
Optimization of Molecular Methods for Detecting Duckweed-Associated Bacteria
 , , , , , and
, , , , , and 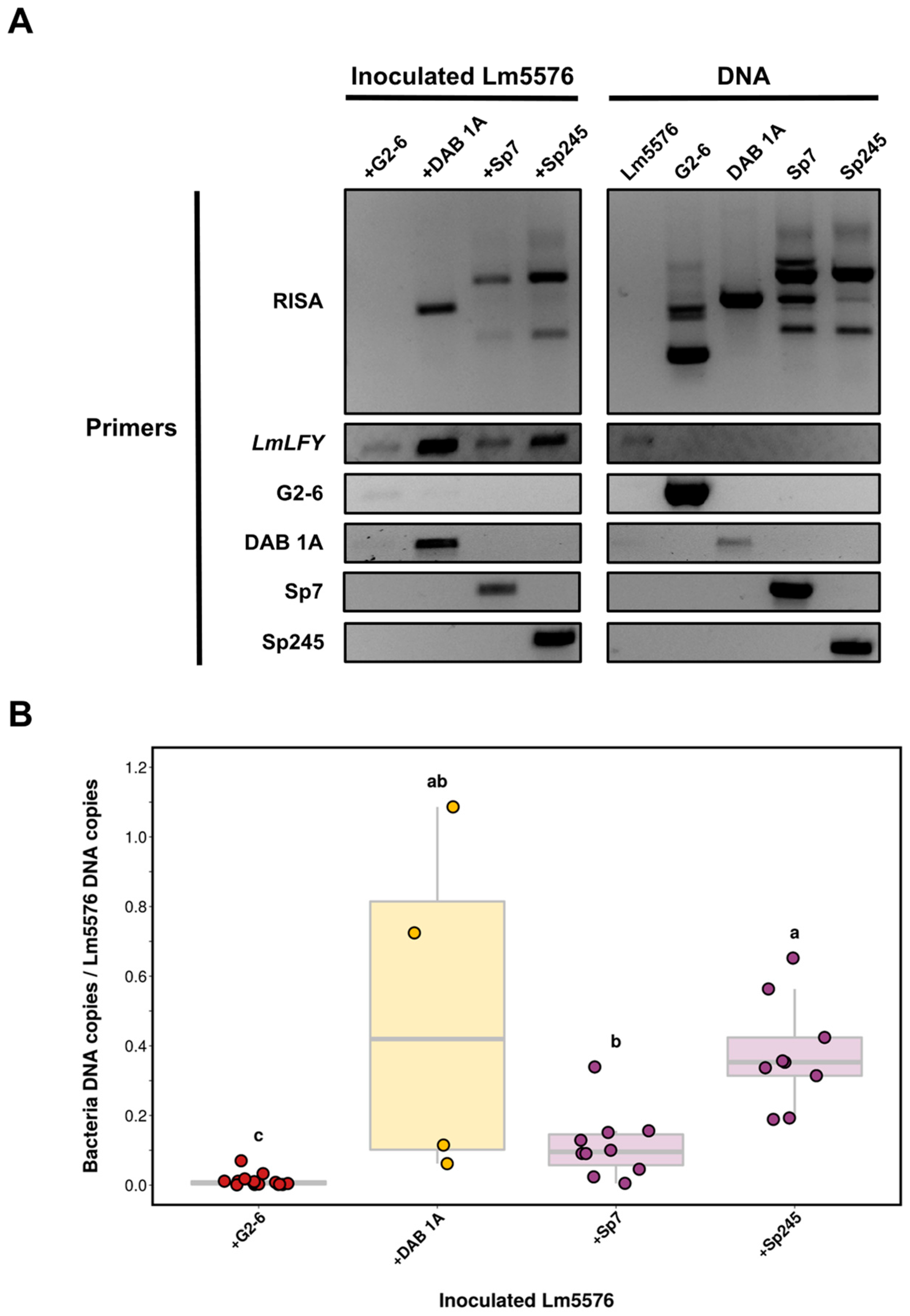{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Selection of Duckweed Strain and Bacterial Isolates
2.2. Systematic Optimization of a Nucleic Acid Extraction Method for Duckweed and Bacteria
2.3. Establishment of a PCR-Based DNA Fingerprinting Assay for Detecting Duckweed–Associated Bacteria
2.4. Standardization and Validation of Attachment PCR Assay for Detecting Duckweed–Associated Bacteria
2.5. Computational Pipeline for Primer Design to Detect and Quantify Specific Duckweed-Associated Bacteria
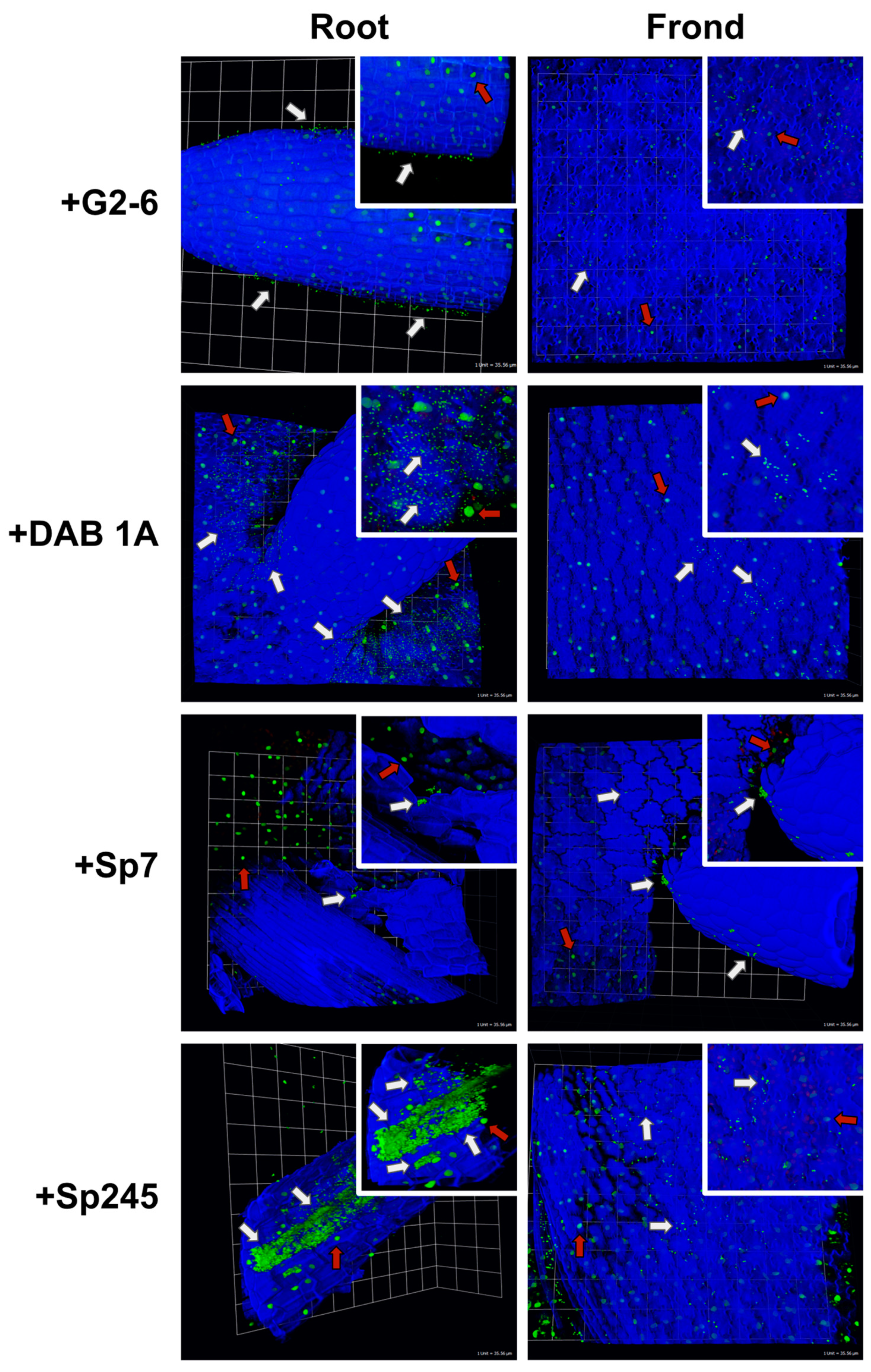
2.6. Bacterial Colonization of Lemna minor Visualized by Confocal Microscopy
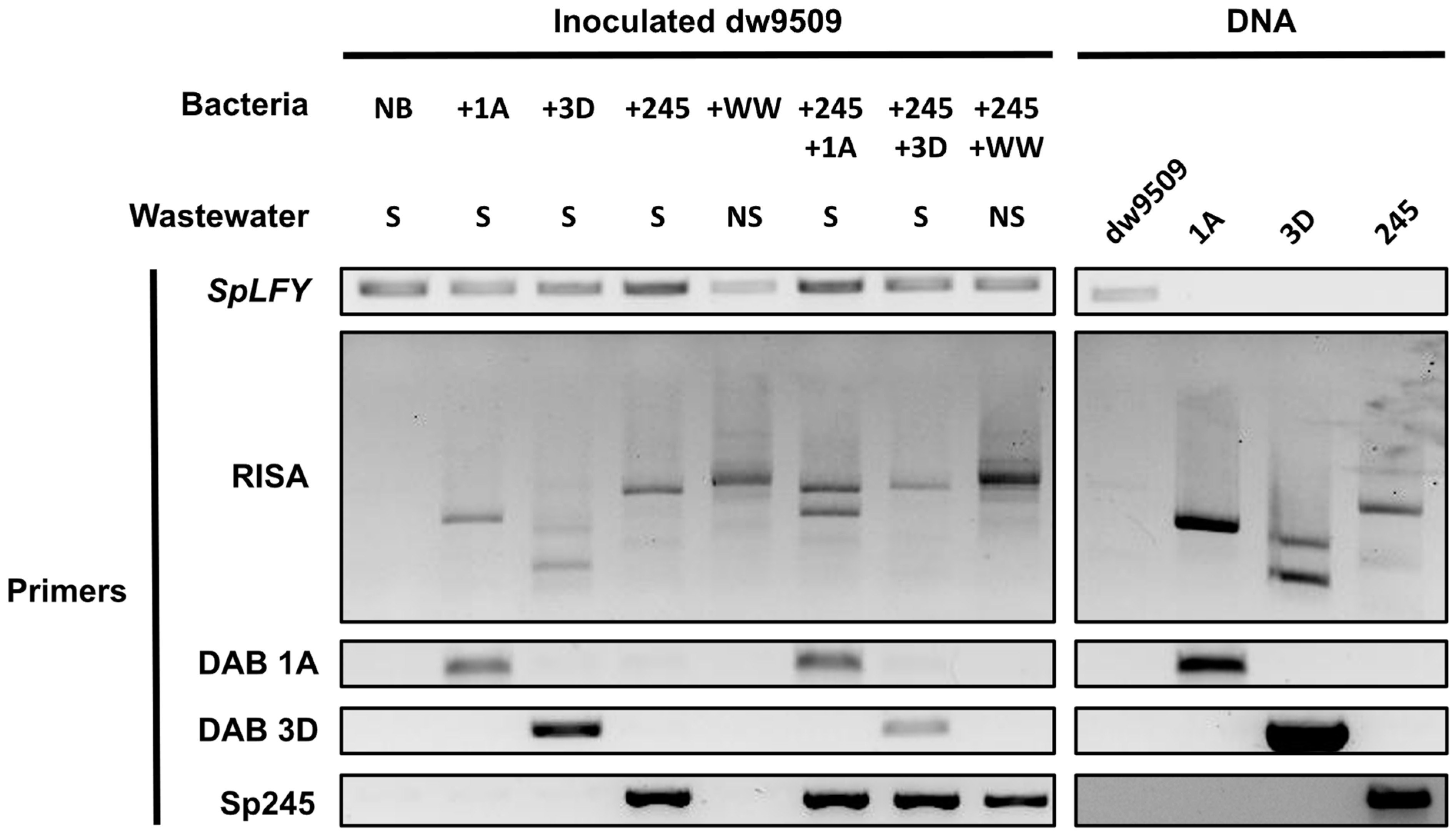
2.7. Strain-Specific Monitoring of Duckweed–Associated Bacteria in a Community Context
3. Discussion
3.1. Localization of Bacteria on Duckweed
3.2. Molecular Methods for Detection of Duckweed-Associated Bacteria
3.3. Bacterial Adaptation to Plant Habitats and Colonization Dynamics
3.4. Molecular Detection of Specific Duckweed–Associated Bacteria within a Community Context
4. Conclusions
5. Materials and Methods
5.1. Duckweed Sterilization and Propagation
5.2. Isolation and Identification of Bacteria
5.3. Genome Sequencing of Bacterial Isolates
5.4. Inoculation of Duckweed with Bacteria
5.5. Nucleic Acid Isolation from Duckweed and Bacteria
5.6. Molecular Detection of Duckweed-Associated Bacteria by rDNA Intergenic Spacer Analysis (RISA)
5.7. Confocal Microscopy of Lm5576 Colonized by Bacteria
5.8. Strain-Specific Primer Design and End-Point PCR
5.9. Quantification of Bacteria Colonization Levels on Duckweed
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.K.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a Model Plant System in the Genomics and Postgenomics Era. Plant Cell 2021, 33, 3207–3234. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, C.R.; Salas-González, I.; Conway, J.M.; Finkel, O.M.; Gilbert, S.; Russ, D.; Teixeira, P.J.P.L.; Dangl, J.L. The Plant Microbiome: From Ecology to Reductionism and Beyond. Annu. Rev. Microbiol. 2020, 74, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Acosta, K.; Xu, J.; Gilbert, S.; Denison, E.; Brinkman, T.; Lebeis, S.; Lam, E. Duckweed Hosts a Taxonomically Similar Bacterial Assemblage as the Terrestrial Leaf Microbiome. PLoS ONE 2020, 15, e0228560. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.-Y.; Su, J.-Q.; Zhu, Y.-G. Phyllosphere Bacterial Community of Floating Macrophytes in Paddy Soil Environments as Revealed by Illumina High-Throughput Sequencing. Appl. Environ. Microbiol. 2015, 81, 522–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, D.; Hiroshima, N.; Ishizawa, H.; Ike, M. Whole Structures, Core Taxa, and Functional Properties of Duckweed Microbiomes. Bioresour. Technol. Rep. 2022, 18, 101060. [Google Scholar] [CrossRef]
- Beilsmith, K.; Perisin, M.; Bergelson, J. Natural Bacterial Assemblages in Arabidopsis thaliana Tissues Become More Distinguishable and Diverse during Host Development. mBio 2021, 12, e02723-20. [Google Scholar] [CrossRef]
- Hacquard, S.; Garrido-Oter, R.; González, A.; Spaepen, S.; Ackermann, G.; Lebeis, S.; McHardy, A.C.; Dangl, J.L.; Knight, R.; Ley, R.; et al. Microbiota and Host Nutrition across Plant and Animal Kingdoms. Cell Host Microbe 2015, 17, 603–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.R.; Wiesmann, C.L.; Melnyk, R.A.; Hossain, S.S.; Chi, M.-H.; Martens, K.; Craven, K.; Haney, C.H. Commensal Pseudomonas Fluorescens Strains Protect Arabidopsis from Closely Related Pseudomonas Pathogens in a Colonization-Dependent Manner. mBio 2022, 13, e02892-21. [Google Scholar] [CrossRef]
- Innerebner, G.; Knief, C.; Vorholt, J.A. Protection of Arabidopsis thaliana against Leaf-Pathogenic Pseudomonas syringae by Sphingomonas Strains in a Controlled Model System. Appl. Environ. Microbiol. 2011, 77, 3202–3210. [Google Scholar] [CrossRef] [Green Version]
- Hassani, M.A.; Amine Hassani, M.; Durán, P.; Hacquard, S. Microbial Interactions within the Plant Holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef]
- Wippel, K.; Tao, K.; Niu, Y.; Zgadzaj, R.; Kiel, N.; Guan, R.; Dahms, E.; Zhang, P.; Jensen, D.B.; Logemann, E.; et al. Host Preference and Invasiveness of Commensal Bacteria in the Lotus and Arabidopsis Root Microbiota. Nat. Microbiol. 2021, 6, 1150–1162. [Google Scholar] [CrossRef] [PubMed]
- Van Veen, J.A.; van Overbeek, L.S.; van Elsas, J.D. Fate and Activity of Microorganisms Introduced into Soil. Microbiol. Mol. Biol. Rev. 1997, 61, 121–135. [Google Scholar] [PubMed]
- Gamalero, E.; Lingua, G.; Berta, G.; Lemanceau, P. Methods for Studying Root Colonization by Introduced Beneficial Bacteria. Sustain. Agric. 2009, 601–615. [Google Scholar] [CrossRef] [Green Version]
- Romano, I.; Ventorino, V.; Pepe, O. Effectiveness of Plant Beneficial Microbes: Overview of the Methodological Approaches for the Assessment of Root Colonization and Persistence. Front. Plant Sci. 2020, 11, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katagiri, F.; Thilmony, R.; He, S.Y. The Arabidopsis Thaliana-Pseudomonas Syringae Interaction. Arabidopsis Book Am. Soc. Plant Biol. 2002, 1, e0039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tornero, P.; Dangl, J.L. A High-Throughput Method for Quantifying Growth of Phytopathogenic Bacteria in Arabidopsis thaliana. Plant J. 2002, 28, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Haney, C.H.; Samuel, B.S.; Bush, J.; Ausubel, F.M. Associations with Rhizosphere Bacteria Can Confer an Adaptive Advantage to Plants. Nat. Plants 2015, 1, 15051. [Google Scholar] [CrossRef] [Green Version]
- Zinniel, D.K.; Lambrecht, P.; Harris, N.B.; Feng, Z.; Kuczmarski, D.; Higley, P.; Ishimaru, C.A.; Arunakumari, A.; Barletta, R.G.; Vidaver, A.K. Isolation and Characterization of Endophytic Colonizing Bacteria from Agronomic Crops and Prairie Plants. Appl. Environ. Microbiol. 2002, 68, 2198–2208. [Google Scholar] [CrossRef] [Green Version]
- Niu, B.; Paulson, J.N.; Zheng, X.; Kolter, R. Simplified and Representative Bacterial Community of Maize Roots. Proc. Natl. Acad. Sci. USA 2017, 114, E2450–E2459. [Google Scholar] [CrossRef] [Green Version]
- O’Banion, B.S.; O’Neal, L.; Alexandre, G.; Lebeis, S.L. Bridging the Gap Between Single-Strain and Community-Level Plant-Microbe Chemical Interactions. Mol. Plant. Microbe Interact. 2020, 33, 124–134. [Google Scholar] [CrossRef]
- Reinhold-Hurek, B.; Hurek, T. Life in Grasses: Diazotrophic Endophytes. Trends Microbiol. 1998, 6, 139–144. [Google Scholar] [CrossRef]
- Kandel, S.L.; Joubert, P.M.; Doty, S.L. Bacterial Endophyte Colonization and Distribution within Plants. Microorganisms 2017, 5, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, E.K.; Gyaneshwar, P.; Mathan, N.; Barraquio, W.L.; Reddy, P.M.; Iannetta, P.P.M.; Olivares, F.L.; Ladha, J.K. Infection and Colonization of Rice Seedlings by the Plant Growth-Promoting Bacterium Herbaspirillum seropedicae Z67. Mol. Plant. Microbe Interact. 2002, 15, 894–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloter, M.; Hartmann, A. Endophytic and Surface Colonization of Wheat Roots (Triticum aestivum) by Different Azospirillum brasilense Strains Studied with Strain-Specific Monoclonal Antibodies. Symbiosis 1998, 25, 159–179. [Google Scholar]
- Yamaga, F.; Washio, K.; Morikawa, M. Sustainable Biodegradation of Phenol by Acinetobacter calcoaceticus P23 Isolated from the Rhizosphere of Duckweed Lemna aoukikusa. Environ. Sci. Technol. 2010, 44, 6470–6474. [Google Scholar] [CrossRef] [PubMed]
- Assmus, B.; Hutzler, P.; Kirchhof, G.; Amann, R.; Lawrence, J.R.; Hartmann, A. In Situ Localization of Azospirillum brasilense in the Rhizosphere of Wheat with Fluorescently Labeled, rRNA-Targeted Oligonucleotide Probes and Scanning Confocal Laser Microscopy. Appl. Environ. Microbiol. 1995, 61, 1013–1019. [Google Scholar] [CrossRef] [Green Version]
- Chelius, M.K.; Triplett, E.W. Immunolocalization of Dinitrogenase Reductase Produced by Klebsiella pneumoniae in Association with Zea mays L. Appl. Environ. Microbiol. 2000, 66, 783–787. [Google Scholar] [CrossRef] [Green Version]
- Hurek, T.; Reinhold-Hurek, B.; Van Montagu, M.; Kellenberger, E. Root Colonization and Systemic Spreading of Azoarcus sp. Strain BH72 in Grasses. J. Bacteriol. 1994, 176, 1913–1923. [Google Scholar] [CrossRef] [Green Version]
- Fan, B.; Borriss, R.; Bleiss, W.; Wu, X. Gram-Positive Rhizobacterium Bacillus amyloliquefaciens FZB42 Colonizes Three Types of Plants in Different Patterns. J. Microbiol. 2012, 50, 38–44. [Google Scholar] [CrossRef]
- Tringe, S.G.; Hugenholtz, P. A Renaissance for the Pioneering 16S rRNA Gene. Curr. Opin. Microbiol. 2008, 11, 442–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. A Communal Catalogue Reveals Earth’s Multiscale Microbial Diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef]
- Baker, G.C.; Smith, J.J.; Cowan, D.A. Review and Re-Analysis of Domain-Specific 16S Primers. J. Microbiol. Methods 2003, 55, 541–555. [Google Scholar] [CrossRef] [Green Version]
- Tkacz, A.; Hortala, M.; Poole, P.S. Absolute Quantitation of Microbiota Abundance in Environmental Samples. Microbiome 2018, 6, 110. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Zhang, X.; Qin, Y.; Liu, Y.-X.; Zhang, J.; Zhang, N.; Wu, K.; Qu, B.; He, Z.; Wang, X.; et al. Host-Associated Quantitative Abundance Profiling Reveals the Microbial Load Variation of Root Microbiome. Plant Commun. 2020, 1, 100003. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, D.S.; Pramoj Na Ayutthaya, P.; Strauß, A.; Shirsekar, G.; Lo, W.-S.; Lahaye, T.; Weigel, D. Host-Associated Microbe PCR (hamPCR) Enables Convenient Measurement of Both Microbial Load and Community Composition. Elife 2021, 10, e66186. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Morrison, M.; Yu, Z. Evaluation of Different Partial 16S rRNA Gene Sequence Regions for Phylogenetic Analysis of Microbiomes. J. Microbiol. Methods 2011, 84, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.-Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S rRNA Gene Sequencing for Species and Strain-Level Microbiome Analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, Y.; Komiya, S.; Yasumizu, Y.; Yasuoka, Y.; Mizushima, K.; Takagi, T.; Kryukov, K.; Fukuda, A.; Morimoto, Y.; Naito, Y.; et al. Full-Length 16S rRNA Gene Amplicon Analysis of Human Gut Microbiota Using MinIONTM Nanopore Sequencing Confers Species-Level Resolution. BMC Microbiol. 2021, 21, 35. [Google Scholar] [CrossRef]
- Callahan, B.J.; Wong, J.; Heiner, C.; Oh, S.; Theriot, C.M.; Gulati, A.S.; McGill, S.K.; Dougherty, M.K. High-Throughput Amplicon Sequencing of the Full-Length 16S rRNA Gene with Single-Nucleotide Resolution. Nucleic Acids Res. 2019, 47, e103. [Google Scholar] [CrossRef] [Green Version]
- Janda, J.M.; Abbott, S.L. 16S rRNA Gene Sequencing for Bacterial Identification in the Diagnostic Laboratory: Pluses, Perils, and Pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef] [Green Version]
- Mignard, S.; Flandrois, J.P. 16S rRNA Sequencing in Routine Bacterial Identification: A 30-Month Experiment. J. Microbiol. Methods 2006, 67, 574–581. [Google Scholar] [CrossRef]
- Yang, B.; Wang, Y.; Qian, P.-Y. Sensitivity and Correlation of Hypervariable Regions in 16S rRNA Genes in Phylogenetic Analysis. BMC Bioinform. 2016, 17, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, A.Y.; Oberdorf, W.E.; Nossa, C.W.; Agarwal, A.; Chokshi, P.; Gerz, E.A.; Jin, Z.; Lee, P.; Yang, L.; Poles, M.; et al. Diversity of 16S rRNA Genes within Individual Prokaryotic Genomes. Appl. Environ. Microbiol. 2010, 76, 3886–3897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maroniche, G.A.; García, J.E.; Salcedo, F.; Creus, C.M. Molecular Identification of Azospirillum Spp.: Limitations of 16S rRNA and Qualities of rpoD as Genetic Markers. Microbiol. Res. 2017, 195, 1–10. [Google Scholar] [CrossRef]
- Rilling, J.I.; Acuña, J.J.; Nannipieri, P.; Cassan, F.; Maruyama, F.; Jorquera, M.A. Current opinion and perspectives on the methods for tracking and monitoring plant growth–promoting bacteria. Soil Biol. Biochem. 2019, 130, 205–219. [Google Scholar] [CrossRef]
- Ruppel, S.; Rühlmann, J.; Merbach, W. Quantification and localization of bacteria in plant tissues using quantitative real-time PCR and online emission fingerprinting. Plant Soil 2006, 286, 21–35. [Google Scholar] [CrossRef]
- Couillerot, O.; Poirier, M.-A.; Prigent-Combaret, C.; Mavingui, P.; Caballero-Mellado, J.; Moënne-Loccoz, Y. Assessment of SCAR Markers to Design Real-Time PCR Primers for Rhizosphere Quantification of Azospirillum brasilense Phytostimulatory Inoculants of Maize. J. Appl. Microbiol. 2010, 109, 528–538. [Google Scholar] [CrossRef]
- Pereira, T.P.; do Amaral, F.P.; Dall’Asta, P.; Brod, F.C.A.; Arisi, A.C.M. Real-Time PCR Quantification of the Plant Growth Promoting Bacteria Herbaspirillum seropedicae Strain SmR1 in Maize Roots. Mol. Biotechnol. 2014, 56, 660–670. [Google Scholar] [CrossRef]
- Stets, M.I.; Alqueres, S.M.C.; Souza, E.M.; Pedrosa, F.d.O.; Schmid, M.; Hartmann, A.; Cruz, L.M. Quantification of Azospirillum brasilense FP2 Bacteria in Wheat Roots by Strain-Specific Quantitative PCR. Appl. Environ. Microbiol. 2015, 81, 6700–6709. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Gao, X.; Wang, S.; Zhu, C.; Li, R.; Shen, Q. Application of Bacillus velezensis NJAU-Z9 Enhanced Plant Growth Associated with Efficient Rhizospheric Colonization Monitored by qPCR with Primers Designed from the Whole Genome Sequence. Curr. Microbiol. 2018, 75, 1574–1583. [Google Scholar] [CrossRef] [Green Version]
- Mendis, H.C.; Thomas, V.P.; Schwientek, P.; Salamzade, R.; Chien, J.-T.; Waidyarathne, P.; Kloepper, J.; De La Fuente, L. Strain-Specific Quantification of Root Colonization by Plant Growth Promoting Rhizobacteria Bacillus firmus I-1582 and Bacillus amyloliquefaciens QST713 in Non-Sterile Soil and Field Conditions. PLoS ONE 2018, 13, e0193119. [Google Scholar] [CrossRef]
- Urrea-Valencia, S.; Etto, R.M.; Takahashi, W.Y.; Caires, E.F.; Bini, A.R.; Ayub, R.A.; Stets, M.I.; Cruz, L.M.; Galvao, C.W. Detection of Azospirillum brasilense by qPCR throughout a maize field trial. Appl. Soil Ecol. 2021, 160, 103849. [Google Scholar] [CrossRef]
- Kaminsky, L.M.; Bell, T.H. Novel primers for quantification of Priestia megaterium populations in soil using qPCR. Appl. Soil Ecol. 2022, 180, 104628. [Google Scholar] [CrossRef]
- Gilbert, S.; Xu, J.; Acosta, K.; Poulev, A.; Lebeis, S.; Lam, E. Bacterial Production of Indole Related Compounds Reveals Their Role in Association Between Duckweeds and Endophytes. Front. Chem. 2018, 6, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, S.; Poulev, A.; Chrisler, W.; Acosta, K.; Orr, G.; Lebeis, S.; Lam, E. Auxin-Producing Bacteria from Duckweeds Have Different Colonization Patterns and Effects on Plant Morphology. Plants 2022, 11, 721. [Google Scholar] [CrossRef] [PubMed]
- Steenhoudt, O.; Vanderleyden, J. Azospirillum, a Free-Living Nitrogen-Fixing Bacterium Closely Associated with Grasses: Genetic, Biochemical and Ecological Aspects. FEMS Microbiol. Rev. 2000, 24, 487–506. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, N.d.S.; dos Santos Ferreira, N.; Anna, F.H.S.; Reis, V.M.; Ambrosini, A.; Volpiano, C.G.; Rothballer, M.; Schwab, S.; Baura, V.A.; Balsanelli, E.; et al. Genome-Based Reclassification of Azospirillum brasilense Sp245 as the Type Strain of Azospirillum baldaniorum Sp. Nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 6203–6212. [Google Scholar] [CrossRef]
- Borisjuk, N.; Chu, P.; Gutierrez, R.; Zhang, H.; Acosta, K.; Friesen, N.; Sree, K.S.; Garcia, C.; Appenroth, K.J.; Lam, E. Assessment, Validation and Deployment Strategy of a Two-Barcode Protocol for Facile Genotyping of Duckweed Species. Plant Biol. 2015, 17 (Suppl. S1), 42–49. [Google Scholar] [CrossRef] [PubMed]
- García-Martínez, J.; Acinas, S.G.; Antón, A.I.; Rodríguez-Valera, F. Use of the 16S–23S Ribosomal Genes Spacer Region in Studies of Prokaryotic Diversity. J. Microbiol. Methods 1999, 36, 55–64. [Google Scholar] [CrossRef]
- Gürtler, V.; Stanisich, V.A. New Approaches to Typing and Identification of Bacteria Using the 16S-23S rDNA Spacer Region. Microbiology 1996, 142, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Michael, T.P.; Bryant, D.; Gutierrez, R.; Borisjuk, N.; Chu, P.; Zhang, H.; Xia, J.; Zhou, J.; Peng, H.; El Baidouri, M.; et al. Comprehensive Definition of Genome Features in Spirodela polyrhiza by High-Depth Physical Mapping and Short-Read DNA Sequencing Strategies. Plant J. 2017, 89, 617–635. [Google Scholar] [CrossRef] [PubMed]
- Hoang, P.N.T.; Michael, T.P.; Gilbert, S.; Chu, P.; Motley, S.T.; Appenroth, K.J.; Schubert, I.; Lam, E. Generating a High-Confidence Reference Genome Map of the Greater Duckweed by Integration of Cytogenomic, Optical Mapping, and Oxford Nanopore Technologies. Plant J. 2018, 96, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, P.G.; Miller, A.J.; Hirsch, P.R. Are Root Exudates More Important than Other Sources of Rhizodeposits in Structuring Rhizosphere Bacterial Communities? FEMS Microbiol. Ecol. 2010, 72, 313–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, J.H.; Orr, D.R. Distribution of Epiphytic Bacteria on Freshwater Plants. J. Ecol. 1986, 74, 155–165. [Google Scholar] [CrossRef]
- Borisjuk, N.; Peterson, A.A.; Lv, J.; Qu, G.; Luo, Q.; Shi, L.; Chen, G.; Kishchenko, O.; Zhou, Y.; Shi, J. Structural and Biochemical Properties of Duckweed Surface Cuticle. Front. Chem. 2018, 6, 317. [Google Scholar] [CrossRef] [PubMed]
- Ware, A.; Jones, D.H.; Flis, P.; Smith, K.; Kümpers, B.; Yant, L.; Atkinson, J.A.; Wells, D.M.; Bishopp, A. Duckweed Roots Are Dispensable and Are on a Trajectory toward Vestigiality. bioRxiv 2022, 1, 475062. [Google Scholar] [CrossRef]
- Chaintreuil, C.; Giraud, E.; Prin, Y.; Lorquin, J.; Bâ, A.; Gillis, M.; de Lajudie, P.; Dreyfus, B. Photosynthetic Bradyrhizobia Are Natural Endophytes of the African Wild Rice Oryza breviligulata. Appl. Environ. Microbiol. 2000, 66, 5437–5447. [Google Scholar] [CrossRef] [Green Version]
- Gopalaswamy, G.; Kannaiyan, S.; O’Callaghan, K.J.; Davey, M.R.; Cocking, E.C. The Xylem of Rice (Oryza Sativa) Is Colonized by Azorhizobium caulinodans. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2000, 267, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Baldani, V.L.D.; Alvarez, M.A.; Baldani, J.I.; Döbereiner, J. Establishment of Inoculated Azospirillum spp. in the Rhizosphere and in Roots of Field Grown Wheat and Sorghum. Plant Soil 1986, 90, 35–46. [Google Scholar] [CrossRef]
- Tarrand, J.J.; Krieg, N.R.; Döbereiner, J. A Taxonomic Study of the Spirillum lipoferum Group, with Descriptions of a New Genus, Azospirillum Gen. Nov. and Two Species, Azospirillum lipoferum (Beijerinck) Comb. Nov. and Azospirillum brasilense Sp. Nov. Can. J. Microbiol. 1978, 24, 967–980. [Google Scholar] [CrossRef]
- Gafny, R.; Okon, Y.; Kapulnik, Y.; Fischer, M. Adsorption of Azospirillum brasilense to Corn Roots. Soil Biol. Biochem. 1986, 18, 69–75. [Google Scholar] [CrossRef]
- Mori, K.; Toyama, T.; Sei, K. Surfactants Degrading Activities in the Rhizosphere of Giant Duckweed (Spirodela polyrhiza). Jpn. J. Water Treat. Biol. 2005, 41, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Ishizawa, H.; Kuroda, M.; Inoue, K.; Inoue, D.; Morikawa, M.; Ike, M. Colonization and Competition Dynamics of Plant Growth-Promoting/Inhibiting Bacteria in the Phytosphere of the Duckweed Lemna minor. Microb. Ecol. 2019, 77, 440–450. [Google Scholar] [CrossRef]
- Ishizawa, H.; Kuroda, M.; Inoue, D.; Morikawa, M.; Ike, M. Community Dynamics of Duckweed-Associated Bacteria upon Inoculation of Plant Growth-Promoting Bacteria. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef] [PubMed]
- Haro, C.; Anguita-Maeso, M.; Metsis, M.; Navas-Cortés, J.A.; Landa, B.B. Evaluation of Established Methods for DNA Extraction and Primer Pairs Targeting 16S rRNA Gene for Bacterial Microbiota Profiling of Olive Xylem Sap. Front. Plant Sci. 2021, 12, 640829. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Kittelmann, S.; Miri, V.H.; Zethof, M.; Noel, S.J.; Waghorn, G.C.; Janssen, P.H. Effect of DNA Extraction Methods and Sampling Techniques on the Apparent Structure of Cow and Sheep Rumen Microbial Communities. PLoS ONE 2013, 8, e74787. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.W.; Martin, J.C.; Scott, P.; Parkhill, J.; Flint, H.J.; Scott, K.P. 16S rRNA Gene-Based Profiling of the Human Infant Gut Microbiota Is Strongly Influenced by Sample Processing and PCR Primer Choice. Microbiome 2015, 3, 26. [Google Scholar] [CrossRef] [Green Version]
- Maukonen, J.; Simões, C.; Saarela, M. The Currently Used Commercial DNA-Extraction Methods Give Different Results of Clostridial and Actinobacterial Populations Derived from Human Fecal Samples. FEMS Microbiol. Ecol. 2012, 79, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Salonen, A.; Nikkilä, J.; Jalanka-Tuovinen, J.; Immonen, O.; Rajilić-Stojanović, M.; Kekkonen, R.A.; Palva, A.; de Vos, W.M. Comparative Analysis of Fecal DNA Extraction Methods with Phylogenetic Microarray: Effective Recovery of Bacterial and Archaeal DNA Using Mechanical Cell Lysis. J. Microbiol. Methods 2010, 81, 127–134. [Google Scholar] [CrossRef]
- Leite, G.M.; Magan, N.; Medina, Á. Comparison of Different Bead-Beating RNA Extraction Strategies: An Optimized Method for Filamentous Fungi. J. Microbiol. Methods 2012, 88, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.N.; Bryant, J.E.; Madsen, E.L.; Ghiorse, W.C. Evaluation and Optimization of DNA Extraction and Purification Procedures for Soil and Sediment Samples. Appl. Environ. Microbiol. 1999, 65, 4715–4724. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Cohen, D.B.; Ravel, J.; Abdo, Z.; Forney, L.J. Evaluation of Methods for the Extraction and Purification of DNA from the Human Microbiome. PLoS ONE 2012, 7, e33865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollock, J.; Glendinning, L.; Wisedchanwet, T.; Watson, M. The Madness of Microbiome: Attempting to Find Consensus “Best Practice” for 16S Microbiome Studies. Appl. Environ. Microbiol. 2018, 84, e02627-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouhy, F.; Clooney, A.G.; Stanton, C.; Claesson, M.J.; Cotter, P.D. 16S rRNA Gene Sequencing of Mock Microbial Populations- Impact of DNA Extraction Method, Primer Choice and Sequencing Platform. BMC Microbiol. 2016, 16, 123. [Google Scholar] [CrossRef]
- Brooks, J.P.; Edwards, D.J.; Harwich, M.D., Jr.; Rivera, M.C.; Fettweis, J.M.; Serrano, M.G.; Reris, R.A.; Sheth, N.U.; Huang, B.; Girerd, P.; et al. The Truth about Metagenomics: Quantifying and Counteracting Bias in 16S rRNA Studies. BMC Microbiol. 2015, 15, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, M.M.; Triplett, E.W. Automated Approach for Ribosomal Intergenic Spacer Analysis of Microbial Diversity and Its Application to Freshwater Bacterial Communities. Appl. Environ. Microbiol. 1999, 65, 4630–4636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodenhausen, N.; Bortfeld-Miller, M.; Ackermann, M.; Vorholt, J.A. A Synthetic Community Approach Reveals Plant Genotypes Affecting the Phyllosphere Microbiota. PLoS Genet. 2014, 10, e1004283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowker, D.W.; Duffield, A.N.; Denny, P. Methods for the Isolation, Sterilization and Cultivation of Lemnaceae. Freshw. Biol. 1980, 10, 385–388. [Google Scholar] [CrossRef]
- Huang, W.; Gilbert, S.; Poulev, A.; Acosta, K.; Lebeis, S.; Long, C.; Lam, E. Host-Specific and Tissue-Dependent Orchestration of Microbiome Community Structure in Traditional Rice Paddy Ecosystems. Plant Soil 2020, 452, 379–395. [Google Scholar] [CrossRef]
- Bodenhausen, N.; Deslandes-Hérold, G.; Waelchli, J.; Held, A.; van der Heijden, M.G.A.; Schlaeppi, K. Relative qPCR to Quantify Colonization of Plant Roots by Arbuscular Mycorrhizal Fungi. Mycorrhiza 2021, 31, 137–148. [Google Scholar] [CrossRef]
- Garrido-Oter, R.; Nakano, R.T.; Dombrowski, N.; Ma, K.-W.; AgBiome Team; McHardy, A.C.; Schulze-Lefert, P. Modular Traits of the Rhizobiales Root Microbiota and Their Evolutionary Relationship with Symbiotic Rhizobia. Cell Host Microbe 2018, 24, 155–167.e5. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, S.; Phillippy, A.; Delcher, A.L.; Smoot, M.; Shumway, M.; Antonescu, C.; Salzberg, S.L. Versatile and Open Software for Comparing Large Genomes. Genome Biol. 2004, 5, R12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The Plant Microbiota: Systems-Level Insights and Perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome Interactions: From Community Assembly to Plant Health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Salas Gonzalez, I.; Mittelviefhaus, M.; Clingenpeel, S.; Herrera Paredes, S.; Miao, J.; Wang, K.; Devescovi, G.; Stillman, K.; Monteiro, F.; et al. Genomic Features of Bacterial Adaptation to Plants. Nat. Genet. 2017, 50, 138–150. [Google Scholar] [CrossRef] [Green Version]
- Hardoim, P.R.; van Overbeek, L.S.; van Elsas, J.D. Properties of Bacterial Endophytes and Their Proposed Role in Plant Growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef]
- Cole, B.J.; Feltcher, M.E.; Waters, R.J.; Wetmore, K.M.; Mucyn, T.S.; Ryan, E.M.; Wang, G.; Ul-Hasan, S.; McDonald, M.; Yoshikuni, Y.; et al. Genome-Wide Identification of Bacterial Plant Colonization Genes. PLoS Biol. 2017, 15, e2002860. [Google Scholar] [CrossRef]
- Ishizawa, H.; Kuroda, M.; Inoue, D.; Ike, M. Genome-Wide Identification of Bacterial Colonization and Fitness Determinants on the Floating Macrophyte, Duckweed. Commun. Biol. 2022, 5, 68. [Google Scholar] [CrossRef]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-Acetic Acid in Plant–microbe Interactions. Antonie Van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Tzipilevich, E.; Russ, D.; Dangl, J.L.; Benfey, P.N. Plant Immune System Activation Is Necessary for Efficient Root Colonization by Auxin-Secreting Beneficial Bacteria. Cell Host Microbe 2021, 29, 1507–1520.e4. [Google Scholar] [CrossRef]
- Ishizawa, H.; Kuroda, M.; Morikawa, M.; Ike, M. Evaluation of Environmental Bacterial Communities as a Factor Affecting the Growth of Duckweed Lemna Minor. Biotechnol. Biofuels 2017, 10, 62. [Google Scholar] [CrossRef]
- Vorholt, J.A.; Vogel, C.; Carlström, C.I.; Müller, D.B. Establishing Causality: Opportunities of Synthetic Communities for Plant Microbiome Research. Cell Host Microbe 2017, 22, 142–155. [Google Scholar] [CrossRef] [PubMed]
- Carper, D.L.; Lawrence, T.J.; Carrell, A.A.; Pelletier, D.A.; Weston, D.J. DISCo-Microbe: Design of an Identifiable Synthetic Community of Microbes. PeerJ 2020, 8, e8534. [Google Scholar] [CrossRef] [PubMed]
- Miché, L.; Balandreau, J. Effects of Rice Seed Surface Sterilization with Hypochlorite on Inoculated Burkholderia vietnamiensis. Appl. Environ. Microbiol. 2001, 67, 3046–3052. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de Novo Short Read Assembly Using de Bruijn Graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Butler, J.; MacCallum, I.; Kleber, M.; Shlyakhter, I.A.; Belmonte, M.K.; Lander, E.S.; Nusbaum, C.; Jaffe, D.B. ALLPATHS: De Novo Assembly of Whole-Genome Shotgun Microreads. Genome Res. 2008, 18, 810–820. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Crouse, J.; Amorese, D. Ethanol Precipitation: Ammonium Acetate as an Alternative to Sodium Acetate. Focus 1996, 19, 17–20. [Google Scholar]
- Van Hoeck, A.; Horemans, N.; Monsieurs, P.; Cao, H.X.; Vandenhove, H.; Blust, R. The First Draft Genome of the Aquatic Model Plant Lemna minor Opens the Route for Future Stress Physiology Research and Biotechnological Applications. Biotechnol. Biofuels 2015, 8, 188. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Laing, C.; Buchanan, C.; Taboada, E.N.; Zhang, Y.; Kropinski, A.; Villegas, A.; Thomas, J.E.; Gannon, V.P.J. Pan-Genome Sequence Analysis Using Panseq: An Online Tool for the Rapid Analysis of Core and Accessory Genomic Regions. BMC Bioinform. 2010, 11, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an Enhanced Web Interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaumeil, P.-A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A Toolkit to Classify Genomes with the Genome Taxonomy Database. Bioinformatics 2019, 36, 1925–1927. [Google Scholar] [CrossRef]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Staerfeldt, H.-H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and Rapid Annotation of Ribosomal RNA Genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A Flexible Suite of Utilities for Comparing Genomic Features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Stothard, P. The Sequence Manipulation Suite: JavaScript Programs for Analyzing and Formatting Protein and DNA Sequences. Biotechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acosta, K.; Sorrels, S.; Chrisler, W.; Huang, W.; Gilbert, S.; Brinkman, T.; Michael, T.P.; Lebeis, S.L.; Lam, E. Optimization of Molecular Methods for Detecting Duckweed-Associated Bacteria. Plants 2023, 12, 872. https://doi.org/10.3390/plants12040872
Acosta K, Sorrels S, Chrisler W, Huang W, Gilbert S, Brinkman T, Michael TP, Lebeis SL, Lam E. Optimization of Molecular Methods for Detecting Duckweed-Associated Bacteria. Plants. 2023; 12(4):872. https://doi.org/10.3390/plants12040872
Chicago/Turabian StyleAcosta, Kenneth, Shawn Sorrels, William Chrisler, Weijuan Huang, Sarah Gilbert, Thomas Brinkman, Todd P. Michael, Sarah L. Lebeis, and Eric Lam. 2023. "Optimization of Molecular Methods for Detecting Duckweed-Associated Bacteria" Plants 12, no. 4: 872. https://doi.org/10.3390/plants12040872