
Machine Learning for Plant Stress Modeling: A Perspective towards Hormesis Management

 , , and
, , and
Abstract
:1. Introduction
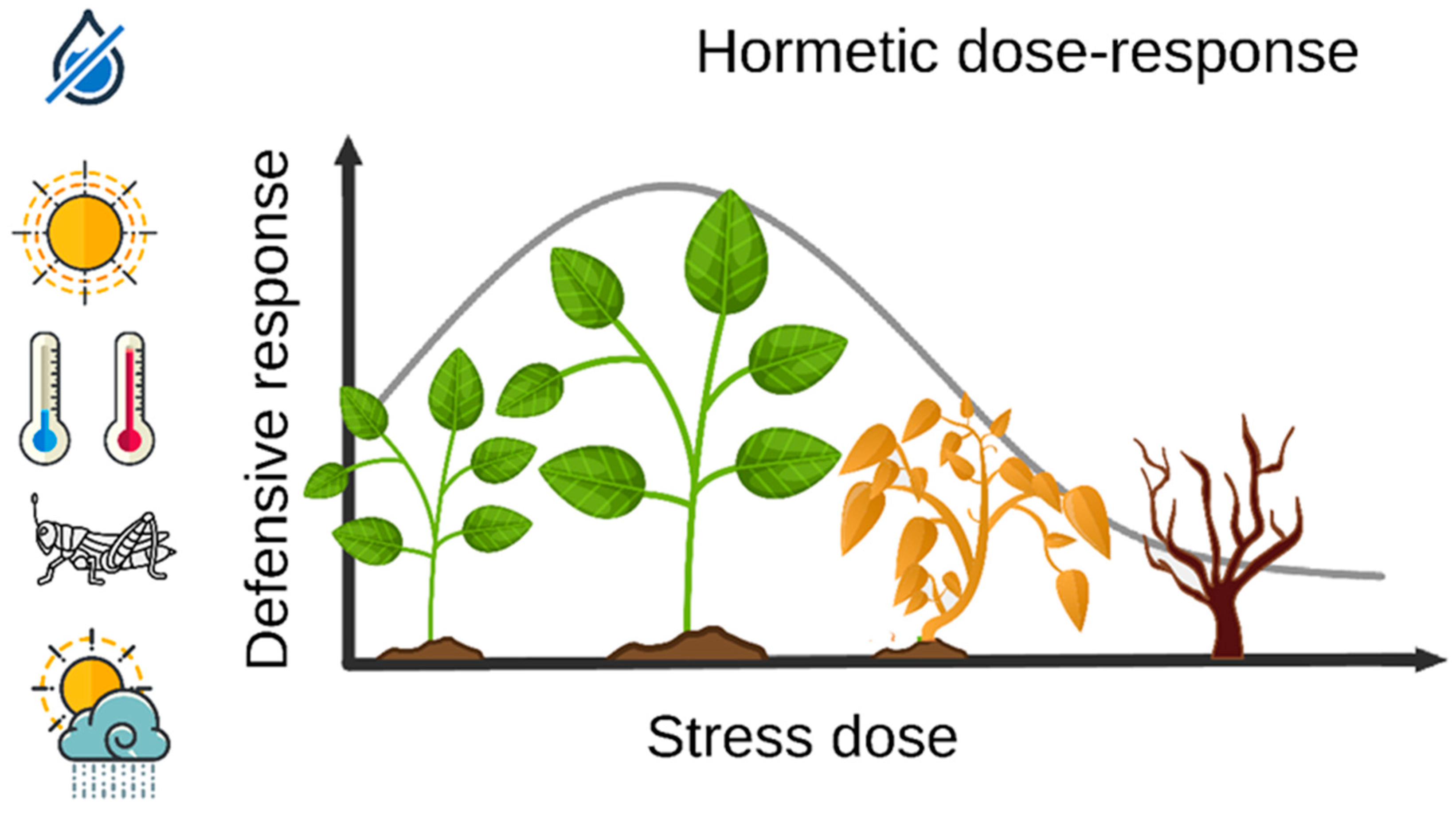
2. Hormesis and Plant Stress Science
3. Data in Plant Hormesis Research
3.1. Genomics and Transcriptomics
3.2. Proteomics
3.3. Metabolomics
3.4. Phenomics
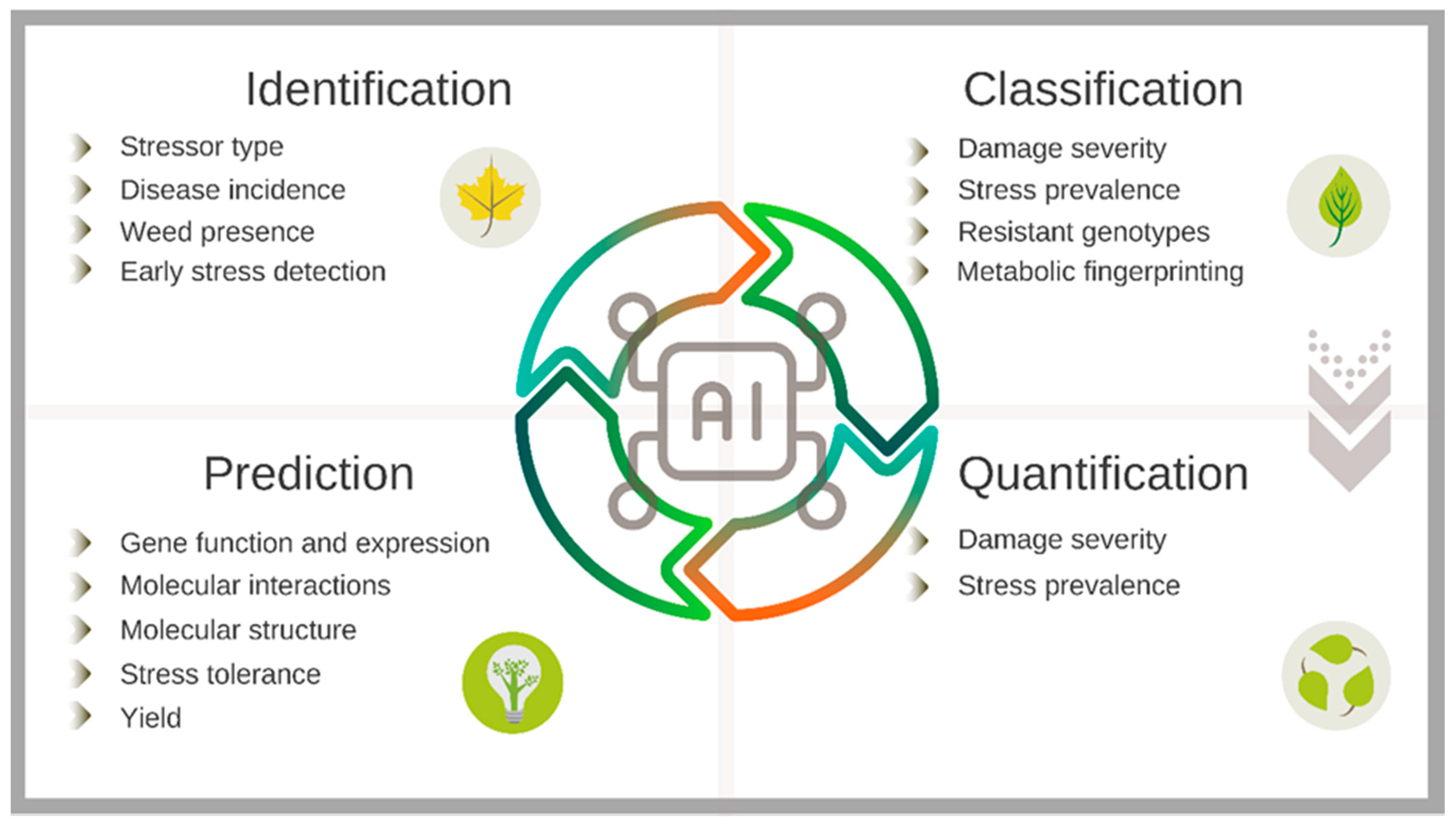
4. Artificial Intelligence Applications in Plant Stress Science
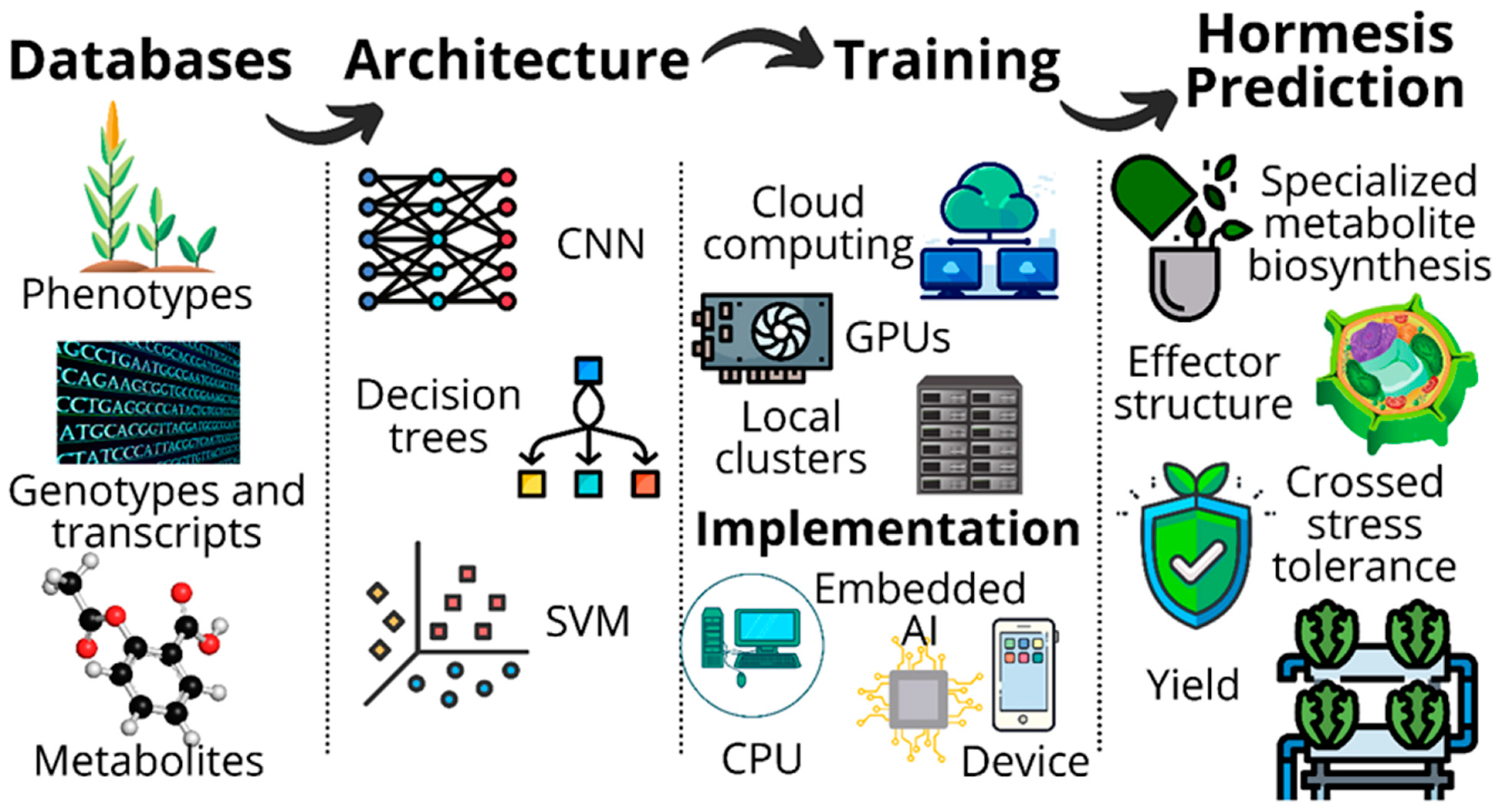
Deep Learning Platforms and Potential Applications for Plant Hormesis Management
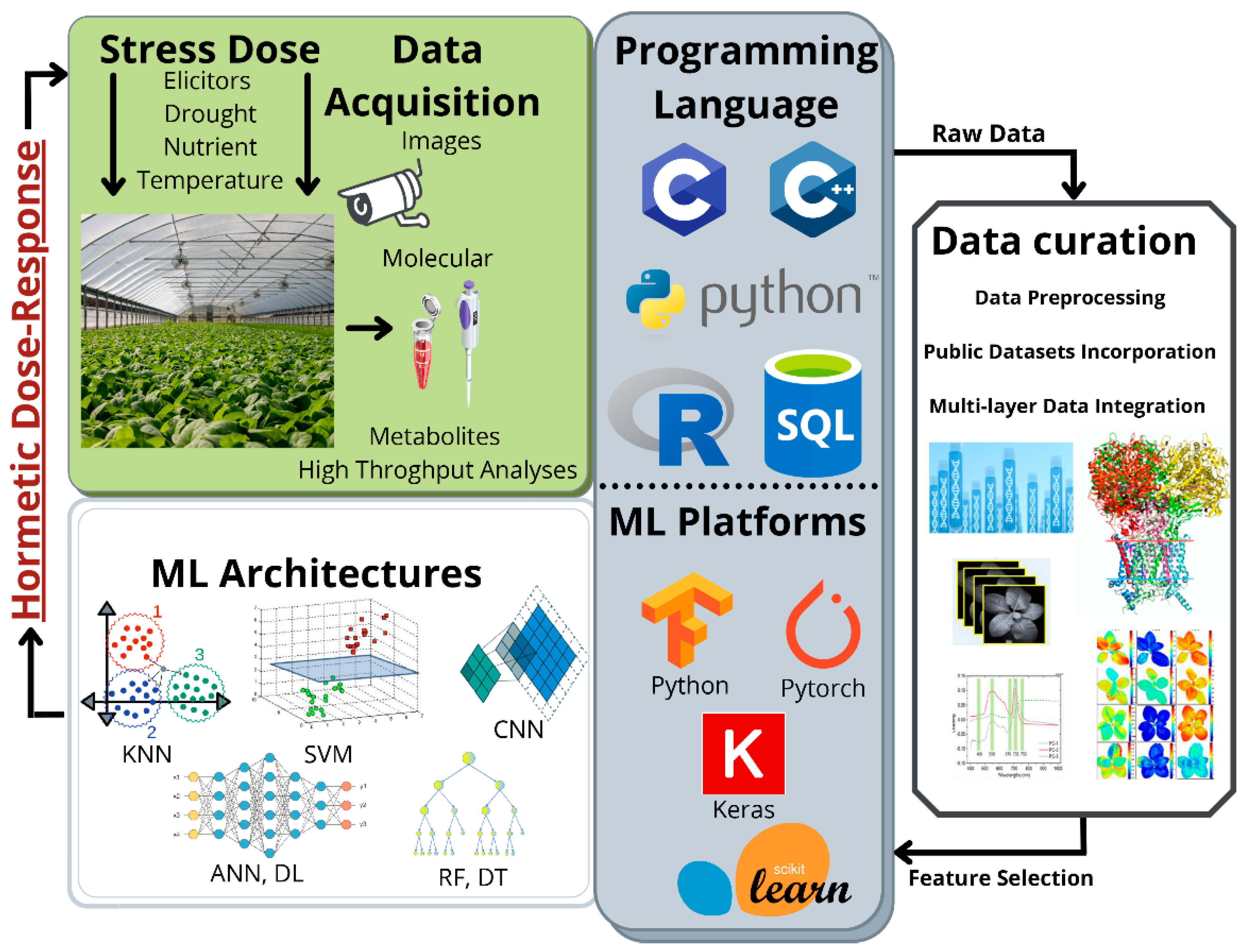
5. Limitations, Challenges, and Future Outlook
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jansen, M.A.; Potters, G. Stress: The Way of Life. In Plant Stress Physiology, 2nd ed.; CABI: London, UK, 2017; pp. ix–xiv. [Google Scholar]
- O’Brien, P.; Kral-O’Brien, K.; Hatfield, J.L. Agronomic Approach to Understanding Climate Change and Food Security. Agron. J. 2021, 113, 4616–4626. [Google Scholar] [CrossRef]
- Vázquez-Hernández, M.; Parola-Contreras, I.; Montoya-Gómez, L.; Torres-Pacheco, I.; Schwarz, D.; Guevara-González, R. Eustressors: Chemical and Physical Stress Factors Used to Enhance Vegetables Production. Sci. Hortic. 2019, 250, 223–229. [Google Scholar] [CrossRef]
- Bienertova-Vasku, J.; Lenart, P.; Scheringer, M. Eustress and Distress: Neither Good nor Bad, but Rather the Same? BioEssays 2020, 42, 1900238. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, V. Less Can Be More: The Hormesis Theory of Stress Adaptation in the Global Biosphere and Its Implications. Biomedicines 2021, 9, 293. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E.; Kitao, M.; Calabrese, E.J. Hormesis: Highly Generalizable and beyond Laboratory. Trends Plant Sci. 2020, 25, 1076–1086. [Google Scholar] [CrossRef]
- Agathokleous, E.; Kitao, M.; Calabrese, E.J. Hormesis: A Compelling Platform for Sophisticated Plant Science. Trends Plant Sci. 2019, 24, 318–327. [Google Scholar] [CrossRef]
- Vargas-Hernandez, M.; Macias-Bobadilla, I.; Guevara-Gonzalez, R.G.; Romero-Gomez, S.d.J.; Rico-Garcia, E.; Ocampo-Velazquez, R.V.; Alvarez-Arquieta, L.d.L.; Torres-Pacheco, I. Plant Hormesis Management with Biostimulants of Biotic Origin in Agriculture. Front. Plant Sci. 2017, 8, 1762. [Google Scholar] [CrossRef]
- Aguirre-Becerra, H.; Vazquez-Hernandez, M.C.; Saenz de la, O.D.; Alvarado-Mariana, A.; Guevara-Gonzalez, R.G.; Garcia-Trejo, J.F.; Feregrino-Perez, A.A. Role of Stress and Defense in Plant Secondary Metabolites Production. In Bioactive Natural Products for Pharmaceutical Applications; Springer: Berlin/Heidelberg, Germany, 2021; pp. 151–195. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C. Enhancing Quality of Fresh Vegetables through Salinity Eustress and Biofortification Applications Facilitated by Soilless Cultivation. Front. Plant Sci. 2018, 9, 1254. [Google Scholar] [CrossRef]
- Erofeeva, E.A. Plant Hormesis and Shelford’s Tolerance Law Curve. J. Res. 2021, 32, 1789–1802. [Google Scholar] [CrossRef]
- Jalal, A.; de Oliveira Junior, J.C.; Ribeiro, J.S.; Fernandes, G.C.; Mariano, G.G.; Trindade, V.D.R.; Dos Reis, A.R. Hormesis in Plants: Physiological and Biochemical Responses. Ecotoxicol. Environ. Saf. 2021, 207, 111225. [Google Scholar] [CrossRef]
- Agathokleous, E.; Barceló, D.; Calabrese, E.J. US EPA: Is There Room to Open a New Window for Evaluating Potential Sub-Threshold Effects and Ecological Risks? Environ. Pollut. 2021, 284, 117372. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E.; Calabrese, E.J. Hormesis: The Dose Response for the 21st Century: The Future Has Arrived. Toxicology 2019, 425, 152249. [Google Scholar] [CrossRef] [PubMed]
- Mazhar, S.; Ditta, A.; Bulgariu, L.; Ahmad, I.; Ahmed, M.; Nadiri, A.A. Sequential treatment of paper and pulp industrial wastewater: Prediction of water quality parameters by Mamdani Fuzzy Logic model and phytotoxicity assessment. Chemosphere 2019, 227, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, Z.; Arshad, M.; Crowley, D.; Khoshnevisan, B.; Yousefi, M.; Imran, M.; Hussain, S. Comparative efficacy of ANN and ANFIS models in estimating biosurfactant production produced by Klebseilla sp. FKOD36. Stoch. Environ. Res. Risk Assess. 2016, 30, 353–363. [Google Scholar] [CrossRef]
- Soltis, P.S.; Nelson, G.; Zare, A.; Meineke, E.K. Plants Meet Machines: Prospects in Machine Learning for Plant Biology. Appl. Plant Sci. 2020, 8, e11371. [Google Scholar] [CrossRef]
- van Dijk, A.D.J.; Kootstra, G.; Kruijer, W.; de Ridder, D. Machine Learning in Plant Science and Plant Breeding. Iscience 2021, 24, 101890. [Google Scholar] [CrossRef]
- Mahood, E.H.; Kruse, L.H.; Moghe, G.D. Machine Learning: A Powerful Tool for Gene Function Prediction in Plants. Appl. Plant Sci. 2020, 8, e11376. [Google Scholar] [CrossRef]
- Benos, L.; Tagarakis, A.C.; Dolias, G.; Berruto, R.; Kateris, D.; Bochtis, D. Machine Learning in Agriculture: A Comprehensive Updated Review. Sensors 2021, 21, 3758. [Google Scholar] [CrossRef]
- Gao, Z.; Luo, Z.; Zhang, W.; Lv, Z.; Xu, Y. Deep Learning Application in Plant Stress Imaging: A Review. AgriEngineering 2020, 2, 430–446. [Google Scholar] [CrossRef]
- Singh, A.K.; Ganapathysubramanian, B.; Sarkar, S.; Singh, A. Deep Learning for Plant Stress Phenotyping: Trends and Future Perspectives. Trends Plant Sci. 2018, 23, 883–898. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Ganapathysubramanian, B.; Singh, A.K.; Sarkar, S. Machine Learning for High-Throughput Stress Phenotyping in Plants. Trends Plant Sci. E 2016, 21, 110–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, E.J.; Agathokleous, E. Hormesis: Transforming Disciplines That Rely on the Dose Response. IUBMB Life 2022, 74, 8–23. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Historical Foundations of Hormesis. Homeopathy 2015, 104, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. Hormesis: Path and Progression to Significance. Int. J. Mol. Sci. 2018, 19, 2871. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Calabrese, E.J.; Lin, Z.; Lian, B.; Zhang, X. Similarities between the Yin/Yang Doctrine and Hormesis in Toxicology and Pharmacology. Trends Pharm. Sci. 2020, 41, 544–556. [Google Scholar] [CrossRef]
- Agathokleous, E.; Calabrese, E.J. Hormesis Can Enhance Agricultural Sustainability in a Changing World. Glob. Food Secur. 2019, 20, 150–155. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Mattson, M.P. Hormesis Provides a Generalized Quantitative Estimate of Biological Plasticity. J. Cell Commun. Signal. 2011, 5, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, E.J.; Blain, R.B. Hormesis and Plant Biology. Environ. Pollut. 2009, 157, 42–48. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormetic Mechanisms. Crit. Rev. Toxicol. 2013, 43, 580–606. [Google Scholar] [CrossRef]
- Mitchell, C.; Brennan, R.M.; Graham, J.; Karley, A.J. Plant Defense against Herbivorous Pests: Exploiting Resistance and Tolerance Traits for Sustainable Crop Protection. Front. Plant Sci. 2016, 7, 1132. [Google Scholar] [CrossRef] [Green Version]
- Lamers, J.; Van Der Meer, T.; Testerink, C. How Plants Sense and Respond to Stressful Environments. Plant Physiol. 2020, 182, 1624–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jez, J.M.; Topp, C.N.; Silva, G.; Tomlinson, J.; Onkokesung, N.; Sommer, S.; Mrisho, L.; Legg, J.; Adams, I.P.; Gutierrez-Vazquez, Y. Plant Pest Surveillance: From Satellites to Molecules. Emerg. Top. Life Sci. 2021, 5, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Ngou, B.P.M.; Jones, J.D.; Ding, P. Plant Immune Networks. Trends Plant Sci. 2021, 27, 255–273. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Ngou, B.P.M.; Ding, P.; Xin, X.-F. PTI-ETI Crosstalk: An Integrative View of Plant Immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef] [PubMed]
- Zarattini, M.; Farjad, M.; Launay, A.; Cannella, D.; Soulié, M.-C.; Bernacchia, G.; Fagard, M. Every Cloud Has a Silver Lining: How Abiotic Stresses Affect Gene Expression in Plant-Pathogen Interactions. J. Exp. Bot. 2021, 72, 1020–1033. [Google Scholar] [CrossRef]
- Aerts, N.; Pereira Mendes, M.; Van Wees, S.C. Multiple Levels of Crosstalk in Hormone Networks Regulating Plant Defense. Plant J. 2021, 105, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Loo, E.P. Plant Immunity in Signal Integration between Biotic and Abiotic Stress Responses. New Phytol. 2020, 225, 87–104. [Google Scholar] [CrossRef] [Green Version]
- Jamil, I.N.; Remali, J.; Azizan, K.A.; Nor Muhammad, N.A.; Arita, M.; Goh, H.-H.; Aizat, W.M. Systematic Multi-Omics Integration (MOI) Approach in Plant Systems Biology. Front. Plant Sci. 2020, 11, 944. [Google Scholar] [CrossRef]
- Naithani, S.; Tripathi, J.N.; Kumar, D. Systems Biology Approach for Improving and Sustaining Agriculture. Curr. Plant Biol. 2021, 28, 100230. [Google Scholar] [CrossRef]
- Argueso, C.T.; Assmann, S.M.; Birnbaum, K.D.; Chen, S.; Dinneny, J.R.; Doherty, C.J.; Eveland, A.L.; Friesner, J.; Greenlee, V.R.; Law, J.A. Directions for Research and Training in Plant Omics: Big Questions and Big Data. Plant Direct 2019, 3, e00133. [Google Scholar] [CrossRef] [Green Version]
- Libault, M.; Pingault, L.; Zogli, P.; Schiefelbein, J. Plant Systems Biology at the Single-Cell Level. Trends Plant Sci. 2017, 22, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Mir, R.R.; Reynolds, M.; Pinto, F.; Khan, M.A.; Bhat, M.A. High-throughput phenotyping for crop improvement in the genomics era. Plant Sci. 2019, 282, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Purugganan, M.D.; Jackson, S.A. Advancing Crop Genomics from Lab to Field. Nat. Genet. 2021, 53, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Cui, J.; Wang, L.; Teng, N.; Zhang, S.; Lam, H.-M.; Zhu, Y.; Xiao, S.; Ke, W.; Lin, J. Genome-Wide DNA Mutations in Arabidopsis Plants after Multigenerational Exposure to High Temperatures. Genome Biol. 2021, 22, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Zhu, G.; Li, Y.; Li, W.; Fu, J.; Niu, E.; Li, L.; Zhang, D.; Guo, W. Genome-Wide Association Studies Reveal Genetic Variation and Candidate Genes of Drought Stress Related Traits in Cotton (Gossypium Hirsutum L.). Front. Plant Sci. 2018, 9, 1276. [Google Scholar] [CrossRef]
- Hu, H.; Scheben, A.; Verpaalen, B.; Tirnaz, S.; Bayer, P.E.; Hodel, R.G.; Batley, J.; Soltis, D.E.; Soltis, P.S.; Edwards, D. Amborella Gene Presence/Absence Variation Is Associated with Abiotic Stress Responses That May Contribute to Environmental Adaptation. New Phytol. 2022, 233, 1548–1555. [Google Scholar] [CrossRef]
- Mo, F.; Wang, M.; Li, H.; Li, Y.; Li, Z.; Deng, N.; Chai, R.; Wang, H. Biological Effects of Silver Ions to Trifolium Pratense L. Revealed by Analysis of Biochemical Indexes, Morphological Alteration and Genetic Damage Possibility with Special Reference to Hormesis. Environ. Exp. Bot. 2021, 186, 104458. [Google Scholar] [CrossRef]
- Sertse, D.; You, F.M.; Ravichandran, S.; Soto-Cerda, B.J.; Duguid, S.; Cloutier, S. Loci Harboring Genes with Important Role in Drought and Related Abiotic Stress Responses in Flax Revealed by Multiple GWAS Models. Theor. Appl. Genet. 2021, 134, 191–212. [Google Scholar] [CrossRef]
- Luo, Z.; Szczepanek, A.; Abdel-Haleem, H. Genome-Wide Association Study (GWAS) Analysis of Camelina Seedling Germination under Salt Stress Condition. Agronomy 2020, 10, 1444. [Google Scholar] [CrossRef]
- Xiao, Q.; Bai, X.; Zhang, C.; He, Y. Advanced High-Throughput Plant Phenotyping Techniques for Genome-Wide Association Studies: A Review. J. Adv. Res. 2022, 35, 215–230. [Google Scholar] [CrossRef]
- Crossa, J.; Fritsche-Neto, R.; Montesinos-Lopez, O.A.; Costa-Neto, G.; Dreisigacker, S.; Montesinos-Lopez, A.; Bentley, A.R. The Modern Plant Breeding Triangle: Optimizing the Use of Genomics, Phenomics, and Enviromics Data. Front. Plant Sci. 2021, 12, 651480. [Google Scholar] [CrossRef] [PubMed]
- Fugeray-Scarbel, A.; Bastien, C.; Dupont-Nivet, M.; Lemarié, S. R2D2 Consortium Why and How to Switch to Genomic Selection: Lessons from Plant and Animal Breeding Experience. Front. Genet. 2021, 12, 1185. [Google Scholar]
- Zhan, X.; Lu, Y.; Zhu, J.; Botella, J.R. Genome Editing for Plant Research and Crop Improvement. J. Integr. Plant Biol. 2021, 63, 3–33. [Google Scholar] [CrossRef] [PubMed]
- Zappia, L.; Theis, F.J. Over 1000 tools reveal trends in the single-cell RNA-seq analysis landscape. Genome Biol 2021, 22, 1–8. [Google Scholar] [CrossRef]
- Imadi, S.R.; Kazi, A.G.; Ahanger, M.A.; Gucel, S.; Ahmad, P. Plant Transcriptomics and Responses to Environmental Stress: An Overview. J. Genet. 2015, 94, 525–537. [Google Scholar] [CrossRef]
- Gorbatova, I.V.; Kazakova, E.A.; Podlutskii, M.S.; Pishenin, I.A.; Bondarenko, V.S.; Dontsova, A.A.; Dontsov, D.P.; Snegirev, A.S.; Makarenko, E.S.; Bitarishvili, S.V. Studying Gene Expression in Irradiated Barley Cultivars: PM19L-like and CML31-like Expression as Possible Determinants of Radiation Hormesis Effect. Agronomy 2020, 10, 1837. [Google Scholar] [CrossRef]
- Duarte-Sierra, A.; Nadeau, F.; Angers, P.; Michaud, D.; Arul, J. UV-C Hormesis in Broccoli Florets: Preservation, Phyto-Compounds and Gene Expression. Postharvest Biol. Technol. 2019, 157, 110965. [Google Scholar] [CrossRef]
- Scott, G.; Dickinson, M.; Shama, G.; Rupar, M. A Comparison of the Molecular Mechanisms Underpinning High-Intensity, Pulsed Polychromatic Light and Low-Intensity UV-C Hormesis in Tomato Fruit. Postharvest Biol. Technol. 2018, 137, 46–55. [Google Scholar] [CrossRef]
- Volkova, P.Y.; Duarte, G.T.; Soubigou-Taconnat, L.; Kazakova, E.A.; Pateyron, S.; Bondarenko, V.S.; Bitarishvili, S.V.; Makarenko, E.S.; Churyukin, R.S.; Lychenkova, M.A. Early Response of Barley Embryos to Low-and High-dose Gamma Irradiation of Seeds Triggers Changes in the Transcriptional Profile and an Increase in Hydrogen Peroxide Content in Seedlings. J. Agron. Crop Sci. 2020, 206, 277–295. [Google Scholar] [CrossRef]
- Guo, J.; Ma, Z.; Peng, J.; Mo, J.; Li, Q.; Guo, J.; Yang, F. Transcriptomic Analysis of Raphidocelis Subcapitata Exposed to Erythromycin: The Role of DNA Replication in Hormesis and Growth Inhibition. J. Hazard. Mater. 2021, 402, 123512. [Google Scholar] [CrossRef]
- He, Y.; Wang, Y.; Hu, Y.; Chen, W.; Yan, Z. Superconducting Electrode Capacitor Based on Double-Sided YBCO Thin Film for Wireless Power Transfer Applications. Supercond. Sci. Technol. 2018, 32, 015010. [Google Scholar] [CrossRef]
- Arisha, M.H.; Ahmad, M.Q.; Tang, W.; Liu, Y.; Yan, H.; Kou, M.; Wang, X.; Zhang, Y.; Li, Q. RNA-Sequencing Analysis Revealed Genes Associated Drought Stress Responses of Different Durations in Hexaploid Sweet Potato. Sci. Rep. 2020, 10, 12573. [Google Scholar] [CrossRef]
- García-Gómez, M.L.; Castillo-Jiménez, A.; Martínez-García, J.C.; Álvarez-Buylla, E.R. Multi-Level Gene Regulatory Network Models to Understand Complex Mechanisms Underlying Plant Development. Curr. Opin. Plant Biol. 2020, 57, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.; Ahmad, J.S.; Clark, N.M.; Williams, C.M.; Sozzani, R. Computational Prediction of Gene Regulatory Networks in Plant Growth and Development. Curr. Opin. Plant Biol. 2019, 47, 96–105. [Google Scholar] [CrossRef]
- Wang, J.; Chen, B.; Ali, S.; Zhang, T.; Wang, Y.; Zhang, H.; Wang, L.; Zhang, Y.; Xie, L.; Jiang, T. Epigenetic Modification Associated with Climate Regulates Betulin Biosynthesis in Birch. J. Res. 2021, 1–15. [Google Scholar] [CrossRef]
- Qian, Y.; Huang, S.C. Improving Plant Gene Regulatory Network Inference by Integrative Analysis of Multi-Omics and High Resolution Data Sets. Curr. Opin. Syst. Biol. 2020, 22, 8–15. [Google Scholar] [CrossRef]
- Smith-Sonneborn, J. The Role of the ”Stress Protein Response” in Hormesis. In Biological Effects of Low Level Exposures to Chemicals and Radiation; CRC Press: Boca Raton, FL, USA, 2017; pp. 41–52. ISBN 1-315-15028-X. [Google Scholar]
- Koussounadis, A.; Langdon, S.P.; Um, I.H.; Harrison, D.J.; Smith, V.A. Relationship between Differentially Expressed MRNA and MRNA-Protein Correlations in a Xenograft Model System. Sci. Rep. 2015, 5, 10775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahoo, J.P.; Behera, L.; Sharma, S.S.; Praveena, J.; Nayak, S.K.; Samal, K.C. Omics Studies and Systems Biology Perspective towards Abiotic Stress Response in Plants. Am. J. Plant Sci. 2020, 11, 2172. [Google Scholar] [CrossRef]
- Buccitelli, C.; Selbach, M. MRNAs, Proteins and the Emerging Principles of Gene Expression Control. Nat. Rev. Genet. 2020, 21, 630–644. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Urban, M.O.; Prášil, I.T.; Renaut, J. Plant Abiotic Stress Proteomics: The Major Factors Determining Alterations in Cellular Proteome. Front. Plant Sci. 2018, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Mehmood, S.S.; Lu, G.; Luo, D.; Hussain, M.A.; Raza, A.; Zafar, Z.; Zhang, X.; Cheng, Y.; Zou, X.; Lv, Y. Integrated Analysis of Transcriptomics and Proteomics Provides Insights into the Molecular Regulation of Cold Response in Brassica Napus. Environ. Exp. Bot. 2021, 187, 104480. [Google Scholar] [CrossRef]
- Frukh, A.; Siddiqi, T.O.; Khan, M.I.R.; Ahmad, A. Modulation in Growth, Biochemical Attributes and Proteome Profile of Rice Cultivars under Salt Stress. Plant Physiol. Biochem. 2020, 146, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Chawade, A.; Alexandersson, E.; Bengtsson, T.; Andreasson, E.; Levander, F. Targeted Proteomics Approach for Precision Plant Breeding. J. Proteome Res. 2016, 15, 638–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Obaidi, J.R. Proteoinformatics and Agricultural Biotechnology Research: Applications and Challenges. In Essentials of Bioinformatics; Springer: Berlin/Heidelberg, Germany, 2019; Volume III, pp. 1–27. [Google Scholar]
- Komatsu, S. Plant Proteomic Research 2.0: Trends and Perspectives. Int. J. Mol. Sci. 2019, 20, 2495. [Google Scholar] [CrossRef] [Green Version]
- Jorrin-Novo, J.V. What Is New in (Plant) Proteomics Methods and Protocols: The 2015–2019 Quinquennium. In Plant Proteomics; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–10. [Google Scholar] [CrossRef]
- Jan, R.; Asaf, S.; Numan, M.; Kim, K.-M. Plant Secondary Metabolite Biosynthesis and Transcriptional Regulation in Response to Biotic and Abiotic Stress Conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Kosmacz, M.; Sokołowska, E.M.; Bouzaa, S.; Skirycz, A. Towards a Functional Understanding of the Plant Metabolome. Curr. Opin. Plant Biol. 2020, 55, 47–51. [Google Scholar] [CrossRef]
- Weng, J.-K.; Lynch, J.H.; Matos, J.O.; Dudareva, N. Adaptive Mechanisms of Plant Specialized Metabolism Connecting Chemistry to Function. Nat. Chem. Biol. 2021, 17, 1037–1045. [Google Scholar] [CrossRef]
- Rinschen, M.M.; Ivanisevic, J.; Giera, M.; Siuzdak, G. Identification of Bioactive Metabolites Using Activity Metabolomics. Nat. Rev. Mol. Cell Biol. 2019, 20, 353–367. [Google Scholar] [CrossRef]
- Verpoorte, R.; Choi, Y.H.; Kim, H.K. Metabolomics: Will It Stay? Phytochem. Anal. PCA 2010, 21, 2–3. [Google Scholar] [CrossRef]
- Lautie, E.; Russo, O.; Ducrot, P.; Boutin, J.A. Unraveling Plant Natural Chemical Diversity for Drug Discovery Purposes. Front. Pharm. 2020, 11, 397. [Google Scholar] [CrossRef]
- Sharma, D.; Kumar, S.; Kumar, V.; Thakur, A. Comprehensive Review on Nutraceutical Significance of Phytochemicals as Functional Food Ingredients for Human Health Management. J. Pharm. Phytochem. 2019, 8, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Pishenin, I.; Gorbatova, I.; Kazakova, E.; Podobed, M.; Mitsenyk, A.; Shesterikova, E.; Dontsova, A.; Dontsov, D.; Volkova, P. Free Amino Acids and Methylglyoxal as Players in the Radiation Hormesis Effect after Low-Dose γ-Irradiation of Barley Seeds. Agriculture 2021, 11, 918. [Google Scholar] [CrossRef]
- Mengdi, X.; Wenqing, C.; Haibo, D.; Xiaoqing, W.; Li, Y.; Yuchen, K.; Hui, S.; Lei, W. Cadmium-Induced Hormesis Effect in Medicinal Herbs Improves the Efficiency of Safe Utilization for Low Cadmium-Contaminated Farmland Soil. Ecotoxicol. Environ. Saf. 2021, 225, 112724. [Google Scholar] [CrossRef] [PubMed]
- Corrado, G.; Vitaglione, P.; Giordano, M.; Raimondi, G.; Napolitano, F.; Di Stasio, E.; Di Mola, I.; Mori, M.; Rouphael, Y. Phytochemical Responses to Salt Stress in Red and Green Baby Leaf Lettuce (Lactuca Sativa L.) Varieties Grown in a Floating Hydroponic Module. Separations 2021, 8, 175. [Google Scholar] [CrossRef]
- Alvarado, A.M.; Aguirre-Becerra, H.; Vázquez-Hernández, M.; Magaña-Lopez, E.; Parola-Contreras, I.; Caicedo-Lopez, L.H.; Contreras-Medina, L.M.; Garcia-Trejo, J.F.; Guevara-Gonzalez, R.G.; Feregrino-Perez, A.A. Influence of Elicitors and Eustressors on the Production of Plant Secondary Metabolites. In Natural Bio-Active Compounds; Springer: Berlin/Heidelberg, Germany, 2019; pp. 333–388. [Google Scholar] [CrossRef]
- Villate, A.; San Nicolas, M.; Gallastegi, M.; Aulas, P.-A.; Olivares, M.; Usobiaga, A.; Etxebarria, N.; Aizpurua-Olaizola, O. Metabolomics as a Prediction Tool for Plants Performance under Environmental Stress. Plant Sci. 2021, 303, 110789. [Google Scholar] [CrossRef]
- Miggiels, P.; Wouters, B.; van Westen, G.J.; Dubbelman, A.-C.; Hankemeier, T. Novel Technologies for Metabolomics: More for Less. TrAC Trends Anal. Chem. 2019, 120, 115323. [Google Scholar] [CrossRef]
- Hong, J.; Yang, L.; Zhang, D.; Shi, J. Plant Metabolomics: An Indispensable System Biology Tool for Plant Science. Int. J. Mol. Sci. 2016, 17, 767. [Google Scholar] [CrossRef]
- Pinu, F.R.; Beale, D.J.; Paten, A.M.; Kouremenos, K.; Swarup, S.; Schirra, H.J.; Wishart, D. Systems Biology and Multi-Omics Integration: Viewpoints from the Metabolomics Research Community. Metabolites 2019, 9, 76. [Google Scholar] [CrossRef] [Green Version]
- Pieruschka, R.; Schurr, U. Plant Phenotyping: Past, Present, and Future. Plant Phenomics 2019, 2019, 7507131. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity Induced Physiological and Biochemical Changes in Plants: An Omic Approach towards Salt Stress Tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Singh, V.; Sharma, N.; Singh, S. A Review of Imaging Techniques for Plant Disease Detection. Artif. Intell. Agric. 2020, 4, 229–242. [Google Scholar] [CrossRef]
- Zheng, C.; Abd-Elrahman, A.; Whitaker, V. Remote Sensing and Machine Learning in Crop Phenotyping and Management, with an Emphasis on Applications in Strawberry Farming. Remote Sens. 2021, 13, 531. [Google Scholar] [CrossRef]
- Li, D.; Li, C.; Yao, Y.; Li, M.; Liu, L. Modern Imaging Techniques in Plant Nutrition Analysis: A Review. Comput. Electron. Agric. 2020, 174, 105459. [Google Scholar] [CrossRef]
- Martinez, H.E.; de Souza, B.P.; Caixeta, E.T.; de Carvalho, F.P.; Clemente, J.M. Water Deficit Changes Nitrate Uptake and Expression of Some Nitrogen Related Genes in Coffee-Plants (Coffea Arabica L.). Sci. Hortic. 2020, 267, 109254. [Google Scholar] [CrossRef]
- Susič, N.; Žibrat, U.; Širca, S.; Strajnar, P.; Razinger, J.; Knapič, M.; Vončina, A.; Urek, G.; Stare, B.G. Discrimination between Abiotic and Biotic Drought Stress in Tomatoes Using Hyperspectral Imaging. Sens. Actuators B Chem. 2018, 273, 842–852. [Google Scholar] [CrossRef] [Green Version]
- Mayta, M.L.; Hajirezaei, M.-R.; Carrillo, N.; Lodeyro, A.F. Leaf Senescence: The Chloroplast Connection Comes of Age. Plants 2019, 8, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agathokleous, E.; Feng, Z.; Peñuelas, J. Chlorophyll Hormesis: Are Chlorophylls Major Components of Stress Biology in Higher Plants? Sci. Total Environ. 2020, 726, 138637. [Google Scholar] [CrossRef]
- Fenu, G.; Malloci, F.M. Forecasting Plant and Crop Disease: An Explorative Study on Current Algorithms. Big Data Cogn. Comput. 2021, 5, 2. [Google Scholar] [CrossRef]
- Pérez-Bueno, M.L.; Pineda, M.; Barón, M. Phenotyping Plant Responses to Biotic Stress by Chlorophyll Fluorescence Imaging. Front. Plant Sci. 2019, 1135. [Google Scholar] [CrossRef]
- Jakusch, P.; Kocsis, T.; Székely, I.K.; Hatvani, I.G. The Application of Magnetic Resonance Imaging (Mri) to the Examination of Plant Tissues and Water Barriers. Acta Biol. Hung. 2018, 69, 423–436. [Google Scholar] [CrossRef]
- Pineda, M.; Barón, M.; Pérez-Bueno, M.-L. Thermal Imaging for Plant Stress Detection and Phenotyping. Remote Sens. 2021, 13, 68. [Google Scholar] [CrossRef]
- Benavente, E.; García-Toledano, L.; Carrillo, J.; Quemada, M. Thermographic Imaging: Assessment of Drought and Heat Tolerance in Spanish Germplasm of Brachypodium Distachyon. Procedia Environ. Sci. 2013, 19, 262–266. [Google Scholar] [CrossRef] [Green Version]
- Lowe, A.; Harrison, N.; French, A.P. Hyperspectral Image Analysis Techniques for the Detection and Classification of the Early Onset of Plant Disease and Stress. Plant Methods 2017, 13, 80. [Google Scholar] [CrossRef] [PubMed]
- Großkinsky, D.K.; Svensgaard, J.; Christensen, S.; Roitsch, T. Plant Phenomics and the Need for Physiological Phenotyping across Scales to Narrow the Genotype-to-Phenotype Knowledge Gap. J. Exp. Bot. 2015, 66, 5429–5440. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Jones, S.; Ganapathysubramanian, B.; Sarkar, S.; Mueller, D.; Sandhu, K.; Nagasubramanian, K. Challenges and Opportunities in Machine-Augmented Plant Stress Phenotyping. Trends Plant Sci. 2021, 26, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Osama, K.; Mishra, B.N.; Somvanshi, P. Machine Learning Techniques in Plant Biology. In PlantOmics: The Omics of Plant Science; Springer: Berlin/Heidelberg, Germany, 2015; pp. 731–754. [Google Scholar]
- Chandel, N.S.; Chakraborty, S.K.; Rajwade, Y.A.; Dubey, K.; Tiwari, M.K.; Jat, D. Identifying Crop Water Stress Using Deep Learning Models. Neural Comput. Appl. 2021, 33, 5353–5367. [Google Scholar] [CrossRef]
- Yu, K.; Fang, S.; Zhao, Y. Heavy Metal Hg Stress Detection in Tobacco Plant Using Hyperspectral Sensing and Data-Driven Machine Learning Methods. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 245, 118917. [Google Scholar] [CrossRef]
- Rahman, C.R.; Arko, P.S.; Ali, M.E.; Khan, M.A.I.; Apon, S.H.; Nowrin, F.; Wasif, A. Identification and Recognition of Rice Diseases and Pests Using Convolutional Neural Networks. Biosyst. Eng. 2020, 194, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Blumenthal, J.; Megherbi, D.B.; Lussier, R. Unsupervised Machine Learning via Hidden Markov Models for Accurate Clustering of Plant Stress Levels Based on Imaged Chlorophyll Fluorescence Profiles & Their Rate of Change in Time. Comput. Electron. Agric. 2020, 174, 105064. [Google Scholar] [CrossRef]
- Esgario, J.G.; Krohling, R.A.; Ventura, J.A. Deep Learning for Classification and Severity Estimation of Coffee Leaf Biotic Stress. Comput. Electron. Agric. 2020, 169, 105162. [Google Scholar] [CrossRef] [Green Version]
- Das, B.; Manohara, K.; Mahajan, G.; Sahoo, R.N. Spectroscopy Based Novel Spectral Indices, PCA-and PLSR-Coupled Machine Learning Models for Salinity Stress Phenotyping of Rice. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 229, 117983. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, A.; Yang, C.; Marchetto, P.M. Ensemble Feature Selection for Plant Phenotyping: A Journey from Hyperspectral to Multispectral Imaging. IEEE Access 2018, 6, 56870–56884. [Google Scholar] [CrossRef]
- Barbedo, J.G.A. Plant Disease Identification from Individual Lesions and Spots Using Deep Learning. Biosyst. Eng. 2019, 180, 96–107. [Google Scholar] [CrossRef]
- Dao, P.D.; He, Y.; Proctor, C. Plant Drought Impact Detection Using Ultra-High Spatial Resolution Hyperspectral Images and Machine Learning. Int. J. Appl. Earth Obs. Geoinf. 2021, 102, 102364. [Google Scholar] [CrossRef]
- Karthickmanoj, R.; Sasilatha, T.; Padmapriya, J. Automated Machine Learning Based Plant Stress Detection System. Mater. Today Proc. 2021, 47, 1887–1891. [Google Scholar] [CrossRef]
- Ghosal, S.; Blystone, D.; Singh, A.K.; Ganapathysubramanian, B.; Singh, A.; Sarkar, S. An Explainable Deep Machine Vision Framework for Plant Stress Phenotyping. Proc. Natl. Acad. Sci. USA 2018, 115, 4613–4618. [Google Scholar] [CrossRef] [Green Version]
- Zahid, A.; Dashtipour, K.; Abbas, H.T.; Mabrouk, I.B.; Al-Hasan, M.; Ren, A.; Imran, M.A.; Alomainy, A.; Abbasi, Q.H. Machine Learning Enabled Identification and Real-Time Prediction of Living Plants’ Stress Using Terahertz Waves. Def. Technol. 2022; in press. [Google Scholar] [CrossRef]
- Niu, Y.; Han, W.; Zhang, H.; Zhang, L.; Chen, H. Estimating Fractional Vegetation Cover of Maize under Water Stress from UAV Multispectral Imagery Using Machine Learning Algorithms. Comput. Electron. Agric. 2021, 189, 106414. [Google Scholar] [CrossRef]
- Kang, D.; Ahn, H.; Lee, S.; Lee, C.-J.; Hur, J.; Jung, W.; Kim, S. Identifying Stress-Related Genes and Predicting Stress Types in Arabidopsis Using Logical Correlation Layer and CMCL Loss through Time-Series Data. In Proceedings of the 2018 IEEE International Conference on Bioinformatics and Biomedicine (BIBM), Madrid, Spain, 3–6 December 2018; IEEE: Piscataway, NJ, USA, 2018; pp. 399–404. [Google Scholar] [CrossRef]
- Azimi, S.; Kaur, T.; Gandhi, T.K. A Deep Learning Approach to Measure Stress Level in Plants Due to Nitrogen Deficiency. Measurement 2021, 173, 108650. [Google Scholar] [CrossRef]
- Fürtauer, L.; Pschenitschnigg, A.; Scharkosi, H.; Weckwerth, W.; Nägele, T. Combined Multivariate Analysis and Machine Learning Reveals a Predictive Module of Metabolic Stress Response in Arabidopsis Thaliana. Mol. Omics 2018, 14, 437–449. [Google Scholar] [CrossRef] [Green Version]
- Khanna, R.; Schmid, L.; Walter, A.; Nieto, J.; Siegwart, R.; Liebisch, F. A Spatio Temporal Spectral Framework for Plant Stress Phenotyping. Plant Methods 2019, 15, 13. [Google Scholar] [CrossRef] [Green Version]
- Naik, H.S.; Zhang, J.; Lofquist, A.; Assefa, T.; Sarkar, S.; Ackerman, D.; Singh, A.; Singh, A.K.; Ganapathysubramanian, B. A Real-Time Phenotyping Framework Using Machine Learning for Plant Stress Severity Rating in Soybean. Plant Methods 2017, 13, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, D.R.; Papa, J.P.; Saraiva, G.F.R.; Souza, G.M. Automatic Classification of Plant Electrophysiological Responses to Environmental Stimuli Using Machine Learning and Interval Arithmetic. Comput. Electron. Agric. 2018, 145, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Mondal, M.; Edida, M.; Sharma, N.; Lall, B.; Raju, D. Plants Stress Response Detection by Selecting Minimal Bands of Hyperspectral Images. In Proceedings of the 2019 9th International Conference on Advances in Computing and Communication (ICACC), Kochi, India, 6–8 November 2019; IEEE: Piscataway, NJ, USA, 2019; pp. 230–233. [Google Scholar] [CrossRef]
- Venal, M.C.A.; Fajardo, A.C.; Hernandez, A.A. Plant Stress Classification for Smart Agriculture Utilizing Convolutional Neural Network-Support Vector Machine. In Proceedings of the 2019 International Conference on ICT for Smart Society (ICISS), Bandung, Indonesia, 19–20 November 2019; IEEE: Piscataway, NJ, USA, 2019; Volume 7, pp. 1–5. [Google Scholar] [CrossRef]
- González-Camacho, J.M.; Crossa, J.; Pérez-Rodríguez, P.; Ornella, L.; Gianola, D. Genome-Enabled Prediction Using Probabilistic Neural Network Classifiers. BMC Genom. 2016, 17, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vakilian, A.K. Machine Learning Improves Our Knowledge about MiRNA Functions towards Plant Abiotic Stresses. Sci. Rep. 2020, 10, 3041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikha, M.; Kanika, A.; Rao, A.R.; Mallikarjuna, M.G.; Gupta, H.S.; Nepolean, T. Genomic Selection for Drought Tolerance Using Genome-Wide SNPs in Maize. Front. Plant Sci. 2017, 8, 550. [Google Scholar] [CrossRef] [PubMed]
- Montesinos-López, A.; Montesinos-López, O.A.; Gianola, D.; Crossa, J.; Hernández-Suárez, C.M. Multi-Environment Genomic Prediction of Plant Traits Using Deep Learners with Dense Architecture. G3 Genes Genomes Genet. 2018, 8, 3813–3828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, D.; Huet, S.; Gauffreteau, A.; Rincent, R.; Touzy, G.; Mini, A.; Jannink, J.-L.; Cormier, F.; Paux, E.; Lafarge, S. Whole-Genome Prediction of Reaction Norms to Environmental Stress in Bread Wheat (Triticum Aestivum L.) by Genomic Random Regression. Field Crops Res. 2018, 216, 32–41. [Google Scholar] [CrossRef]
- Silva, J.C.F.; Teixeira, R.M.; Silva, F.F.; Brommonschenkel, S.H.; Fontes, E.P. Machine Learning Approaches and Their Current Application in Plant Molecular Biology: A Systematic Review. Plant Sci. 2019, 284, 37–47. [Google Scholar] [CrossRef]
- Hiddar, H.; Rehman, S.; Lakew, B.; Verma, R.P.S.; Al-Jaboobi, M.; Moulakat, A.; Kehel, Z.; Filali-Maltouf, A.; Baum, M.; Amri, A. Assessment and Modeling Using Machine Learning of Resistance to Scald (Rhynchosporium Commune) in Two Specific Barley Genetic Resources Subsets. Sci. Rep. 2021, 11, 15967. [Google Scholar] [CrossRef]
- Song, Q.; Lee, J.; Akter, S.; Rogers, M.; Grene, R.; Li, S. Prediction of Condition-Specific Regulatory Genes Using Machine Learning. Nucleic Acids Res. 2020, 48, e62. [Google Scholar] [CrossRef] [Green Version]
- Shook, J.; Gangopadhyay, T.; Wu, L.; Ganapathysubramanian, B.; Sarkar, S.; Singh, A.K. Crop Yield Prediction Integrating Genotype and Weather Variables Using Deep Learning. PLoS ONE 2021, 16, e0252402. [Google Scholar] [CrossRef] [PubMed]
- Yasrab, R.; Zhang, J.; Smyth, P.; Pound, M.P. Predicting Plant Growth from Time-Series Data Using Deep Learning. Remote Sens. 2021, 13, 331. [Google Scholar] [CrossRef]
- García-Pérez, P.; Zhang, L.; Miras-Moreno, B.; Lozano-Milo, E.; Landin, M.; Lucini, L.; Gallego, P.P. The Combination of Untargeted Metabolomics and Machine Learning Predicts the Biosynthesis of Phenolic Compounds in Bryophyllum Medicinal Plants (Genus Kalanchoe). Plants 2021, 10, 2430. [Google Scholar] [CrossRef] [PubMed]
- Dargan, S.; Kumar, M.; Ayyagari, M.R.; Kumar, G. A Survey of Deep Learning and Its Applications: A New Paradigm to Machine Learning. Arch. Comput. Methods Eng. 2020, 27, 1071–1092. [Google Scholar] [CrossRef]
- Nicodeme, C. Build Confidence and Acceptance of AI-Based Decision Support Systems-Explainable and Liable AI. In Proceedings of the 2020 13th International Conference on Human System Interaction (HSI), Tokyo, Japan, 6–8 June 2020; IEEE: Piscataway, NJ, USA, 2020; pp. 20–23. [Google Scholar] [CrossRef]
- Pavlopoulou, A.; Karaca, E.; Balestrazzi, A.; Georgakilas, A.G. In Silico Phylogenetic and Structural Analyses of Plant Endogenous Danger Signaling Molecules upon Stress. Oxidative Med. Cell. Longev. 2019, 2019, 8683054. [Google Scholar] [CrossRef] [Green Version]
- Stokes, J.M.; Yang, K.; Swanson, K.; Jin, W.; Cubillos-Ruiz, A.; Donghia, N.M.; MacNair, C.R.; French, S.; Carfrae, L.A.; Bloom-Ackermann, Z. A Deep Learning Approach to Antibiotic Discovery. Cell 2020, 180, 688–702. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhou, M.; Zou, Q.; Xu, L. Machine Learning for Phytopathology: From the Molecular Scale towards the Network Scale. Brief. Bioinform. 2021, 22, bbab037. [Google Scholar] [CrossRef]
- Mayr, A.; Klambauer, G.; Unterthiner, T.; Hochreiter, S. DeepTox: Toxicity Prediction Using Deep Learning. Front. Environ. Sci. 2016, 3, 80. [Google Scholar] [CrossRef] [Green Version]
- Prilianti, K.R.; Setiyono, E.; Kelana, O.H.; Brotosudarmo, T.H.P. Deep Chemometrics for Nondestructive Photosynthetic Pigments Prediction Using Leaf Reflectance Spectra. Inf. Processing Agric. 2021, 8, 194–204. [Google Scholar] [CrossRef]
- Mu, C.; Yuan, Z.; Ouyang, X.; Sun, P.; Wang, B. Non-destructive Detection of Blueberry Skin Pigments and Intrinsic Fruit Qualities Based on Deep Learning. J. Sci. Food Agric. 2021, 101, 3165–3175. [Google Scholar] [CrossRef]
- Durmuş, H.; Güneş, E.O.; Kırcı, M. Disease Detection on the Leaves of the Tomato Plants by Using Deep Learning. In Proceedings of the 2017 6th International Conference on Agro-Geoinformatics, Fairfax, VA, USA, 7–10 August 2017; IEEE: Piscataway, NJ, USA, 2017; pp. 1–5. [Google Scholar] [CrossRef]
- Ferentinos, K.P. Deep Learning Models for Plant Disease Detection and Diagnosis. Comput. Electron. Agric. 2018, 145, 311–318. [Google Scholar] [CrossRef]
- Too, E.C.; Yujian, L.; Njuki, S.; Yingchun, L. A Comparative Study of Fine-Tuning Deep Learning Models for Plant Disease Identification. Comput. Electron. Agric. 2019, 161, 272–279. [Google Scholar] [CrossRef]
- Wang, G.; Sun, Y.; Wang, J. Automatic Image-Based Plant Disease Severity Estimation Using Deep Learning. Comput. Intell. Neurosci. 2017, 2017, 2917536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Zeng, R.; Pan, J.; Wang, C.C.; Liu, Y.-J. Plant Phenotyping by Deep-Learning-Based Planner for Multi-Robots. IEEE Robot. Autom. Lett. 2019, 4, 3113–3120. [Google Scholar] [CrossRef]
- Ma, W.; Qiu, Z.; Song, J.; Li, J.; Cheng, Q.; Zhai, J.; Ma, C. A Deep Convolutional Neural Network Approach for Predicting Phenotypes from Genotypes. Planta 2018, 248, 1307–1318. [Google Scholar] [CrossRef]
- Tausen, M.; Clausen, M.; Moeskjær, S.; Shihavuddin, A.; Dahl, A.B.; Janss, L.; Andersen, S.U. Greenotyper: Image-Based Plant Phenotyping Using Distributed Computing and Deep Learning. Front. Plant Sci. 2020, 11, 1181. [Google Scholar] [CrossRef]
- Khan, A.I.; Al-Badi, A. Open Source Machine Learning Frameworks for Industrial Internet of Things. Procedia Comput. Sci. 2020, 170, 571–577. [Google Scholar] [CrossRef]
- Bresilla, K.; Perulli, G.D.; Boini, A.; Morandi, B.; Corelli Grappadelli, L.; Manfrini, L. Single-Shot Convolution Neural Networks for Real-Time Fruit Detection within the Tree. Front. Plant Sci. 2019, 10, 611. [Google Scholar] [CrossRef] [Green Version]
- Thomas, G.; Balocco, S.; Mann, D.; Simundsson, A.; Khorasani, N. Intelligent Agricultural Machinery Using Deep Learning. IEEE Instrum. Meas. Mag. 2021, 24, 93–100. [Google Scholar] [CrossRef]
- Valencia-Hernandez, J.-A.; Solano-Alvarez, N.; Rico-Rodriguez, M.-A.; Rodriguez-Ontiveros, A.; Torres-Pacheco, I.; Rico-Garcia, E.; Guevara-Gonzalez, R.-G. Eustressic Dose of Cadmium in Soil Induces Defense Mechanisms and Protection Against Clavibacter Michiganensis in Tomato (Solanum Lycopersicum L.). J. Plant Growth Regul. 2022, 1–8. [Google Scholar] [CrossRef]
- Sáenz-de la, O.D.; Morales, L.O.; Strid, Å.; Torres-Pacheco, I.; Guevara-González, R.G. Ultraviolet-B Exposure and Exogenous Hydrogen Peroxide Application Lead to Cross-tolerance toward Drought in Nicotiana Tabacum L. Physiol. Plant. 2021, 173, 666–679. [Google Scholar] [CrossRef] [PubMed]
- Khaki, S.; Wang, L. Crop Yield Prediction Using Deep Neural Networks. Front. Plant Sci. 2019, 10, 621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeCun, Y.; Bengio, Y.; Hinton, G. Deep Learning. Nature 2015, 521, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Naser, M.Z.; Kodur, V.; Thai, H.T.; Hawileh, R.; Abdalla, J.; Degtyarev, V.V. StructuresNet and FireNet: Benchmarking databases and machine learning algorithms in structural and fire engineering domains. J. Build. Eng. 2021, 44, 102977. [Google Scholar] [CrossRef]
- Liang, Y.; Li, S.; Yan, C.; Li, M.; Jiang, C. Explaining the black-box model: A survey of local interpretation methods for deep neural networks. Neurocomputing 2021, 419, 168–182. [Google Scholar] [CrossRef]
- Nalepa, J.; Kawulok, M. Selecting training sets for support vector machines: A review. Artif. Intell. Rev. 2019, 52, 857–900. [Google Scholar] [CrossRef] [Green Version]
- Sarker, I.H. Machine Learning: Algorithms, Real-World Applications and Research Directions. SN Comput. Sci. 2021, 2, 160. [Google Scholar] [CrossRef]
- Xu, C.; Jackson, S.A. Machine learning and complex biological data. Genome Biol. 2019, 20, 76. [Google Scholar] [CrossRef]
- Harfouche, A.L.; Jacobson, D.A.; Kainer, D.; Romero, J.C.; Harfouche, A.H.; Mugnozza, G.S.; Moshelion, M.; Tuskan, G.A.; Keurentjes, J.J.; Altman, A. Accelerating Climate Resilient Plant Breeding by Applying Next-Generation Artificial Intelligence. Trends Biotechnol. 2019, 37, 1217–1235. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Artificial Intelligence Technique | Algorithms | (ICQP) Application | Datasets | Model Plant Reported | Stressor | Reference |
|---|---|---|---|---|---|---|
| Deep Learning (image) | Convolutional neural networks (CNN), AlexNet, GoogLeNet, and Inception V3 | Identification | 1200 images acquired by camera under stress and non-stress conditions | Maize (Zea mays), okra (Abelmoschus esculentus), and soybean (Glycine max) | Water stress | Chandel et al. (2020) [113] |
| Unsupervised Machine learning | Least squares discriminant analysis (PLS-DA) and least-squares support vector machine (LS-SVM) | Identification | Hyperspectral images of the canopy of tobacco plants | Tobacco | Heavy metal stress Hg | Yu et al. (2021) [114] |
| Deep Learning (image) | CNN | Identification | 1426 images of rice diseases and pests from paddy fields | Rice | Biotic stress | Rahman et al. (2020) [115] |
| Unsupervised Machine learning (video imaging) | Hidden Markov models (HMMs) | Identification and classification | Chlorophyll fluorescence (ChlF) digital profiles from GrowTech Inc. | Phaseolus vulgaris L. (Snap bean) | Stressor “level” groups (low, medium, and high stressed) and three stressor “type” categories (drought, nutrient, and chemical stress) | Blumenthal et al. (2020) [116] |
| Deep Learning (image) | CNN | Identification and Quantification | 1747 smartphones images of arabica coffee leaves. | Arabica coffee | Biotic stress; leaf miner, rust, brown leaf spot, and Cercospora leaf spot | Esgario et al. (2020) [117] |
| Supervised Machine Learning, Partial Least Square Regression, Principal Component Analysis, and combined models | K-nearest neighbors (KNN) | Identification and classification | Spectral signature of leaf samples obtained with a visible, near-infrared spectrometer | Rice | Salt stress | Das et al. (2020) [118] |
| Supervised Machine Learning | ReliefF, support vector machine (SVM), recursive feature elimination (RFE), and random forest (RF) | Identification and classification | Hyperspectral images from four wheat lines | Wheat | Salt stress | Moghimi et al. (2018) [119] |
| Deep Learning (image) | CNN | Identification and classification | 1575 images (smartphones, compact cameras, DSLR | Different plant specimens | Biotic stress | Arnal Barbedo (2019) [120] |
| Deep Learning | RF, SVM, multilayer perceptron (MLP) | Identification and classification | Hyperspectral images | Bromus inermis | Drought stress | Dao et al. (2021) [121] |
| Supervised Machine Learning | SVM | Identification and classification | RGB leave images from the Kaggle database | Brinjal leaves | Biotic stress | Karthickmanoj et al. (2021) [122] |
| Deep Learning (image) | Deep convolutional neural network (DCNN) | Identification, classification, and quantification | Collection of images of stressed and healthy soybean leaflets in the field | Soybean [Glycine max (L.) Merr.] | Bacterial blight (Pseudomonas savastanoi pv. glycinea), bacterial pustule (Xanthomonas axonopodis pv. glycines), sudden death syndrome (Fusarium virguliforme), septoria brown spot (Septoria glycines), frogeye leaf spot (Cercospora sojina), iron deficiency chlorosis, potassium deficiency, and herbicide injury | Ghosal et al. (2018) [123] |
| Supervised Machine Learning | RF, SVM, KNN | Classification and prediction | Real time terahertz time-domain spectroscopic data (THz-TDS) | Basil, coriander, parsley, baby-leaf, coffee, pea- | Water Stress | Zahid et al. (2022) [124] |
| Supervised Machine Learning | RF, artificial neural networks (ANN), and | Classification | Multispectral images | Maize | Water stress | Niu et al. (2021) [125] |
| Supervised Machine Learning | Confident multiple-choice learning | Identification and prediction | Gene expression time-series datasets | Arabidopsis thaliana | Heat, cold, salt, and drought | Kang et al. (2018) [126] |
| Deep Learning (image) | CNN | Classification | Images of Sorghum plant shoot from the Donald Danforth Plant Science Center. | Sorghum plants | Nitrogen deficiency | Azimi et al. (2021) [127] |
| Supervised Machine Learning | Decision tree (DT), SVM, and Naïve Bayes (NB) | Classification | Metabolite and protein content | Arabidopsis thaliana | Metabolic stress | Fürtauer et al. (2018) [128] |
| Supervised Machine Learning | SVM | Classification | Biweekly RGB, stereo and hyperspectral spatio-temporal images | Sugar beet plants | Abiotic stress conditions (drought and nitrogen deficiency) and one biotic stressor (weed) | Khanna et al. (2019) [129] |
| Supervised Machine Learning | Hierarchical models | Classification | 5916 RGB images (493 plots including Plant Introduction (PI) accessions in different time points) | Soybean (Glycine max (L.) Merr.) | Iron deficiency chlorosis | Naik et al. (2017) [130] |
| Supervised Machine Learning | ANN, CNN, optimum-path forest, KNN, and SVM | Classification | Electrical signal under cold, low light and osmotic stimuli. | Soybean plants | Cold, low light, and osmotic stimuli. | Pereira et al. (2018) [131] |
| Supervised Machine Learning | RF | Classification | Hyperspectral dataset acquired from the Indian Agricultural Research Institute (IARI) | Wheat | Water stress | Mondal et al. (2019) [132] |
| Deep Learning (image) | CNN, SVM | Classification | 65,184 labeled images from Github resources | Soybean | Biotic (fungal and bacterial diseases) and abiotic (nutrient deficiency and chemical injury) stresses | Venal et al. (2019) [133] |
| Supervised Machine Learning | MLP and probabilistic neural network (PNN) | Classification | 16 maize and 17 wheat genomic and phenotypic datasets with different trait-environment combinations | Maize and Wheat | Drought | González-Camacho et al. (2016) [134] |
| Supervised Machine Learning | Decision tree (DT), SVM, and NB | Prediction | miRNA concentration. | Arabidopsis thaliana plants | Drought, salinity, cold, and heat | Vakilian (2020) [135] |
| Supervised Machine Learning | Ridge regression, LASSO, elastic net, RF, reproducing kernel Hilbert space, Bayes A and Bayes B | Prediction | A set of 29,619 cured Single Nucleotide Polymorphisms, genotyped across a panel of 240 maize inbred lines | Maize | Drought stress | Shikha et al. (2017) [136] |
| Deep Learning | CNN | Prediction | Three maize and six wheat data sets. | Maize and wheat | Environmental stress | Montesinos-López et al. (2018) [137] |
| Supervised Machine Learning | Genomic random regression | Prediction | Complete genotypes, molecular markers, and phenotypic traits of stressed and control groups. | Wheat | Environmental stress | Ly et al. (2018) [138] |
| DL Architecture | Application | Hardware | Reference |
|---|---|---|---|
| Deep Neural Networks | Toxicity Prediction | Nvidia Tesla K40 | Mayr et al. (2016) [150] |
| Convolutional Neural Network | Photosynthetic pigments Prediction | CPU core i5 1.6 GHz, 8 GB DDR3 RAM, GPU not specified | Prilianti et al. (2020) [151] |
| Convolutional Neural Network | Pigments Prediction | Nvidia GTX 1020Ti, Intel Xeon W-2133, 32 GB | Mu et al. (2020) [152] |
| AlexNet and SqueezeNet | Plant Disease Detection | Nvidia Jetson TX1 | Durmus et al. (2017) [153] |
| Convolutional Neural Network | Plant Disease Detection | Nvidia GTX1080 | Ferentinos (2018) [154] |
| Convolutional Neural Network | Plant Disease Detection | Nvidia Tesla K40c | Too et al. (2019) [155] |
| Deconvolutional Neural Network | Plant Disease Detection | Nvidia GeForce Titan X, Intel Core I7 3.5 GHz | Wang et al. (2017) [156] |
| Point Completion Network | Plant Phenotyping | Nvidia Titan V. Xeon Gold 6146 3.20 GHz, 128 GB RAM | Wu et al. (2019) [157] |
| Deep Convolutional Neural Network | Predicting Phenotypes from Genotypes | Nvidia GeForce TITAN-XGPU | Ma et al. (2018) [158] |
| U-net | Phenotyping and Plant Growth | Nvidia Tela V100 | Tausen et al. (2020) [159] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rico-Chávez, A.K.; Franco, J.A.; Fernandez-Jaramillo, A.A.; Contreras-Medina, L.M.; Guevara-González, R.G.; Hernandez-Escobedo, Q. Machine Learning for Plant Stress Modeling: A Perspective towards Hormesis Management. Plants 2022, 11, 970. https://doi.org/10.3390/plants11070970
Rico-Chávez AK, Franco JA, Fernandez-Jaramillo AA, Contreras-Medina LM, Guevara-González RG, Hernandez-Escobedo Q. Machine Learning for Plant Stress Modeling: A Perspective towards Hormesis Management. Plants. 2022; 11(7):970. https://doi.org/10.3390/plants11070970
Chicago/Turabian StyleRico-Chávez, Amanda Kim, Jesus Alejandro Franco, Arturo Alfonso Fernandez-Jaramillo, Luis Miguel Contreras-Medina, Ramón Gerardo Guevara-González, and Quetzalcoatl Hernandez-Escobedo. 2022. "Machine Learning for Plant Stress Modeling: A Perspective towards Hormesis Management" Plants 11, no. 7: 970. https://doi.org/10.3390/plants11070970