. Crantz) seeds and oils. J. Am. Oil Chem. Soc. 1995, 72, 309–315. [Google Scholar] [CrossRef]
- Onyilagha, J.; Bala, A.; Hallett, R.; Gruber, M.; Soroka, J.; Westcott, N. Leaf flavonoids of the cruciferous species, Camelina sativa, Crambe spp., Thlaspi arvense and several other genera of the family Brassicaceae. Biochem. Syst. Ecol. 2003, 31, 1309–1322. [Google Scholar] [CrossRef]
- Kumar, K.; Gupta, S.M.; Arya, M.C.; Nasim, M. In vitro antimicrobial and antioxidant activity of camelina seed extracts as potential source of bioactive compounds. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 521–526. [Google Scholar] [CrossRef]
- Kurasiak-Popowska, D.; Ryńska, B.; Stuper-Szablewska, K. Analysis of distribution of selected bioactive compounds in Camelina sativa from seeds to pomace and oil. Agronomy 2019, 9, 168. [Google Scholar] [CrossRef] [Green Version]
- Cojocariu, R.O.; Balmus, I.-M.; Lefter, R.; Hritcu, L.; Ababei, D.C.; Ciobica, A.; Copaci, S.; Mot, S.E.L.; Copolovici, L.; Copolovici, D.M.; et al. Camelina sativa methanolic and ethanolic extract potential in alleviating oxidative stress, memory deficits, and affective impairments in stress exposure-based irritable bowel syndrome mouse models. Oxidative Med. Cell. Longev. 2020, 2020, 9510305. [Google Scholar] [CrossRef] [PubMed]
- Cojocariu, R.; Balmus, I.; Lefter, R.; Hritcu, L.; Ababei, D.; Ciobica, A.; Copaci, S.; Copolovici, L.; Copolovici, D.; Jurcoane, S. Beneficial effects of Camelina sativa oil on behavioural (memory, anxiety, depression and social-related) manifestations and oxidative stress parameters in a mice model of irritable bowel syndrome. Rom. Biotechnol. Lett. 2020, 25, 1532–1540. [Google Scholar] [CrossRef]
- Gao, L.; Caldwell, C.D.; Jiang, Y. Photosynthesis and growth of camelina and canola in response to water deficit and applied nitrogen. Crop Sci. 2018, 58, 393–401. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Ahmad, R.; Ahmed, Z.; Ahmad, Z.; Barutcular, C.; Erman, M.; Cig, F.; Saneoka, H.; Öztürk, F. Comparative study of growth, physiology and yield attributes of camelina (Camelina sativa L.) and canola (Brassica napus L.) under different irrigation regimes. Pak. J. Bot. 2020, 52, 1537–1544. [Google Scholar] [CrossRef]
- Yadav, P.V.; Khatri, D.; Nasim, M. Salt and PEG induced osmotic stress tolerance at germination and seedling stage in Camelina sativa: A potential biofuel crop. J. Seed Sci. 2017, 10, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, Z.; Waraich, E.A.; Rashid, A.; Shahbaz, M. Morpho-physiological and biochemical responses of camelina (Camelina sativa Crantz) genotypes under drought stress. Int. J. Agric. Biol. 2017, 19, 1–7. [Google Scholar] [CrossRef]
- Čanak, P.; Jeromela, A.M.; Vujošević, B.; Kiprovski, B.; Mitrović, B.; Alberghini, B.; Facciolla, E.; Monti, A.; Zanetti, F. Is drought stress tolerance affected by biotypes and seed size in the emerging oilseed crop Camelina? Agronomy 2020, 10, 1856. [Google Scholar] [CrossRef]
- Ahmad, Z.; Anjum, S.; Skalicky, M.; Waraich, E.A.; Tariq, R.M.S.; Ayub, M.A.; Hossain, A.; Hassan, M.M.; Brestic, M.; Islam, M.S.; et al. Selenium alleviates the adverse effect of drought in oilseed crops Camelina (Camelina sativa L.) and Canola (Brassica napus L.). Molecules 2021, 26, 1699. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, Z.; Waraich, E.A.; Barutcular, C.; Alharby, H.; Bamagoos, A.; Kizilgeci, F.; Öztürk, F.; Hossain, A.; Bayoumi, Y.; El Sabagh, A. Enhancing drought tolerance in Camelina sativa L. and canola (Brassica napus L.) through application of selenium. Pak. J. Bot. 2020, 52, 1927–1939. [Google Scholar] [CrossRef]
- Ahmad, M.; Waraich, E.A.; Zulfiqar, U.; Ullah, A.; Farooq, M. Thiourea application improves heat tolerance in camelina (Camelina sativa L. Crantz) by modulating gas exchange, antioxidant defense and osmoprotection. Ind. Crops Prod. 2021, 170, 113826. [Google Scholar] [CrossRef]
- Liang, C.; Liu, X.; Yiu, S.-M.; Lim, B.L. De novo assembly and characterization of Camelina sativa transcriptome by paired-end sequencing. BMC Genom. 2013, 14, 146. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.T.; Silva, J.E.; Podicheti, R.; Macrander, J.; Yang, W.; Nazarenus, T.J.; Nam, J.W.; Jaworski, J.G.; Lu, C.; Scheffler, B.E.; et al. Camelina seed transcriptome: A tool for meal and oil improvement and translational research. Plant Biotechnol. J. 2013, 11, 759–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, K.; Li, H.; Kang, J.; Lu, C. Mapping quantitative trait loci for seed traits in Camelina sativa. Theor. Appl. Genet. 2019, 132, 2567–2577. [Google Scholar] [CrossRef]
- Luo, Z.; Brock, J.; Dyer, J.M.; Kutchan, T.; Schachtman, D.; Augustin, M.; Ge, Y.; Fahlgren, N.; Abdel-Haleem, H. Genetic diversity and population structure of a Camelina sativa spring panel. Front. Plant Sci. 2019, 10, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Szczepanek, A.; Abdel-Haleem, H. Genome-wide association study (GWAS) analysis of Camelina seedling germination under salt stress sondition. Agronomy 2020, 10, 1444. [Google Scholar] [CrossRef]
- Vollmann, J.; A, D.; Baumgartner, S.; Ruckenbauer, P. Selection of induced mutants with improved linolenic acid content in camelina. Fett/Lipid 1997, 99, 357–361. [Google Scholar] [CrossRef]
- Lohaus, R.H.; Zager, J.J.; Kosma, D.K.; Cushman, J.C. Characterization of seed, oil, and fatty acid methyl esters of an ethyl methanesulfonate mutant of Camelina sativa with reduced seed-coat mucilage. J. Am. Oil Chem. Soc. 2020, 97, 157–174. [Google Scholar] [CrossRef]
- Lovett, J.V.; Sagar, G.R. Influence of bacteria in the phyllosphere of Camelina sativa (L.) Crantz on germination of Linum usitatissimum L. New Phytol. 1978, 81, 617–625. [Google Scholar] [CrossRef]
- Miyashita, H.; Fujita, K. Improvement of Camelina sativa yield using a plant growth-promoting rhizobacterium. Soil Sci. Plant Nutr. 2019, 65, 566–569. [Google Scholar] [CrossRef]
- Bekuzarova, S.A.; Weisfeld, L.I.; Dulaev, T.A. Biological methods increasing the productivity of the winter false flax Camelina silvestris L. In Antioxidants in Systems of Varying Complexity; Apple Academic Press: Palm Bay, FL, USA, 2019; pp. 293–304. [Google Scholar]
- Chhikara, S.; Abdullah, H.M.; Akbari, P.; Schnell, D.; Dhankher, O.P. Engineering Camelina sativa (L.) Crantz for enhanced oil and seed yields by combining diacylglycerol acyltransferase1 and glycerol-3-phosphate dehydrogenase expression. Plant Biotechnol J. 2018, 16, 1034–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heydarian, Z.; Yu, M.; Gruber, M.; Glick, B.R.; Zhou, R.; Hegedus, D.D. Inoculation of soil with plant growth promoting bacteria producing 1-aminocyclopropane-1-carboxylate deaminase or expression of the corresponding acdS gene in transgenic plants increases salinity tolerance in Camelina sativa. Front. Microbiol. 2016, 7, 1966. [Google Scholar] [CrossRef]
- Heydarian, Z.; Gruber, M.; Glick, B.R.; Hegedus, D.D. Gene expression patterns in roots of Camelina sativa with enhanced salinity tolerance arising from inoculation of soil with plant growth promoting bacteria producing 1-aminocyclopropane-1-carboxylate deaminase or expression the corresponding acds gene. Front. Microbiol. 2018, 9, 1297. [Google Scholar] [CrossRef] [PubMed]
- Kanth, B.K.; Kumari, S.; Choi, S.H.; Ha, H.-J.; Lee, G.-J. Generation and analysis of expressed sequence tags (ESTs) of Camelina sativa to mine drought stress-responsive genes. Biochem. Biophys. Res. Commun. 2015, 467, 83–93. [Google Scholar] [CrossRef]
- Heydarian, Z.; Yu, M.; Gruber, M.; Coutu, C.; Robinson, S.J.; Hegedus, D.D. Changes in gene expression in Camelina sativa roots and vegetative tissues in response to salinity stress. Sci. Rep. 2018, 8, 9804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.B.; Kim, H.; Kim, R.J.; Suh, M.C. Overexpression of Arabidopsis MYB96 confers drought resistance in Camelina sativa via cuticular wax accumulation. Plant Cell Rep. 2014, 33, 1535–1546. [Google Scholar] [CrossRef]
- Kim, R.J.; Kim, H.U.; Suh, M.C. Development of camelina enhanced with drought stress resistance and seed oil production by co-overexpression of MYB96A and DGAT1C. Ind. Crops Prod. 2019, 138, 111475. [Google Scholar] [CrossRef]
- Subedi, U.; Jayawardhane, K.N.; Pan, X.; Ozga, J.; Chen, G.; Foroud, N.A.; Singer, S.D. The potential of genome editing for improving seed oil content and fatty acid composition in oilseed crops. Lipids 2020, 55, 495–512. [Google Scholar] [CrossRef] [PubMed]
- Kawall, K. Genome-edited Camelina sativa with a unique fatty acid content and its potential impact on ecosystems. Environ. Sci. Eur. 2021, 33, 38. [Google Scholar] [CrossRef]
- Yuan, L.; Li, R. Metabolic engineering a model oilseed Camelina sativa for the sustainable production of high-value designed oils. Front. Plant Sci. 2020, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- Aznar-Moreno, J.A.; Durrett, T.P. Simultaneous targeting of multiple gene homeologs to alter seed oil production in Camelina sativa. Plant Cell Physiol. 2017, 58, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-R.; Jeon, I.; Yu, H.; Kim, S.-G.; Kim, H.-S.; Ahn, S.-J.; Lee, J.; Lee, S.-K.; Kim, H.U. Increasing monounsaturated fatty acid contents in hexaploid Camelina sativa seed oil by FAD2 gene knockout using CRISPR-Cas9. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Lyzenga, W.J.; Harrington, M.; Bekkaoui, D.; Wigness, M.; Hegedus, D.D.; Rozwadowski, K.L. CRISPR/Cas9 editing of three CRUCIFERIN C homoeologues alters the seed protein profile in Camelina sativa. BMC Plant Biol. 2019, 19, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veljković, V.B.; Kostić, M.D.; Stamenković, O.S. Camelina seed harvesting, storing, pretreating, and processing to recover oil: A review. Ind. Crops Prod. 2022, 178, 114539. [Google Scholar] [CrossRef]
- Pagnotta, E.; Ugolini, L.; Matteo, R.; Lazzeri, L.; Foschi, L.; Angelini, L.; Tavarini, S. Exploring the Camelina sativa value chain: A new opportunity for bio-based products and overall crop sustainability. La Riv. Italliana Delle Sostanze Grasse 2019, 96, 259–268. [Google Scholar]
- Righini, D.; Zanetti, F.; Monti, A. The bio-based economy can serve as the springboard for camelina and crambe to quit the limbo. Oilseeds Fats Crops Lipids (OCL) 2016, 23, D504. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Waraich, E.A.; Zulfiqar, U.; Ullah, A.; Farooq, M. Thiourea application increases seed and oil yields in Camelina under heat stress by modulating the plant water relations and antioxidant defense system. J. Soil Sci. Plant Nutr. 2022, 1–18. [Google Scholar] [CrossRef]
Figure 1.
Average seed yield (kg ha−1), oil (%), and protein content (%) of C. sativa grown at various locations across the world.
Figure 1.
Average seed yield (kg ha−1), oil (%), and protein content (%) of C. sativa grown at various locations across the world.

Figure 2.
Diverse uses of C. sativa adapted from Chaturvedi et al., 2017 [17]. Created with BioRender.com.
Figure 2.
Diverse uses of C. sativa adapted from Chaturvedi et al., 2017 [17]. Created with BioRender.com.

Table 2.
Fatty acid profiles of C. sativa seed oil reported by various researchers. Bold font (bottom row) indicates mean values of fatty acid profile in C. sativa from the different reports. Dashes indicate missing data.
Table 2.
Fatty acid profiles of C. sativa seed oil reported by various researchers. Bold font (bottom row) indicates mean values of fatty acid profile in C. sativa from the different reports. Dashes indicate missing data.
| Concentration of Major Fatty Acids (%) | Reference | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C16:0 | C18:0 | C18:1 | C18:2 | C18:3 | C20:1 | C22:1 | SFA | MUFA | PUFA | |
| - | - | - | 21–23 | 27–29 | - | - | 11–12 | 35–36 | 51–52 | [75] |
| - | - | 14–16 | 15–23 | 31–40 | 12–15 | - | - | - | - | [2] |
| 5.3–5.6 | 2.2–2.7 | 14.7–16.5 | 12.9–16.3 | 35 | 15 | 3 | 12 | 34 | 54 | [159,160] |
| - | - | - | - | 30–43 | 11–19 | <3 | - | - | - | [6] |
| 5.4 | 2.4 | 14.3 | 20.6 | 36.9 | 13 | 2.2 | - | - | - | [3] |
| 6.29 | 2.73 | 16.5 | 17.7 | 32.5 | 15.6 | 3.1 | - | - | - | [64] |
| 5.16 | 2.68 | 15.2 | 17.9 | 34.6 | 15.1 | 2.6 | 8.6 | 33.0 | 54.1 | [26] |
| 5.68 | - | 13.9 | 16.6 | 35.1 | 14.3 | 3.0 | 10.1 | 33.5 | 55.9 | [164] |
| - | - | 9.1–22.1 | 15.2–27.1 | 22.8–38.4 | 11.6–18.2 | - | - | - | - | [158] |
| 5.5 | 2.4 | 14.4 | 19.1 | 33.5 | 15 | 3.1 | - | - | - | [165] |
| 7 | 2.5 | 6.9 | 14.5 | 41 | 10.9 | 3.5 | - | - | - | [157] |
| 6.8 | 2.7 | 18.6 | 19.6 | 32.6 | 12.4 | 2.3 | - | - | - | [166] |
| 5.1 | 2.4 | 17.6 | 18.7 | 28.6 | 11.9 | 4.2 | - | - | - | [167] |
| 5.4 | 2.6 | 14.3 | 14.3 | 38.4 | 16.8 | 2.9 | - | - | - | [25] |
| 5.7 | 3.4 | 15.0 | 18.5 | 34.7 | 12.7 | 3.2 | - | - | - | [168] |
| 4.6–5.2 | 2.2–2.5 | 12.8–14.7 | 16.3–17.2 | 36.2–39.4 | 14.0–15.5 | 2.5–3.1 | 8.7–8.8 | - | - | [39] |
| 5.5–9.5 | - | 9.1–17.1 | 16.1–28.6 | 23.5–36.2 | 10.5–16.4 | <2 | - | - | - | [161] |
| 6.2 | 2.6 | 16.2 | 17.5 | 37.3 | 13.1 | 2.3 | 9.7 | 31.7 | 57.7 | [71] |
| 5.8 | 2.6 | 14.7 | 18.3 | 34.4 | 14.0 | 2.9 | 10.1 | 33.5 | 54.6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Neupane, D.; Lohaus, R.H.; Solomon, J.K.Q.; Cushman, J.C. Realizing the Potential of Camelina sativa as a Bioenergy Crop for a Changing Global Climate. Plants 2022, 11, 772. https://doi.org/10.3390/plants11060772
AMA Style
Neupane D, Lohaus RH, Solomon JKQ, Cushman JC. Realizing the Potential of Camelina sativa as a Bioenergy Crop for a Changing Global Climate. Plants. 2022; 11(6):772. https://doi.org/10.3390/plants11060772
Chicago/Turabian StyleNeupane, Dhurba, Richard H. Lohaus, Juan K. Q. Solomon, and John C. Cushman. 2022. "Realizing the Potential of Camelina sativa as a Bioenergy Crop for a Changing Global Climate" Plants 11, no. 6: 772. https://doi.org/10.3390/plants11060772
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.