

A New Approach for Controlling Agrobacterium tumefaciens Post Transformation Using Lytic Bacteriophage
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Characterization
2.1.1. Morphology Plaque Characterization
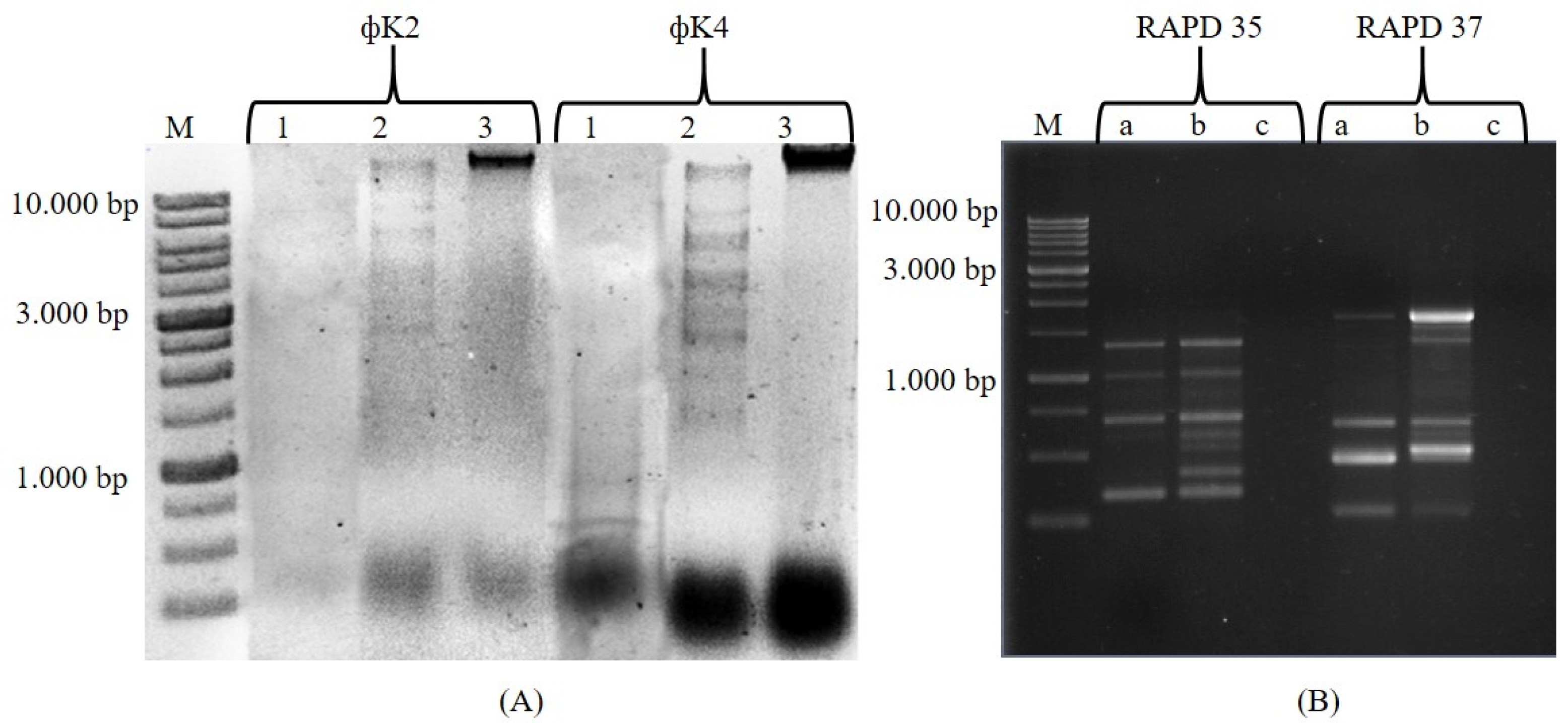
2.1.2. Molecular Characterization Using Restriction Fragment Length Polymorphism (RFLP) and PCR Random Amplified Polymorphic DNA (RAPD)
2.1.3. Host Range Determination
2.2. Stability Physicochemical and Growth Kinetics
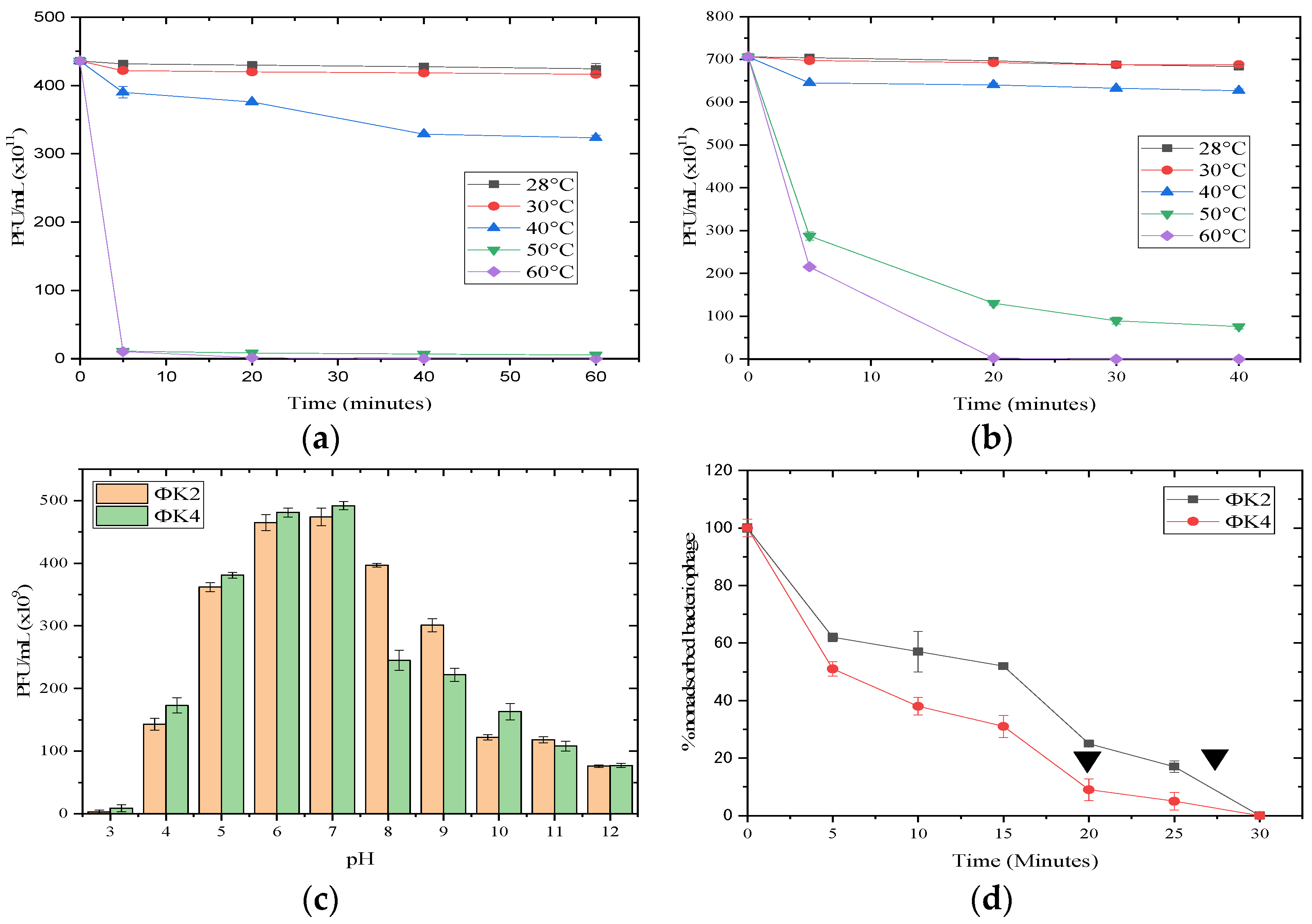
2.2.1. pH and Thermal Stability
2.2.2. Adsorption Assay
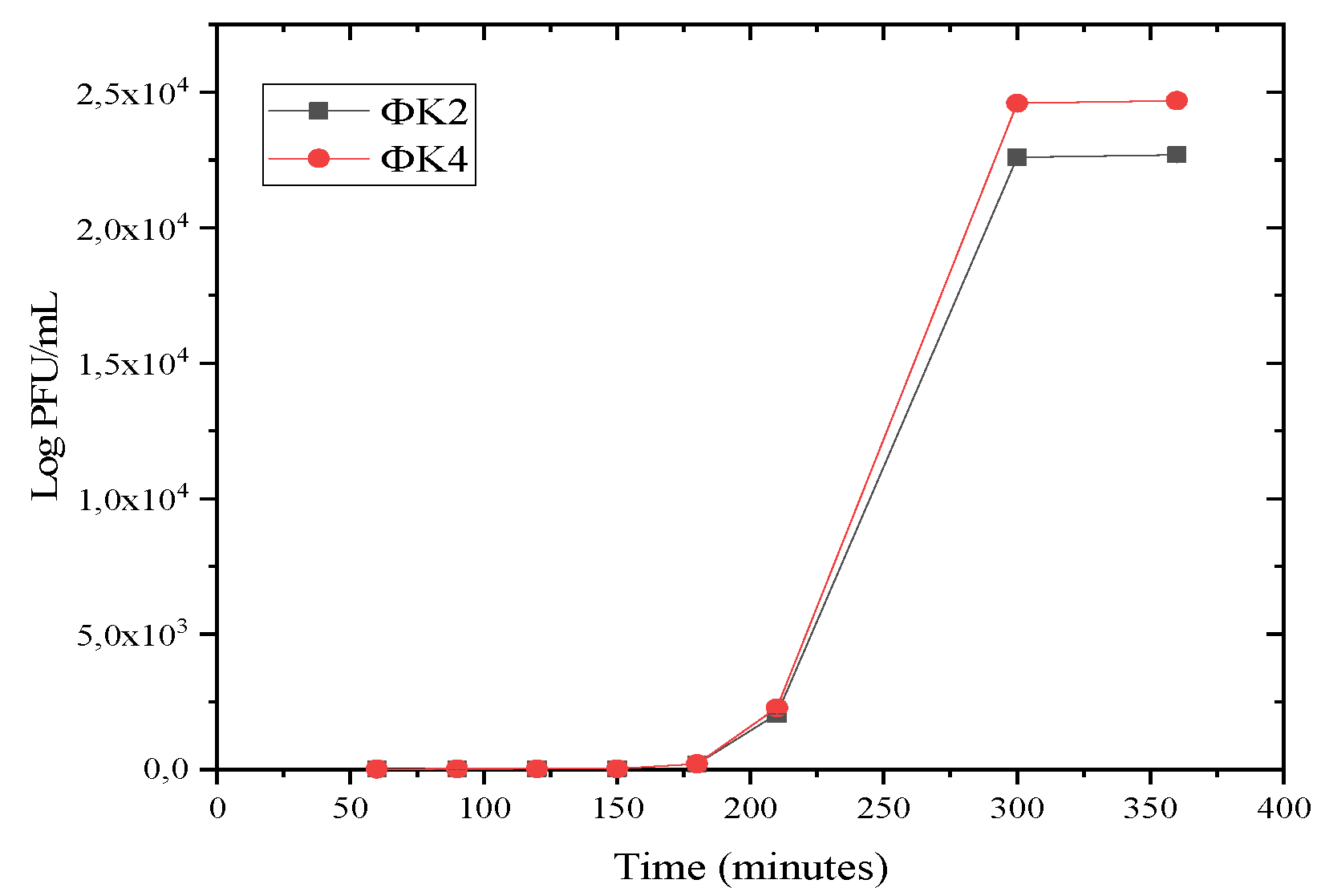
2.2.3. One-Step Growth
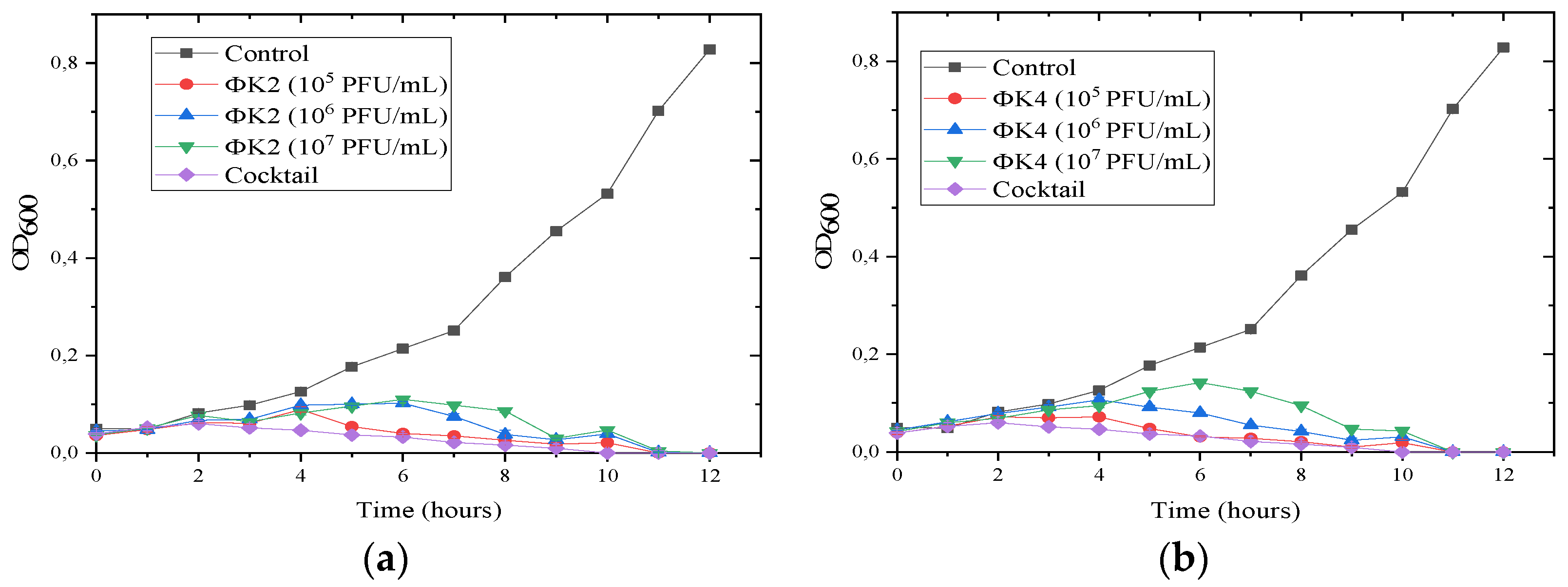
2.2.4. Susceptibility Test
2.3. Controlling Overgrowth of A. tumefaciens Using Lytic Bacteriophage
2.3.1. Simulation Washing
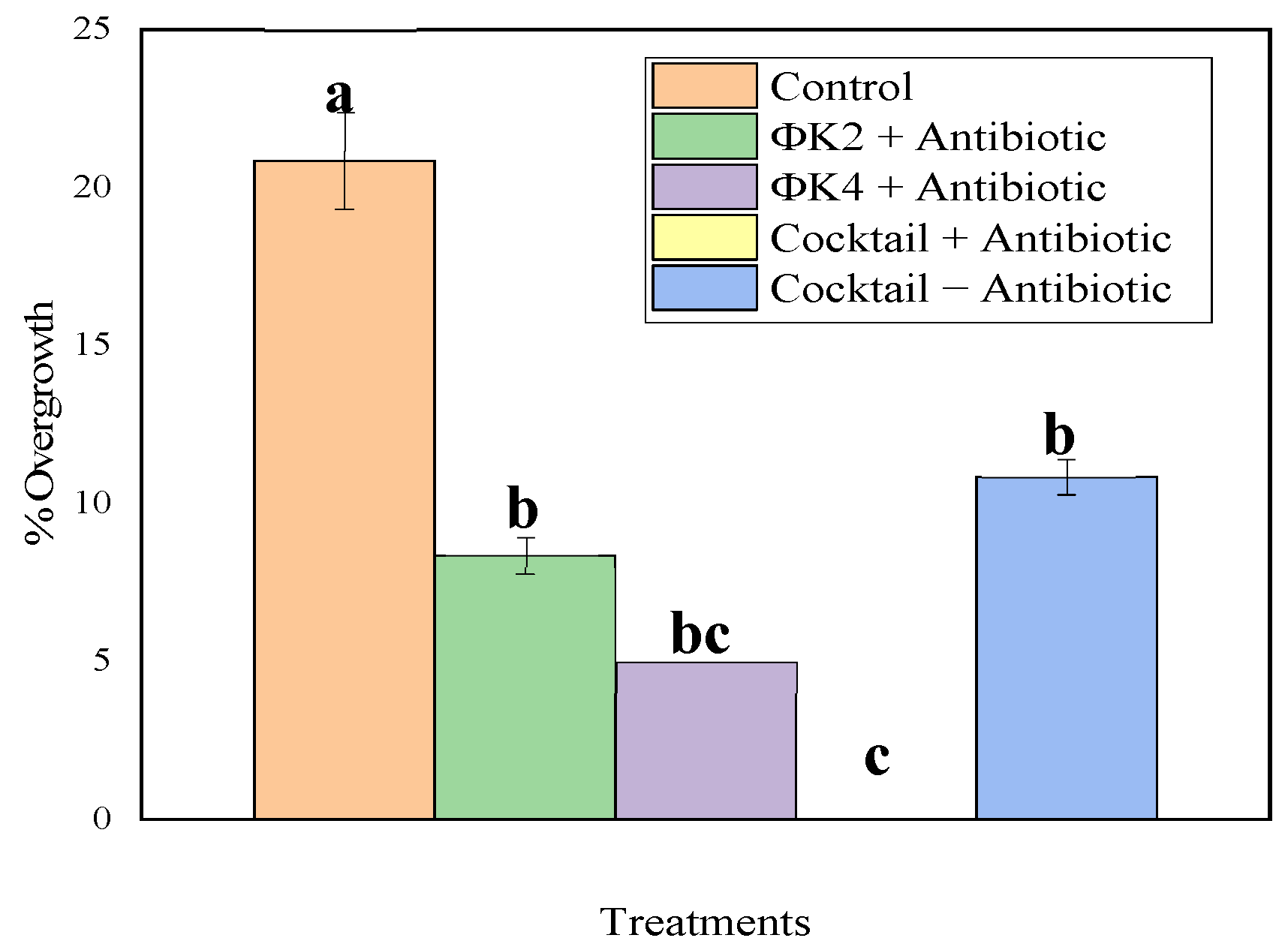
2.3.2. Plant Transformation and Percentage Overgrowth
3. Discussion
4. Materials and Methods
4.1. Characterization
4.1.1. Morphology Plaque and Quantification Bacteriophage
4.1.2. Restriction Fragment Length Polymorphism (RFLP) and PCR Random Amplified Polymorphic DNA (RAPD)
4.1.3. Host Range Determination
4.2. Stability Physicochemical and Growth Kinetics
4.2.1. pH and Thermal Stability
4.2.2. Adsorption Test
4.2.3. One-Step Growth
4.2.4. Susceptible Test
4.3. Controlling Overgrowth of A. tumefaciens Using Lytic Bacteriophage
4.3.1. Simulation Washing
4.3.2. Plant Transformation and Percentage Overgrowth
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yenchon, S.; Te-chato, S. Effect of bacteria density, inoculation and co-cultivation period on Agrobacterium-mediated transformation of oil palm embryogenic callus. J. Agric. Sci. Technol. 2012, 8, 1485–1496. [Google Scholar]
- Sidorov, V.; Gilbertson, L.; Addae, P.; Duncan, D. Agrobacterium-mediated transformation of seedling-derived maize callus. Plant Cell Rep. 2006, 25, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Sandal, I.; Saini, U.; Lacroix, B.; Bhattacharya, A.; Ahuja, P.S.; Citovsky, V. Agrobacterium-mediated genetic transformation of tea leaf explants: Effects of counteracting bactericidity of leaf polyphenols without loss of bacterial virulence. Plant Cell Rep. 2006, 26, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Ogaki, M.; Furuichi, Y.; Kuroda, K.; Chin, D.P.; Ogawa, Y.; Mii, M. Importance of co-cultivation medium pH for successful Agrobacterium-mediated transformation of Lilium x formolongi. Plant Cell Rep. 2008, 27, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Kutty, P.C.; Parveez, G.K.A.; Huyop, F. An easy method for Agrobacterium tumefaciens-mediated gene transfer to Nicotiana tabacum cv. Tapm26. J. Biol. Sci. 2010, 10, 480–489. [Google Scholar] [CrossRef] [Green Version]
- Lessard, P.A.; Kulaveerasingam, H.; York, G.M.; Strong, A.; Sinskey, A. Manipulating gene expression for metabolic engineering of plants. Metab. Eng. 2002, 4, 67–79. [Google Scholar] [CrossRef]
- Coll, N.S.; Epple, P.; Dangl, J.L. Programmed cell death in the plant immune system. Cell Death Differ. 2011, 18, 1247–1256. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Finer, J.J. Low Agrobacterium tumefaciens inoculum levels and a long co-culture period lead to reduced plant defense responses and increase transgenic shoot production of sunflower (Helianthus annus l.). In Vitro Cell. Dev. Biol. Plant. 2016, 52, 354–366. [Google Scholar] [CrossRef] [Green Version]
- Manfroi, E.; Yamazaki-Lau, E.; Grando, M.F.; Roesler, E.A. Acetosyringone, pH and temperature effects on transient genetic transformation of immature embryos of Brazilian wheat genotypes by Agrobacterium tumefaciens. Genet. Mol. Biol. 2015, 38, 470–476. [Google Scholar] [CrossRef]
- de Mayolo, G.A.; Maximova, S.N.; Pisha, S.; Guiltinan, M.J. Moxalactam as a counterselection antibiotic for Agrobacterium mediated transformation and its positive effects on Theobroma cacao somatic embryogenesis. Plant Sci. 2003, 164, 607–615. [Google Scholar] [CrossRef]
- Quisen, R.; de Oliveira, Y.; Pilegg, M.; Cuquel, F.; Quoirin, M. Selective agent and A. tumefaciens overgrowth-control antibiotics in Eucalyptus camaldulensis cotiledonary culture. Braz. Arch. Biol. Technol. 2009, 52, 1485–1492. [Google Scholar] [CrossRef]
- Tamzil, M.S.; Alfiko, Y.; Mubarok, A.F.; Purwantomo, S.; Suwanto, A.; Budiarti, S. Development of Auxotrophic Agrobacterium tumefaciens AGL1 by Tn5 transposon for rice (Oryza sativa L.) transformation. Biotechnol. Bioprocess Eng. 2021, 26, 641–649. [Google Scholar] [CrossRef]
- Liu, Y.; Miao, J.; Traore, S.; Kong, D.; Liu, Y.; Zhang, Z.; Nimchuk, Z.L.; Liu, Z.; Zhao, B. SacB-SacR gene cassette as the negative selection marker to suppress Agrobacterium overgrowth in Agrobacterium-mediated plant transformation. Front. Mol. Biosci. 2016, 3, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, E.; Campbell, K.; Grant, I.; McAuliffe, O. Understanding and exploiting phage-host interaction. Viruses 2019, 11, 567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altamirano, F.L.; Barra, J.J. Phage therapy in the post antibiotic era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar]
- Sohail, H.A.; Coffey, A.; Debrowska, K.; Meyer, I.M.; Middelboe, M.; Sohail, M.; Clokie, M.R.J. Bacteriophages: Emerging applications in medicine, food, and biotechnology. Phage 2020, 1, 75–82. [Google Scholar] [CrossRef]
- Nazir, A.; Dong, Z.; Liu, J.; Tahir, R.A.; Rasheed, M.; Qing, H.; Peng, D.; Tong, Y. Genomic analysis of bacteriophage Xoo-sp13 infecting Xanthomonas oryzae pv. oryzae. Arch. Virol. 2021, 166, 1263–1265. [Google Scholar] [CrossRef]
- Bhunchoth, A.; Phironrit, N.; Leksomboon, C.; Chatchawankanphanich, O.; Kotera, S.; Narulita, E.; Kawasaki, T.; Fujie, M.; Yamada, T. Isolation of Ralstonia solanacearum-infecting bacteriophages from tomato fields in Chiang Mai, Thailand, and their experimental use as biocontrol agents. J. Appl. Microbiol. 2015, 4, 1023–1033. [Google Scholar] [CrossRef]
- Payne, R.J.; Phil, D.; Jansen, V.A. Phage theraphy: The peculiar kinetics of self replicating pharmaceuticals. Clin. Pharmacol. Ther. 2000, 68, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Payne, R.J.; Jansen, V.A. Understanding bacteriophage theraphy as a density-dependant kinetic process. J. Theor. Biol. 2001, 208, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Ly-Chatain, M.H. The factors affecting effectivetness of treatment in phages theraphy. Front. Microbiol. 2014, 5, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, B.K.; Abedon, S.T.; Loc-Carrillo, C. Phage cocktails and future of phage therapy. Future Microbiol. 2013, 8, 769–783. [Google Scholar] [CrossRef] [PubMed]
- Arivo, D.; Rusmana, I.; Budiarti, S. Isolation and characterization of EPEC phage from domestic waste in Indonesia. Malays. J. Microbiol. 2016, 12, 140–146. [Google Scholar]
- Budiarti, S.; Sunarti, R.N.; Listya, S.A.P.; Rusmana, I. Biological characteristics ad lytic effectiveness of phage isolated from domestic wastewater against indigenous Salmonella spp. J. Envirol. Biol. 2019, 40, 53–60. [Google Scholar] [CrossRef]
- Li, B.; Zhang, S.; Long, L.; Huang, S. Characterization and complete genome sequences of three N4-Like roseobacter phages Isolated from the South China Sea. Curr. Microbiol. 2016, 73, 409–418. [Google Scholar] [CrossRef]
- Li, Y.; Wang, M.; Liu, Q.; Song, X.; Wang, D.; Ma, Y.; Shao, H.; Jiang, Y. Complete genomic sequence of bacteriophage H188: A novel Vibrio kanaloae phage isolated from Yellow Sea. Curr. Microbiol. 2016, 72, 628–633. [Google Scholar] [CrossRef]
- Novianty; Rusmana, I.; Budiarti, S. Lytic bacteriophage for Photobacterium damselae isolated from water environment. Int. J. Innov. Res. Sci. Eng. 2014, 2, 549–553. [Google Scholar]
- Maina, A.N.; Mwaura, F.B.; Oyugi, J.; Goulding, D.; Toribio, A.L.; Kariuki, S. Characterization of Vibrio cholerae bacteriophages isolated from the environmental waters of the Lake Victoria Region of Kenya. Curr. Microbiol. 2014, 68, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Malki, K.; Kula, A.; Bruder, K.; Sible, E.; Hatzopoulos, T.; Steidel, S.; Watkins, S.C.; Putonti, C. Bacteriophages isolated from Lake Michigan demonstrate broad host-range across several bacterial phyla. Virol. J. 2015, 12, 164. [Google Scholar] [CrossRef] [Green Version]
- Kusmiatun, A.; Rusmana, I.; Budiarti, S. Characterization of bacteriophage specific to Bacillus pumilus from Ciapus River in Bogor, West Java, Indonesia. Hayati J. Biosci. 2015, 22, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sufa, H.I.; Budiarti, S.; Rusmana, I. Diversity of uropathogenic Escherichia coli lytic phage from Cisadane River, West Java, Indonesia based on morphology and protein molecular weight characteristics. Biodiversitas 2018, 19, 2359–2364. [Google Scholar] [CrossRef]
- Madsena, L.; Bertelsena, S.K.; Dalsgaard, L.; Middelboe, M. Dispersal and survival of Flavobacterium psychrophilum phages in vivo in rainbow trout and in vitro under laboratory conditions: Implications for their use in phage therapy. Appl. Environ. Microbiol. 2013, 79, 4853–4861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, T.; Bera, B.C.; Virmani, N.; Vaid, R.K.; Vashisth, M.; Tripathi, B.N. Isolation and characterization of a novel, T7-like phage against Aeromonas veronii. Virus Genes 2018, 54, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Nagayoshi, Y.; Kumagae, K.; Mori, K.; Tashiro, K.; Nakamura, A.; Fujino, Y.; Hiromasa, Y.; Iwamoto, T.; Kuhara, S.; Ohshima, T.; et al. Physiological properties and genome structure of the Hyperthermophilic filamentous phage φOH3 which infects Thermus thermophilus HB8. Front. Microbiol. 2016, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Alekshun, M.A.; Levy, S.B. Molecular mechanisms of antibacterial multidrug resistance. Cell 2007, 128, 1037–1050. [Google Scholar] [CrossRef] [Green Version]
- Clokie, M.R.J.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Kutter, E.; Sulakvelidze, A. Bacteriophages: Biology and Applications; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Ellis, C.B.; Winters, A.L. Isolation of potential MS2 bacteriophage strains. Biol. Sci. 1969, 85, 336–345. [Google Scholar]
- Haq, I.; Chaudhry, W.N.; Andleeb, S.; Qadri, I. Isolation and partial characterization of a virulent bacteriophage IHQ1 specific for Aeromonas punctata from stream water. Microb. Ecol. 2012, 63, 954–963. [Google Scholar] [CrossRef]
- Barrangou, R.; Yoon, S.-S., Jr.; Breidt, J.F.; Fleming, H.P.; Klaenhammer, T.R. Characterization of six Leuconostoc fallax bacteriophages isolated from an industrial sauerkraut fermentation. Food Microbiol. 2002, 11, 5452–5458. [Google Scholar] [CrossRef] [Green Version]
- Kalatzis, P.G.; Bastías, R.; Kokkari, C.; Katharios, P. isolation and characterization of two lytic bacteriophages, φSt2 and φGrn1; phage therapy application for biological control of Vibrio alginolyticus in aquaculture live feeds. PLoS ONE 2016, 11, e0151101. [Google Scholar] [CrossRef] [Green Version]
- Moineau, S.; Pandian, S.; Klaenhammer, T.R. Restriction/modification systems and restriction endonucleases are more effective on lactococcal bacteriophages that have emerged recently in the dairy industry. Appl. Environ. Microbiol. 1993, 59, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Jensen, E.C.; Schrader, H.S.; Rieland, B.; Thompson, T.L.; Lee, K.W.; Nickerson, K.W.; Kokjohn, T.A. Prevalence of broad-host-range lytic bacteriophages of Sphaerolitus natans, Escherichia coli, and Pseudomonas aeruginosa. Appl. Environ. Microbiol. 1998, 64, 575–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.; Miller, L.A.; Klaenhammer, T.R. In vivo genetic exchange of a functional domain from a type IIA methylase between lactococcal plasmid pTR2030 and a virulent bacteriophage. J. Bacteriol. 1991, 173, 4363–4370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laantoa, E.; Mäntynena, S.; de Colibusc, L.; Marjakangasa, J.; Gillumc, A.; Stuart, D.I.; Ravanttia, J.J.; Huiskonenc; Sundberga, L.R. Virus found in a boreal lake links ssDNA and dsDNA viruses. Proc. Natl. Acad. Sci. USA 2017, 114, 8378–8383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fauquet, C.M.; Mayo, M.A.; Manilof, J.; Desselberger, U.; Ball, L.A. Virus Taxonomy Eight Report of the International Comitte on Taxonomy of Viruses; Elsevier: London, UK, 2005. [Google Scholar]
- Ács, N.; Gambino, M.; Brøndsted, L. Bacteriophage enumeration and detection methods. Front. Microbiol. 2020, 11, 2662. [Google Scholar] [CrossRef]
- Addablah, A.A.; Kakou-Ngazoa, S.; Akpa, E.E.; Adioumani, E.; Ndombi, F.M.-B.; Aoussi, S.; Dosso, M. RAPD-based evaluation revealed genetically diverse populations of Pseudomonas aeruginosa and Staphylococcus aureus lytic bacteriophages isolated in urban sewage and Ebrie Lagoon, Côte d’Ivoire. Afr. J. Microbiol. Res. 2021, 15, 522–528. [Google Scholar]
- Dini, C.; de Urraza, P.J. Isolation and selection of coliphages as potential biocontrol agents of enterohemorrhagic and Shiga toxin-producing E. coli (EHEC and STEC) in cattle. J. Appl. Microbiol. 2010, 109, 873–887. [Google Scholar] [CrossRef] [Green Version]
- Lin, B.C.; Kado, C.I. Studies on Agrobacterium tumefaciens. VII. avirulence induced by temperature and ethidium bromide. Can. J. Microbiol. 1977, 23, 1554–1561. [Google Scholar] [CrossRef]
- Kovacs, L.S.G.; Pueppke, S.G. Mapping and genetic organization of pTiChry5, a novel Ti plasmid from a highly virulent Agrobacterium tumefaciens strain. Mol. Gen. Genet. 1994, 242, 327–336. [Google Scholar] [CrossRef]
- Montoya, A.L.; Chilton, M.-D.; Gordon, M.P.; Sciaky, D.; Nester, E.W. Octopine and nopaline metabolism in Agrobacterium tumefaciens and crown gall tumor cells: Role of plasmid genes. J. Bacteriol. 1977, 129, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Hood, E.E.; Helmer, G.L.; Fraley, R.T.; Chilton, M.D. The hypervirulence of Agrobacterium tumefaciens A281 is encoded in a region of pTiBo542 outside of T-DNA. J. Bacteriol. 1986, 168, 1291–1301. [Google Scholar] [CrossRef] [Green Version]
- Koncz, C.; Schell, J. The promoter of TL-DNA gene 5 controls the tissue-specific expression of chimaeric genes carried by a novel type of Agrobacterium binary vector. Mol. Gen. Genet. 1986, 204, 383–396. [Google Scholar] [CrossRef]
- Lazo, G.R.; Stein, P.A.; Ludwig, R.A. A DNA transformation–competent Arabidopsis genomic library in Agrobacterium. Nat. Biotechnol. 1991, 9, 963–967. [Google Scholar] [CrossRef]
- Ooms, G.; Hooykaas, P.J.J.; van Veen, R.J.M.; van Beelen, P.; Regensburg-Tuïnk, T.J.G.; Schilperoort, R.A. Octopine Ti-plasmid deletion mutants of Agrobacterium tumefaciens with emphasis on the right side of the T-region. Plasmid 1982, 7, 15–29. [Google Scholar] [CrossRef]
- Attai, H.; Brown, P.J.B. Isolation and characterization T4- and T7-Like phages that infect the bacterial plant pathogen Agrobacterium tumefaciens. Viruses 2019, 11, 528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attai, H.; Rimbey, J.; Smith, G.P.; Brown, P.J.B. Expression of a peptidoglycan hydrolase from lytic bacteriophages Atu_ph02 and Atu_ph03 triggers lysis of Agrobacterium tumefaciens. Appl. Environ. Microbiol. 2017, 83, e01498-17. [Google Scholar] [CrossRef] [PubMed]
- Jończyk-Matysiak, E.; Łodej, N.; Kula, D.; Owczarek, B.; Orwat, F.; Międzybrodzki, R. Factors determining phage stability/activity: Challenges in practical phage application. Expert Rev. Anti-Infect. Ther. 2019, 17, 583–606. [Google Scholar] [CrossRef]
- Toki, S.; Hara, N.; Ono, K.; Onodera, H.; Tagiri, A.; Oka, S.; Tanaka, H. Early infection of scutellum tissue with Agrobacterium allows high-speed transformation of rice. Plant J. 2006, 47, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Thanki, A.M.; Taylor-Joyce, G.; Dowah, A.; Nale, J.Y.; Malik, D.; Clokie, M.R.J. Unravelling the links between phage adsorption and successful infection in Clostridium difficile. Viruses 2018, 10, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Mandlaa; Wen, H.; Ma, L.; Chen, Z. Isolation, characterization and application of bacteriophage PSDA-2 against Salmonella Typhimurium in chilled mutton. PLoS ONE 2022, 17, e0262946. [Google Scholar] [CrossRef] [PubMed]
- Schmerer, M.; Molineux, I.J.; Bull, J.J. Synergy as a rationale for phage therapy using phage cocktails. PeerJ 2014, 2, e590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merabishvili, M.; Pirnay, J.P.; de Vos, D. Guidelines to compose an ideal bacteriophage cocktail. Methods Mol. Biol. 2018, 1693, 99–110. [Google Scholar] [PubMed]
- Fazzino, L.; Anisman, J.; Chacon, J.M.; Harcombe, W.R. Phage cocktail strategies for the suppression of a pathogen in across-feeding coculture. Microb. Biotechnol. 2020, 13, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T.; Danis-Wlodarczyk, K.M.; Wozniak, D.J. Phage cocktail development for bacteriophage therapy: Toward improving spectrum of activity breadth and depth. Pharmaceuticals 2021, 14, 1019. [Google Scholar] [CrossRef]
- Czajkowski, R.; Ozymko, Z.; de Jager, V.; Siwinska, J.; Smolarska, A.; Ossowicki, A.; Narajczyk, M.; Lojkowska, E. Genomic, proteomic and morphological characterization of two novel broad host lytic bacteriophages ϕPD10.3 and ϕ PD23.1 infecting pectinolytic Pectobacterium spp. and Dickeya spp. PLoS ONE 2015, 10, e0119812. [Google Scholar] [CrossRef] [Green Version]
- Rombouts, S.; Volckaert, A.; Venneman, S.; Declercq, B.; Vandenheuvel, D.; Allonsius, C.N.; van Malderghem, C.; Jang, H.B.; Briers, Y.; Noben, J.P.; et al. Characterization of novel bacteriophages for biocontrol of bacterial blight in leek caused by Pseudomonas syringae pv. porri. Front. Microbiol. 2016, 15, 279. [Google Scholar] [CrossRef]
- Carstens, A.B.; Djurhuus, A.M.; Kot, W.; Jacobs-Sera, D.; Hatfull, G.F.; Hansen, L.H. Unlocking the potential of 46 new bacteriophages for biocontrol of Dickeya solani. Viruses 2018, 10, 621. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, B.; Jailani, A.; Lee, J.-H.; Lee, J. Effect of halogenated indoles on biofilm formation, virulence, and root surface colonization by Agrobacterium tumefaciens. Chemosphere 2022, 293, 133603. [Google Scholar] [CrossRef]
- Jailani, A.; Ahmed, B.; Lee, J.-H.; Lee, J. Inhibition of Agrobacterium tumefaciens growth and biofilm formation by tannic acid. Biomedicines 2022, 10, 1619. [Google Scholar] [CrossRef]
- Tomlinson, A.D.; Fuqua, C. Mechanisms and regulation of polar surface attachment in Agrobacterium tumefaciens. Curr. Opin. Microbiol. 2009, 12, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.K.; Abedon, S.T. Bacteriophages and their enzymes in biofilm control. Curr. Pharm. Des. 2015, 21, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Bacteriophage Ecology: Population Growth, Evolution, and Impact of Bacterial Viruses; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Parasion, S.; Kwiatek, M.; Gryko, R.; Mizak, L.; Malm, A. Bacteriophages as an alternative strategy for fighting biofilm development. Pol. J. Microbiol. 2014, 63, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Gallet, R.; Shao, Y.; Wang, I.N. High adsorption rate is detrimental to bacteriophage fitness in a biofilm-like environment. BMC Evol. Biol. 2009, 9, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, D.; Vandenheuvel, D.; Martínez, B.; Rodríguez, A.; Lavigne, R.; García, P. Two phages, phiIPLA-RODI and phiIPLA-C1C, lyse mono- and dual-species staphylococcal biofilms. Appl. Environ. Microbiol. 2015, 81, 3336–3348. [Google Scholar] [CrossRef] [Green Version]
- Cerca, N.; Oliveira, R.; Azeredo, J. Susceptibility of Staphylococcus epidermidis planktonic cells and biofilms to the lytic action of Staphylococcus bacteriophage K. Lett. Appl. Microbiol. 2007, 45, 313–317. [Google Scholar] [CrossRef] [Green Version]
- Fu, W.; Forster, T.; Mayer, O.; Curtin, J.J.; Lehman, S.M.; Donlan, R.M. Bacteriophage cocktail for the prevention of biofilm formation by Pseudomonas aeruginosa on catheters in an in vitro model system. Antimicrob. Agents Chemother. 2010, 54, 397–404. [Google Scholar] [CrossRef]
- Li, X.; He, Y.; Wang, Z.; Wei, J.; Hu, T.; Si, J.; Tao, G.; Zhang, L.; Xie, L.; Abdalla, A.E.; et al. A combination therapy of phages and antibiotics: Two is better than one. Int. J. Biol. Sci. 2021, 17, 3573–3582. [Google Scholar] [CrossRef]
- Manohar, P.; Royam, M.M.; Loh, B.; Bozdogan, B.; Nachimuthu, R.; Leptihn, S. Synergistic effects of phage−antibiotic combinations against Citrobacter amalonaticus. ACS Infect. Dis. 2022, 8, 59–65. [Google Scholar] [CrossRef]
- Letrado, P.; Corsini, B.; Dıez-Martınez, R.; Bustamante, N.; Yuste, J.E.; Garcıa, P. Bactericidal synergism between antibiotics and phage endolysin Cpl-711 to kill multidrug-resistant pneumococcus. Future Microbiol. 2018, 13, 1215–1223. [Google Scholar] [CrossRef] [Green Version]
- Topka, G.; Bloch, S.; Nejman-Falenczyk, B.; Gasior, T.; Jurczak-Kurek, A.; Necel, A.; Dydecka, A.; Richert, M.; Wegrzyn, G.; Wegrzyn, A. Characterization of bacteriophage vB-EcoS-95, isolated From urban sewage and revealing extremely rapid lytic development. Front. Microbiol. 2019, 9, 3326. [Google Scholar] [CrossRef]
- Lingga, R.; Budiarti, S.; Rusmana, I.; Wahyudi, A.T. Isolation, characterization and efficacy of lytic bacteriophages against pathogenic Escherichia coli from hospital liquid waste. Biodiversitas 2020, 21, 3234–3241. [Google Scholar] [CrossRef]
- Narulita, E.; Sulistiyorini, I.; Aji, G.P.; Iqbal, M.; Murdiyah, S. Isolation and characterization of bacteriophage in controlling Escherichia coli in Jember Area, Indonesia. Asian J. Microbiol. Biotechnol. Environ. Sci. 2018, 20, 439–444. [Google Scholar]
- Pallavi, B.; Puneeth, T.G.; Shekar, M.; Girisha, S.K. Isolation, characterization and genomic analysis of vB-AhyM-AP1, a lytic bacteriophage infecting Aeromonas hydrophila. J. Appl. Microbiol. 2021, 131, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Necel, A.; Bloch, S.; Nejman-Faleńczyk, B.; Grabski, M.; Topka, G.; Dydecka, A.; Kosznik-Kwaśnicka, K.; Grabowski, Ł.; Jurczak-Kurek, A.; Wołkowicz, T.; et al. Characterization of a bacteriophage, vb_Eco4M-7, that effectively infects many Escherichia coli O157 strains. Sci. Rep. 2020, 10, 3743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kropinski, A.M. practical advice on the one-step growth curve. Methods Mol. Biol. 2018, 1681, 41–47. [Google Scholar] [PubMed]
- Zhao, J.; Zhang, Z.; Tian, C.; Chen, X.; Hu, L.; Wei, X.; Li, H.; Lin, W.; Jiang, A.; Feng, R.; et al. Characterizing the biologyof lytic bacteriophage vB_EaeM_’Eap-3 infecting multidrug-resistant Enterobacter aerogenes. Front. Microbiol. 2019, 10, 420. [Google Scholar] [CrossRef] [PubMed]
- Muturi, P.; Yu, J.; Maina, A.N.; Kariuki, S.; Mwaura, F.B.; Wei, H. Bacteriophage isolated in China for the control of Pectobacterium carotovorum causing potato soft rot in Kenya. Virol. Sin. 2019, 34, 287–294. [Google Scholar] [CrossRef]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the TDNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage | Plaque Morphology | Plaque Diameter (mm) | Titer Phage (PFU/mL) |
|---|---|---|---|
| ΦK2 | Clear, without halo | ±1.5 | 5.4 × 1017 |
| ΦK4 | Clear, without halo | ±2.3 | 7.8 × 1017 |
| Bacterial Strain | Isolate Bacteriophage | |
|---|---|---|
| ΦK2 | ΦK4 | |
| Agrobacterium tumefaciens | ||
| AGL1 WT | + | + |
| AGL1 Auxotroph Cystein | + | + |
| EHA105 | + | + |
| LB4404 | − | − |
| Escherichia coli | ||
| DH5α | − | − |
| Top10 | − | − |
| BL21 (DE3) | − | − |
| Bacteriophage Concentration (PFU/mL) | ΦK2 | ΦK4 | Cocktail | |||
|---|---|---|---|---|---|---|
| 40 min (Mean ± SD) | 180 min (Mean ± SD) | 40 min (Mean ± SD) | 180 min (Mean ± SD) | 40 min (Mean ± SD) | 180 min (Mean ± SD) | |
| 107 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 108 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| 109 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Detail Treatment | Control | ΦK2 + Antibiotic | ΦK4 + Antibiotic | Cocktail + Antibiotic | Cocktail − Antibiotic |
|---|---|---|---|---|---|
| Washing step | |||||
| Cefotaxime 300 mg/L | √ | √ | √ | √ | × |
| CaCl2/MgCl2 | √ | × | × | × | × |
| Titer phage (1011 PFU/mL) | × | √ | √ | √ | √ |
| Selection medium | |||||
| Cefotaxime 300 mg/L | √ | √ | √ | √ | × |
| 5 μL drop of phage cocktail | × | × | × | × | √ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramadhan, F.; Alfiko, Y.; Purwantomo, S.; Mubarok, A.F.; Budinarta, W.; Suwanto, A.; Budiarti, S. A New Approach for Controlling Agrobacterium tumefaciens Post Transformation Using Lytic Bacteriophage. Plants 2022, 11, 3124. https://doi.org/10.3390/plants11223124
Ramadhan F, Alfiko Y, Purwantomo S, Mubarok AF, Budinarta W, Suwanto A, Budiarti S. A New Approach for Controlling Agrobacterium tumefaciens Post Transformation Using Lytic Bacteriophage. Plants. 2022; 11(22):3124. https://doi.org/10.3390/plants11223124
Chicago/Turabian StyleRamadhan, Fiqih, Yuzer Alfiko, Sigit Purwantomo, Andhika Faisal Mubarok, Widyah Budinarta, Antonius Suwanto, and Sri Budiarti. 2022. "A New Approach for Controlling Agrobacterium tumefaciens Post Transformation Using Lytic Bacteriophage" Plants 11, no. 22: 3124. https://doi.org/10.3390/plants11223124