

Draft Genome Sequence of Priestia sp. Strain TSO9, a Plant Growth-Promoting Bacterium Associated with Wheat (Triticum turgidum subsp. durum) in the Yaqui Valley, Mexico

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterium Culture Conditions
2.2. Metabolic Characterization
- Production of indole acetic acid (IAA) was assessed as mentioned by de los Santos et al. [23]. First, 1 mL (1 × 106 cells/mL) was inoculated in 10 mL of NB supplemented with 100 mg/L of tryptophan at 30 ± 2 °C for 5 days, in a rotary shaker at 120 rpm. After incubation, the production of IAA was determined by spectrophotometry assays according to Glickmann and Dessaux [24].
- Siderophore production. Chrome Azurosol S (CAS) agar was prepared from four solutions, which were sterilized separately before mixing. The culture medium was spot-inoculated in triplicate using 10 µL (1 × 106 cells/mL) and incubated for 7 days at 28 ± 2°C [27]. The presence of a yellow–orange halo around the inoculated colony was recorded as a positive result.
- Abiotic stress. Here, 1 × 106 colony-forming units (CFU) of strain TSO9 were spot-inoculated on Petri dishes containing NA as a culture medium and supplemented with (i) sodium chloride to determine saline stress and (ii) Polyethylene Glycol 6000 (10%, −0.84 mPa) to determine hydric stress; and (iii) the inoculum was incubated at a temperature of 43.5 °C for 3 days, to determine the thermal stress. The control treatment was conducted by spot-inoculating 1 × 105 CFU of strain TSO9 containing only NA and incubating it at 28 °C [22]. The growth of strain TSO9 under these conditions was recorded as tolerance to abiotic stress.
- Biocontrol. A dual confrontation assay against the wheat phytopathogen Bipolaris sorokiniana TPQ3 was carried out. A volume of 10 µL of 1 × 105 conidia/mL was spot-inoculated in the center of a Petri dish (8 cm in diameter) containing potato dextrose agar, and then 10 µL of 1 × 106 CFU of strain TSO9 was spot-inoculated at four equidistant points (2 cm of distance), in triplicate, around B. sorokiniana TPQ3, and the sample was incubated for 5 days at 28 °C [6]. The growth inhibition of the phytopathogen indicated biocontrol by the strain TSO9.
- Bacteria—Wheat plants interactions were assessed under a greenhouse assay. The growth promotion traits of strain TSO9 were analyzed through a greenhouse assay where fifteen wheat seeds (var. CIRNO C2008) per treatment were germinated on Petri dishes containing agar–agar (8 g L−1). Then, the wheat seedlings (7 days post-germination) were transplanted in pots containing 1.5 kg of non-sterilized soil and were inoculated; for this, the bacterial strain TSO9 was grown in 30 mL of sterile nutrient broth contained in a Falcon tube (50 mL) and incubated for 2 days at 28 °C and 180 rpm. Then, the bacterial culture was centrifuged at 3600 g for 10 min and the obtained pellet was washed twice and re-suspended in sterile distilled water. The optical density (630 nm) of strain TSO9 was adjusted to 0.5 (1 × 108 CFU mL−1). Thus, 5 mL (5 × 108 CFU) of this strain was inoculated on the wheat’s rhizosphere, and in the negative control, the cell suspension was replaced with 5 mL sterile distilled water. The greenhouse assay was carried out for 3 months, under the climatic conditions of the Yaqui Valley (13 h of darkness at 14 °C, 2 h of light at 18 °C, 7 h of light at 25 °C, and 2 h of light at 18 °C). Plant biometric parameters such as leaf number, stem diameter, stem height, root length, and plant dry weight were evaluated as described by Valenzuela-Aragon et al. [22]. Results were reported as mean values and percentages, calculated using the following formula: [(Treatment value − Control value)/Control value] × 100.
2.3. Genomic Analysis
2.4. Genome Annotation
2.5. Genome Mining
3. Results
3.1. Morphological and Metabolic Characterization
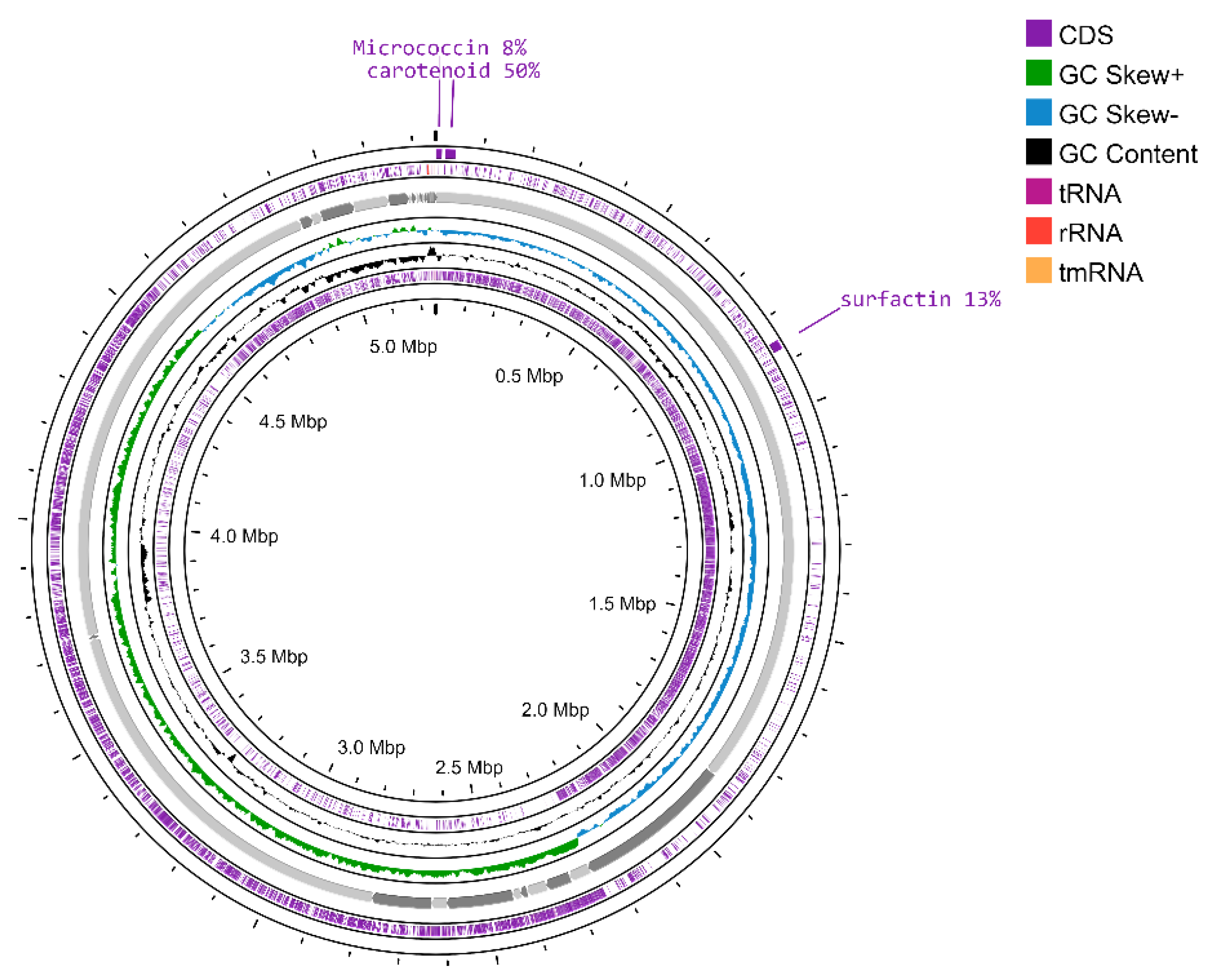
3.2. Genomic Analysis
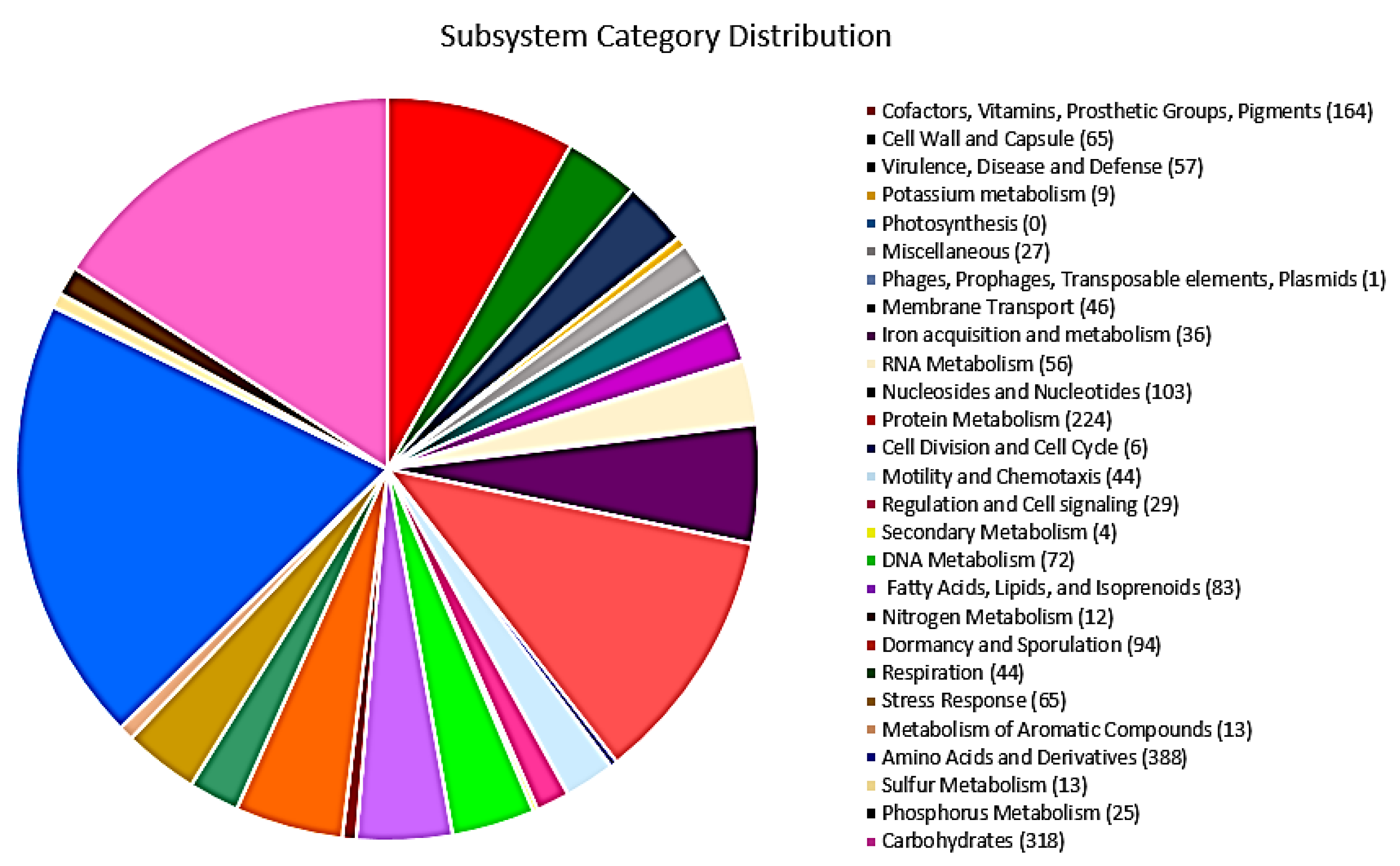
3.3. Genome Annotation
3.4. Genome Mining
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Omomowo, O.I.; Babalola, O.O. Constraints and Prospects of Improving Cowpea Productivity to Ensure Food, Nutritional Security and Environmental Sustainability. Front. Plant Sci. 2021, 12, 751731. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, A.M. Bipolaris Sorokiniana-Induced Black Point, Common Root Rot, and Spot Blotch Diseases of Wheat: A Review. Front. Cell. Infec. Micro. 2021, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- OECD-FAO. Chapter 3. Cereals. In OECD-FAO Agricultural Outlook; OECD Publishing: Paris, France; FAO: Rome, Italy, 2018; pp. 109–126. [Google Scholar] [CrossRef]
- Córdova-Albores, L.C.; Zelaya-Molina, L.X.; Ávila-Alistac, N.; Valenzuela-Ruíz, V.; Cortés-Martínez, N.E.; Parra-Cota, F.I.; Burgos-Canul, Y.Y.; Chávez-Díaz, I.F.; Fajardo-Franco, M.L.; De los Santos-Villalobos, S. Omics Sciences Potential on Bioprospecting of Biological Control Microbial Agents: The Case of the Mexican Agro-Biotechnology. Rev. Mex. Fitopatol. 2020, 39, 147–184. [Google Scholar] [CrossRef]
- Dhakal, A.; Adhikari, C.; Manandhar, D.; Bhattarai, S.; Shrestha, S. Effect of Abiotic Stress in Wheat: A Review. Rev. Food Agric. 2021, 2, 69–72. [Google Scholar] [CrossRef]
- Villa-Rodriguez, E.; Moreno-Ulloa, A.; Castro-Longoria, E.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Integrated Omics Approaches for Deciphering Antifungal Metabolites Produced by a Novel Bacillus Species, B. Cabrialesii TE3T, against the Spot Blotch Disease of Wheat (Triticum Turgidum L. Subsp. Durum). Microbiol. Res. 2021, 251, 126826. [Google Scholar] [CrossRef]
- Robles-Montoya, R.; Chaparro-Encinas, L.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Improving Biometric Traits of Wheat Seedlings with the Inoculation of a Native Consortium of Bacillus. Rev. Mex. Cienc. Agrícolas 2020, 11, 229–235. [Google Scholar] [CrossRef]
- Díaz-Rodríguez, A.M.; Salcedo Gastelum, L.A.; Félix Pablos, C.M.; Parra-Cota, F.I.; Santoyo, G.; Puente, M.L.; Bhattacharya, D.; Mukherjee, J.; de los Santos-Villalobos, S. The Current and Future Role of Microbial Culture Collections in Food Security Worldwide. Front. Sustain. Food Syst. 2021, 4, 614739. [Google Scholar] [CrossRef]
- Kandpal, G. Review on Impact of Chemical Fertilizers on Environment. Int. Journ. Modern Agric. 2021, 10, 758–763. [Google Scholar]
- Mrowczyńska-Kamińska, A.; Bajan, B.; Pawłowski, K.P.; Genstwa N, Z.J. Greenhouse Gas Emissions Intensity of Food Production Systems and Its Determinants. PLoS ONE 2021, 16, e0250995. [Google Scholar] [CrossRef]
- Cui, J.; Jin, Z.; Wang, Y.; Gao, S.; Fu, Z.; Yang, Y.; Wang, Y. Mechanism of Eutrophication Process during Algal Decomposition at the Water/Sediment Interface. J. Clean. Prod. 2021, 309, 127175. [Google Scholar] [CrossRef]
- Chaundhry, F.N.; Malik, M.F. Factors Affecting Water Pollution: A Review. J. Ecosyst. Ecography 2017, 7, 5–8. [Google Scholar] [CrossRef]
- Zhao, F.; Zhan, X.; Xu, H.; Zhu, G.; Zou, W. New Insights into Eutrophication Management: Importance of Temperature and Water Residence. J. Environ. Sci. 2022, 111, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Wu, Y.; Liang, P.; Song, Q.; Zhang, H.; Wu, J.; Wu, W.; Liu, X.; Dong, C. Chemosphere Alkaline Amendments Improve the Health of Soils Degraded by Metal Contamination and Acidi Fi Cation: Crop Performance and Soil Bacterial Community Responses. Chemosphere 2020, 257, 127309. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Riaz, U.; Murtaza, G.; Jamil, M.; Ahmed, M.; Hussain, A.; Abbas, Z. Chemical Fertilizers, Formulation, and Their Influence on Soil Health. In Microbiota and Biofertilizers; Springer Nature: Cham, Switzerland, 2021; pp. 1–15. [Google Scholar] [CrossRef]
- Prashar, P.; Shah, S. Chapter 8. Impact of Fertilizers and Pesticides on Soil Microflora in Agriculture. In Microbiota and Biofertilizers; Springer International Publishing: Cham, Switzerland, 2016; Volume 19, pp. 331–361. [Google Scholar] [CrossRef]
- Liu, G.; Xie, H. Simulation of Regulation Policies for Fertilizer and Pesticide Reduction in Arable Land Based on Farmers’ Behavior-Using Jiangxi Province as an Example. Sustainability 2019, 11, 136. [Google Scholar] [CrossRef]
- Villarreal-Delgado, M.F.; Villa-Rodríguez, E.D.; Cira-Chávez, L.A.; Estrada-Alvarado, M.I.; Parra-Cota, F.I.; De los Santos-Villalobos, S. El Género Bacillus Como Agente de Control Biológico y Sus Implicaciones En La Bioseguridad Agrícola. Rev. Mex. Fitopatol. 2018, 36, 95–130. [Google Scholar] [CrossRef]
- de los Santos-Villalobos, S.; Díaz-Rodríguez, A.M.; Ávila-Mascareño, M.F.; Martínez-Vidales, A.D.; Parra-Cota, F.I. Colmena: A Culture Collection of Native Microorganisms for Harnessing the Agro-Biotechnological Potential in Soils and Contributing to Food Security. Diversity 2021, 13, 337. [Google Scholar] [CrossRef]
- Hesham, A.E.-L.; Kaur, T.; Devi, R.; Kour, D.; Prasad, S.; Yadav, N.; Singh, C.; Singh, J.; Yadav, A.N. Current Trends in Microbial Biotechnology for Agricultural Sustainability: Conclusion and Future Challenges. In Environmental and Microbial Biotechnology; Springer Nature: Singapore, 2021; pp. 555–572. [Google Scholar] [CrossRef]
- De los Santos Villalobos, S.; Parra Cota, F.I.; Herrera Sepúlveda, A.; Valenzuela Aragón, B.; Estrada Mora, J.C. Colmena: Colección de Microorganismos Edáficos y Endófitos Nativos, Para Contribuir a La Seguridad Alimentaria Nacional. Rev. Mex. Cienc. Agrícolas 2018, 9, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela-Aragon, B.; Parra-Cota, F.I.; Santoyo, G.; Arellano-Wattenbarger, G.L.; de los Santos-Villalobos, S. Plant-Assisted Selection: A Promising Alternative for in Vivo Identification of Wheat (Triticum Turgidum L. Subsp. Durum) Growth Promoting Bacteria. Plant Soil 2019, 435, 367–384. [Google Scholar] [CrossRef]
- de los Santos-Villalobos, S.; de Folter, S.; Délano-Frier, J.P.; Gómez-Lim, M.A.; Guzmán-Ortiz, D.A.; Peña-Cabriales, J.J. Growth Promotion and Flowering Induction in Mango (Mangifera Indica L. Cv “Ataulfo”) Trees by Burkholderia and Rhizobium Inoculation: Morphometric, Biochemical, and Molecular Events. J. Plant Growth Regul. 2013, 32, 615–627. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A Critical Examination of the Specificity of the Salkowski Reagent for Indolic Compounds Produced by Phytopathogenic Bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of Phosphorus in Soil in Connection with the Vital Activity of Some Microbial Species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Onyia, C.E.; Anyanwu, C.U. Comparative Study on Solubilization of Tri-Calcium Phosphate (TCP) by Phosphate Solubilizing Fungi (PSF) Isolated from Nsukka Pepper Plant Rhizosphere and Root Free Soil. J. Yeast Fungal Res. 2013, 4, 52–57. [Google Scholar] [CrossRef]
- Alexander, D.B.; Zuberer, D.A. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fertl. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Babraham Bioinformatics. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 5 March 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Rissman, A.I.; Mau, B.; Biehl, B.S.; Darling, A.E.; Glasner, J.D.; Perna, N.T. Reordering Contigs of Draft Genomes Using the Mauve Aligner. Bioinformatics 2009, 25, 2071–2073. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In Silico Detection and Typing of Plasmids Using Plasmidfinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of Microbial Genomes Using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, 206–214. [Google Scholar] [CrossRef]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding Data and Analysis Capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef]
- Grant, J.R.; Stothard, P. The CGView Server: A Comparative Genomics Tool for Circular Genomes. Nucleic Acids Res. 2008, 36, 181–184. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; Van Wezel, G.P.; Medema, M.H.; Weber, T. AntiSMASH 6.0: Improving Cluster Detection and Comparison Capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Patel, S.; Saini, N.; Chen, S. Robust Demarcation of 17 Distinct Bacillus Species Clades, Proposed as Novel Bacillaceae Genera, by Phylogenomics and Comparative Genomic Analyses: Description of Robertmurraya Kyonggiensis Sp. Nov. and Proposal for an Emended Genus Bacillus Limiting It O. Int. J. Syst. Evol. Microbiol. 2020, 70, 5753–5798. [Google Scholar] [CrossRef] [PubMed]
- Biedendieck, R.; Knuuti, T.; Moore, S.J.; Jahn, D. The “Beauty in the Beast”—the Multiple Uses of Priestia Megaterium in Biotechnology. Appl. Microbiol. Biotechnol. 2021, 105, 5719–5737. [Google Scholar] [CrossRef]
- Katsenios, N.; Andreou, V.; Sparangis, P.; Djordjevic, N.; Giannoglou, M.; Chanioti, S.; Stergiou, P.; Xanthou, M.Z.; Kakabouki, I.; Vlachakis, D.; et al. Evaluation of Plant Growth Promoting Bacteria Strains on Growth, Yield and Quality of Industrial Tomato. Microorganisms 2021, 9, 2099. [Google Scholar] [CrossRef]
- Pishchik, V.N.; Filippova, P.S.; Mirskaya, G.V.; Khomyakov, Y.V.; Vertebny, V.E.; Dubovitskaya, V.I.; Ostankova, Y.V.; Semenov, A.V.; Chakrabarty, D.; Zuev, E.V.; et al. Epiphytic Pgpb Bacillus Megaterium Afi1 and Paenibacillus Nicotianae Afi2 Improve Wheat Growth and Antioxidant Status under Ni Stress. Plants 2021, 10, 2334. [Google Scholar] [CrossRef]
- López-Bucio, J.; Campos-Cuevas, J.C.; Hernández-Calderón, E.; Velásquez-Becerra, C.; Farías-Rodríguez, R.; Macías-Rodríguez, L.I.; Valencia-Cantero, E. Bacillus Megaterium Rhizobacteria Promote Growth and Alter Root-System Architecture through an Auxin- and Ethylene-Independent Signaling Mechanism in Arabidopsis Thaliana. Int. Soc. Mol. Plant-Microbe Interact. 2007, 20, 207–217. [Google Scholar] [CrossRef]
- Tan, S.; Luschnig, C.; Friml, J. Pho-View of Auxin: Reversible Protein Phosphorylation in Auxin Biosynthesis, Transport and Signaling. Mol. Plant 2021, 14, 151–165. [Google Scholar] [CrossRef]
- Maheshwari, D.K.; Dheeman, S.; Agarwal, M. Phytohormone-Producing PGPR for Sustainable Agriculture. In Bacteruak Metabolites in Sustainable Agroecosystem; Springer International Publishing: Cham, Switzerland, 2015; Volume 12, pp. 159–182. [Google Scholar] [CrossRef]
- Shah, A.; Nazari, M.; Antar, M.; Msimbira, L.A.; Naamala, J.; Lyu, D.; Rabileh, M.; Zajonc, J.; Smith, D.L. PGPR in Agriculture: A Sustainable Approach to Increasing Climate Change Resilience. Front. Sustain. Food Syst. 2021, 5, 1–22. [Google Scholar] [CrossRef]
- Billah, M.; Khan, M.; Bano, A.; Hassan, T.U.; Munir, A.; Gurmani, A.R. Phosphorus and Phosphate Solubilizing Bacteria: Keys for Sustainable Agriculture. Geomicrobiol. J. 2019, 36, 904–916. [Google Scholar] [CrossRef]
- Ahmad, M.; Ahmad, M.; El-Naggar, A.H.; Usman, A.R.A.; Abduljabbar, A.; Elfaki, J.; Al-Faraj, A.; Al-wabel, M.I. Aging Effects of Organic and Inorganic Fertilizers on Phosphorus Fractionation in a Calcareous Sandy Loam Soil. Pedosphere 2017, 28, 873–883. [Google Scholar] [CrossRef]
- Bünemann, E.K. Assessment of Gross and Net Mineralization Rates of Soil Organic Phosphorus—A Review. Soil Biol. Biochem. 2015, 89, 82–98. [Google Scholar] [CrossRef]
- Dash, N.; Pahari, A.; Dangar, T.K. Chapter 7. Functionalities of Phosphate- Solubilizing Bacteria of Rice Rhizosphere: Techniques and Perspectives. In Recent Adv. Appl. Microbiol. Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2017; pp. 1–290. [Google Scholar] [CrossRef]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying Mechanics of Plant Growth Promoting Rhizobacteria (PGPR): A Review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Singh, M.; Singh, D.; Gupta, A.; Pandey, K.D.; Singh, P.K.; Kumar, A. Chapter 3. Plant Growth Promoting Rhizobacteria: Application in Biofertilizers and Biocontrol of Phytopathogens. In PGPR Amelioration in Sustainable Agriculture; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 41–66. [Google Scholar] [CrossRef]
- Chakraborty, U.; Chakraborty, B.; Basnet, M. Plant Growth Promotion and Induction of Resistance in Camellia Sinensis by Bacillus Megaterium. J. Basic Microbiol. 2006, 46, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Solanki, M.K.; Robert, A.S.; Singh, R.K.; Kumar, S.; Pandey, A.K.; Srivastava, A.K.; Arora, D.K. Characterization of Mycolytic Enzymes of Bacillus Strains and Their Bio-Protection Role against Rhizoctonia Solani in Tomato. Curr. Microbiol. 2012, 65, 330–336. [Google Scholar] [CrossRef]
- Mannaa, M.; Oh, J.Y.; Kim, K.D. Biocontrol Activity of Volatile-Producing Bacillus Megaterium and Pseudomonas Protegens against Aspergillus Flavus and Aflatoxin Production on Stored Rice Grains. Mycobiology 2017, 45, 213–219. [Google Scholar] [CrossRef]
- Cochard, B.; Giroud, B.; Crovadore, J.; Chablais, R.; Arminjon, L.; Lefort, F. Endophytic PGPR from Tomato Roots Isolation, In Vitro Characterization and In Vivo Evaluation of Treated Tomatoes. Microorganisms 2022, 10, 765. [Google Scholar] [CrossRef]
- Kordatzaki, G.; Katsenios, N.; Giannoglou, M.; Andreou, V.; Chanioti, S.; Katsaros, G.; Savvas, D.; Efthimiadou, A. Effect of Foliar and Soil Application of Plant Growth Promoting Bacteria on Kale Production and Quality Characteristics. Sci. Hortic. 2022, 301, 111094. [Google Scholar] [CrossRef]
- Abriouel, H.; Franz, C.M.A.P.; Omar, N.B.; Galvez, A. Diversity and Applications of Bacillus Bacteriocins. FEMS Microbiol. Rev. 2011, 35, 201–232. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M.T.; Sciences, B.; Brunswick, N.; Brunswick, N.; Africa, S. Functions and Emerging Applications of Bacteriocins. Current Opinion in Biotechnology. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Hatakka, K.; Saxelin, M. Probiotics in Intestinal and Non-Intestinal Infectious Diseases—Clinical Evidence. Curr. Pharm. Des. 2008, 14, 1351–1367. [Google Scholar] [CrossRef] [PubMed]
- Fathizadeh, H.; Pakdel, F.; Saffari, M.; Esmaeili, D.D.; Momen-Heravi, M.; Dao, S.; Ganbarov, K.; Kafil, H.S. Bacteriocins: Recent Advances in Application as an Antimicrobial Alternative. Curr. Pharm. Biotechnol. 2021, 23, 1028–1040. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins-a Viable Alternative to Antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameter/Treatment | Leaf Number | Increment (%) vs. Control | Stem Diameter, cm | Increment (%) vs. Control | Stem Height, cm | Increment (%) vs. Control | Root Length, cm | Increment (%) vs. Control | Plant Dry Weight, g | Increment (%) vs. Control |
|---|---|---|---|---|---|---|---|---|---|---|
| Non-inoculated (control) | 9.9 ± 2.9 | - | 6.1 ± 1.9 | - | 42.2 ± 5.9 | - | 31.9 ± 2.8 | - | 2.1 ± 0.7 | - |
| Inoculated (strain TSO9) | 16.8 ± 3.2 * | 68.3 | 11.5 ± 2.5 * | 87.9 | 47.3 ± 4.6 | 12.1 | 36.2 ± 3.4 | 13.5 | 2.3 ± 0.5 | 7.9 |
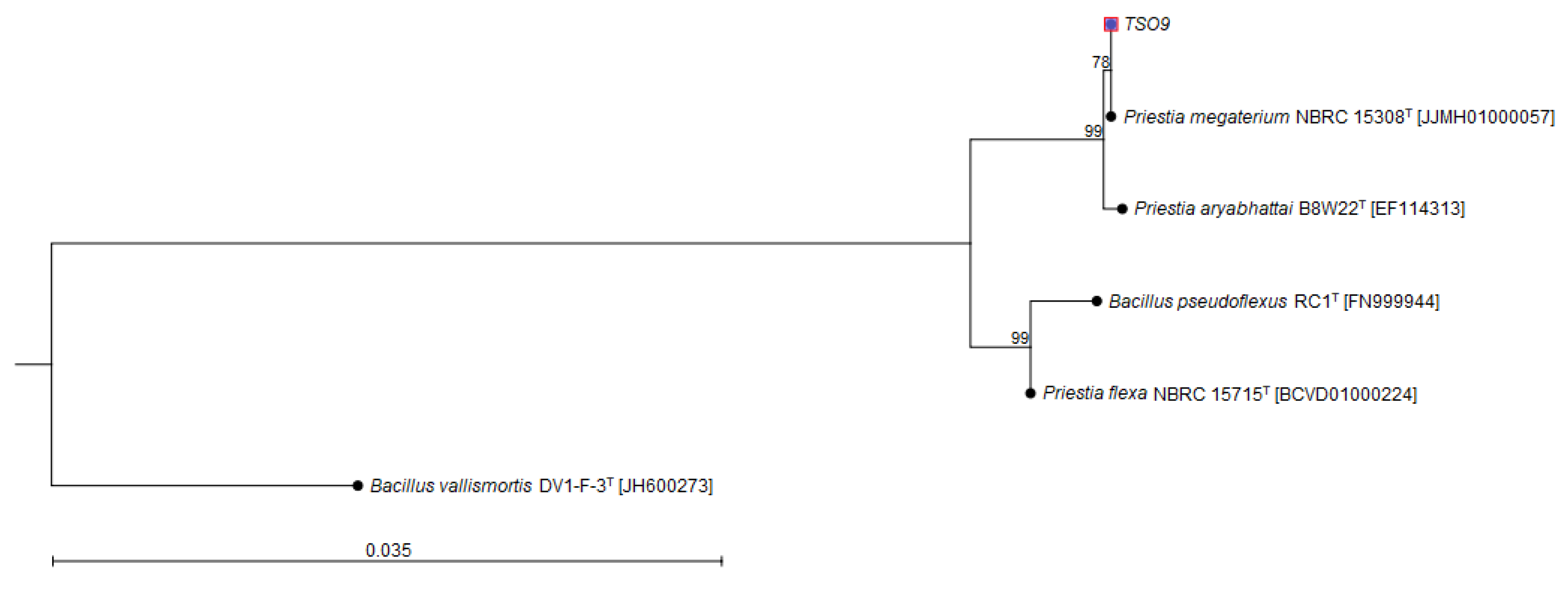
| Taxon Name | Strain | GenBank Accession Number | Similarity (%) |
|---|---|---|---|
| Priestia megaterium | NBRC 15308T | JJMH01000057 | 100 |
| Priestia aryabhattai | B8W22T | EF114313 | 99.86 |
| Priestia flexa | NBRC 15715T | BCVD01000224 | 98.95 |
| Bacillus pseudoflexus | RC1T | FN999944 | 98.72 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortega-Urquieta, M.E.; Valenzuela-Ruíz, V.; Mitra, D.; Hyder, S.; Elsheery, N.I.; Kumar Das Mohapatra, P.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Draft Genome Sequence of Priestia sp. Strain TSO9, a Plant Growth-Promoting Bacterium Associated with Wheat (Triticum turgidum subsp. durum) in the Yaqui Valley, Mexico. Plants 2022, 11, 2231. https://doi.org/10.3390/plants11172231
Ortega-Urquieta ME, Valenzuela-Ruíz V, Mitra D, Hyder S, Elsheery NI, Kumar Das Mohapatra P, Parra-Cota FI, de los Santos-Villalobos S. Draft Genome Sequence of Priestia sp. Strain TSO9, a Plant Growth-Promoting Bacterium Associated with Wheat (Triticum turgidum subsp. durum) in the Yaqui Valley, Mexico. Plants. 2022; 11(17):2231. https://doi.org/10.3390/plants11172231
Chicago/Turabian StyleOrtega-Urquieta, Maria Edith, Valeria Valenzuela-Ruíz, Debasis Mitra, Sajjad Hyder, Nabil I. Elsheery, Pradeep Kumar Das Mohapatra, Fannie Isela Parra-Cota, and Sergio de los Santos-Villalobos. 2022. "Draft Genome Sequence of Priestia sp. Strain TSO9, a Plant Growth-Promoting Bacterium Associated with Wheat (Triticum turgidum subsp. durum) in the Yaqui Valley, Mexico" Plants 11, no. 17: 2231. https://doi.org/10.3390/plants11172231