The Cytoskeleton and Its Role in Determining Cellulose Microfibril Angle in Secondary Cell Walls of Woody Tree Species


 and
and 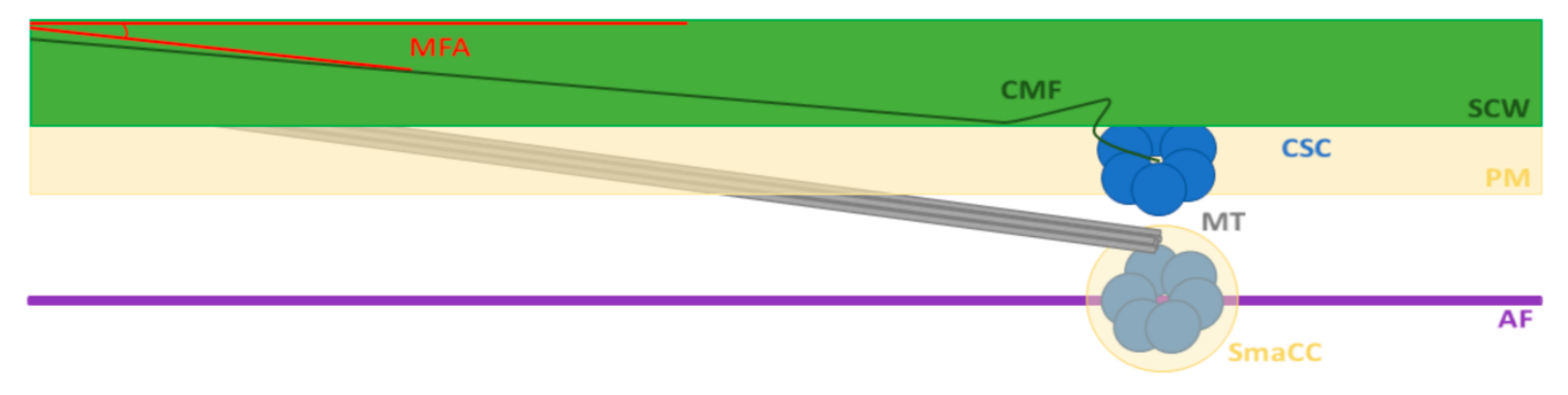{kind=link}
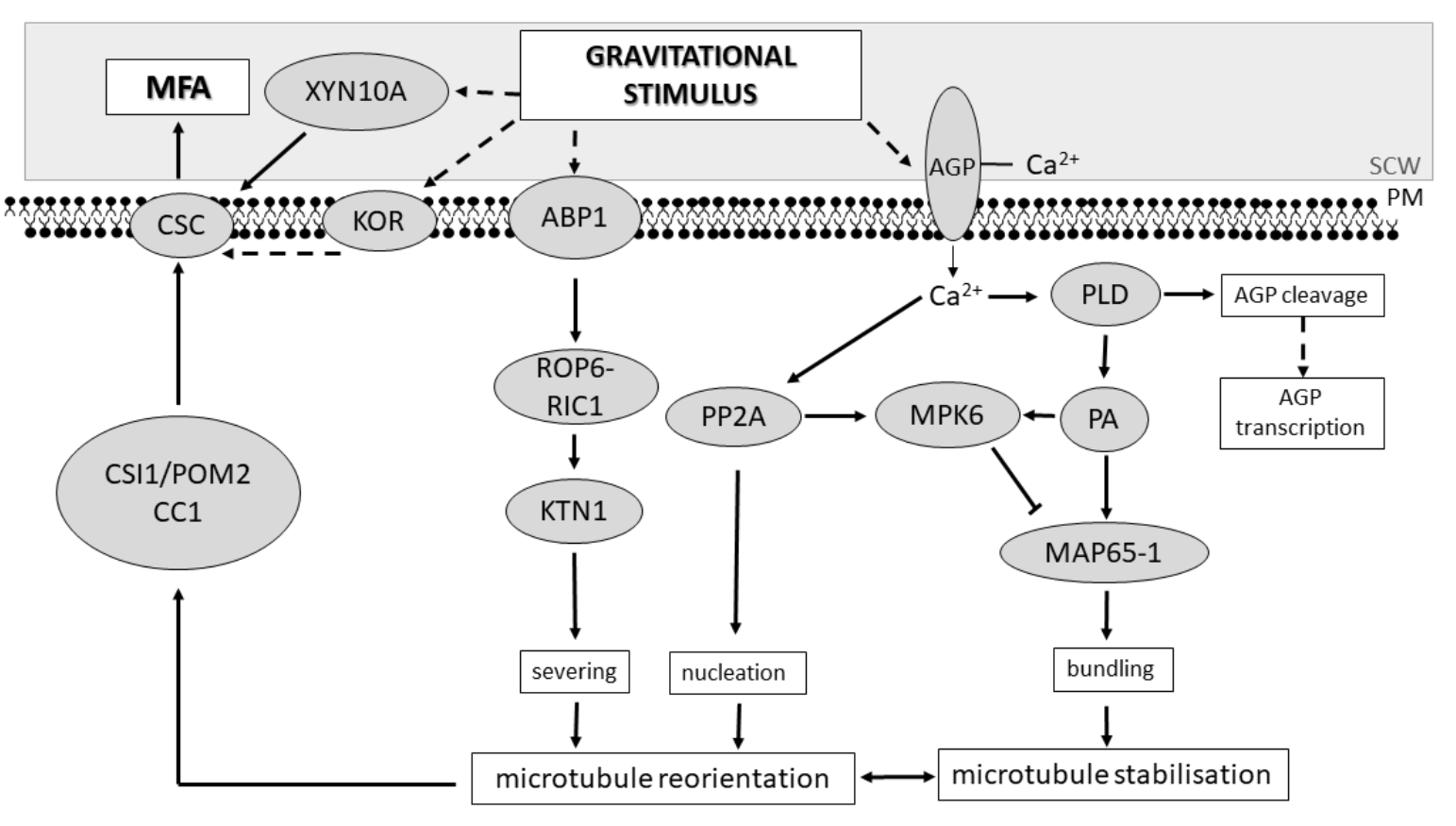{kind=link}
Abstract
:1. Introduction
2. MFA as a Key Feature of SCW Formation
3. Cellulose Properties and the CSC
4. Cytoskeleton Roles in SCW Biosynthesis
4.1. Microtubules Guide the CSC and Play a Role in SCW Patterning
4.2. Actin Filaments and Microtubules Act Together to Deliver CSC to the Plasma Membrane
5. Molecular Control of MFA
5.1. Arabinogalactans
5.2. Tubulins
5.3. Other Cell Wall-Related Genes
5.4. A Molecular Model for MFA Alterations in Response to Gravitational Stimulus
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Carlquist, S. How wood evolves: A new synthesis. Botany 2012, 90, 901–940. [Google Scholar] [CrossRef] [Green Version]
- Herth, W. Plasma membrane rosettes involved in localised wall thickening during xylem vessel formation of Lepidium sativum L. Planta 1985, 164, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Lampugnani, E.R.; Khan, G.A.; Somssich, M.; Persson, S. Building a plant cell wall at a glance. J. Cell Sci. 2018, 131, jcs207373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFarlane, H.E.; Doring, A.; Persson, S. The cell biology of cellulose synthesis. Annu. Rev. Plant Biol. 2014, 65, 69–94. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, S.; Somssich, M.; Nakata, M.T.; Unda, F.; Atsuzawa, K.; Kaneko, Y.; Wang, T.; Bagman, A.M.; Gaudinier, A.; Yoshida, K.; et al. Complete substitution of a secondary cell wall with a primary cell wall in Arabidopsis. Nat. Plants 2018, 4, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.Q.; Cui, D.T.; Ye, Z.H. Secondary cell wall biosynthesis. New Phytol. 2019, 221, 1703–1723. [Google Scholar] [CrossRef] [Green Version]
- Almeras, T.; Clair, B. Critical review on the mechanisms of maturation stress generation in trees. J. R. Soc. Interface 2016, 13, 20160550. [Google Scholar] [CrossRef] [Green Version]
- Meents, M.J.; Watanabe, Y.; Samuels, A.L. The cell biology of secondary cell wall biosynthesis. Ann. Bot. 2018, 121, 1107–1125. [Google Scholar] [CrossRef] [Green Version]
- Baskin, T.I. Anisotropic expansion of the plant cell wall. Annu. Rev. Cell Dev. Biol. 2005, 21, 203–222. [Google Scholar] [CrossRef] [Green Version]
- Green, P.B. Mechanism for plant cellular morphogenesis. Science 1962, 138, 1404–1405. [Google Scholar] [CrossRef]
- Chan, J. Microtubule and cellulose microfibril orientation during plant cell and organ growth. J. Microsc. 2012, 247, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T. Microfibrillar orientation of radially enlarged cells of coumarin- and colchicine-treated pine seedlings. Plant Cell Physiol. 1976, 17, 385–398. [Google Scholar]
- Hardham, A.R.; Gunning, B.E.S. Interpolation of microtubules into cortical arrays during cell elongation and differentiation in roots of Azolla pinnata. J. Cell Sci. 1979, 37, 411–442. [Google Scholar] [PubMed]
- Arioli, T.; Peng, L.C.; Betzner, A.S.; Burn, J.; Wittke, W.; Herth, W.; Camilleri, C.; Hofte, H.; Plazinski, J.; Birch, R.; et al. Molecular analysis of cellulose biosynthesis in Arabidopsis. Science 1998, 279, 717–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagard, M.; Desnos, T.; Desprez, T.; Goubet, F.; Refregier, G.; Mouille, G.; McCann, M.; Rayon, C.; Vernhettes, S.; Hofte, H. PROCUSTE1 encodes a cellulose synthase required for normal cell elongation specifically in roots and dark-grown hypocotyls of Arabidopsis. Plant Cell 2000, 12, 2409–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, E.T.; Graham, B.P.; Stiff, M.R.; Osborne, J.A.; Haigler, C.H. Cultures of Gossypium barbadense cotton ovules offer insights into the microtubule-mediated control of fiber cell expansion. Planta 2019, 249, 1551–1563. [Google Scholar] [CrossRef]
- Baskin, T.I. On the alignment of cellulose microfibrils by cortical microtubules: A review and a model. Protoplasma 2001, 215, 150–171. [Google Scholar] [CrossRef]
- Chan, J.; Eder, M.; Crowell, E.F.; Hampson, J.; Calder, G.; Lloyd, C. Microtubules and CESA tracks at the inner epidermal wall align independently of those on the outer wall of light-grown Arabidopsis hypocotyls. J. Cell Sci. 2011, 124, 1088–1094. [Google Scholar] [CrossRef] [Green Version]
- Crowell, E.F.; Timpano, H.; Desprez, T.; Franssen-Verheijen, T.; Emons, A.-M.; Hoefte, H.; Vernhettes, S. Differential regulation of cellulose orientation at the inner and outer face of epidermal cells in the Arabidopsis hypocotyl. Plant Cell 2011, 23, 2592–2605. [Google Scholar] [CrossRef] [Green Version]
- Preston, R.D. The organization of the cell wall of the conifer tracheid. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1934, 224, 131–145. [Google Scholar]
- Mansfield, S.D.; Parish, R.; Di Lucca, C.M.; Goudie, J.; Kang, K.Y.; Ott, P. Revisiting the transition between juvenile and mature wood: A comparison of fibre length, microfibril angle and relative wood density in Lodgepole pine. Holzforschung 2009, 63, 449–456. [Google Scholar] [CrossRef]
- Donaldson, L. Microfibril angle: Measurement, variation and relationships—A review. IAWA J. 2008, 29, 345–386. [Google Scholar] [CrossRef]
- Barnett, J.R.; Bonham, V.A. Cellulose microfibril angle in the cell wall of wood fibres. Biol. Rev. 2004, 79, 461–472. [Google Scholar] [CrossRef]
- Matsumura, J.; Butterfield, B.G. Microfibril angles in the root wood of Pinus radiata and Pinus nigra. IAWA J. 2001, 22, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Myszewski, J.H.; Bridgwater, F.E.; Lowe, W.J.; Byram, T.D.; Megraw, R.A. Genetic variation in the microfibril angle of Loblolly pine from two test sites. South. J. Appl. For. 2004, 28, 196–204. [Google Scholar]
- Evans, R.; Stringer, S.; Kibblewhite, R.P. Variation of microfibril angle, density and fibre orientation in twenty-nine Eucalyptus nitens trees. Appita J. 2000, 53, 450–457. [Google Scholar]
- Evans, R.; Ilic, J. Rapid prediction of wood stiffness from microfibril, angle and density. For. Prod. J. 2001, 51, 53–57. [Google Scholar]
- Cramer, S.; Kretschmann, D.; Lakes, R.; Schmidt, T. Earlywood and latewood elastic properties in Loblolly pine. Holzforschung 2005, 59, 531–538. [Google Scholar] [CrossRef]
- Hein, P.R.G.; Lima, J.T. Relationships between microfibril angle, modulus of elasticity and compressive strength in Eucalyptus wood. Maderas Cienc. Tecnol. 2012, 14, 267–274. [Google Scholar]
- Burgert, I. Exploring the micromechanical design of plant cell walls. Am. J. Bot. 2006, 93, 1391–1401. [Google Scholar] [CrossRef]
- Ishikura, Y. Structural and bending properties of juvenile and mature softwood. Cell Chem. Technol. 2017, 51, 879–887. [Google Scholar]
- Wessels, C.B.; Crafford, P.L.; Du Toit, B.; Grahn, T.; Johansson, M.; Lundqvist, S.O.; Sall, H.; Seifert, T. Variation in physical and mechanical properties from three drought tolerant Eucalyptus species grown on the dry west coast of Southern Africa. Eur. J. Wood Wood Prod. 2016, 74, 563–575. [Google Scholar] [CrossRef]
- Sattler, D.F.; Comeau, P.G.; Achim, A. Within-tree patterns of wood stiffness for white spruce (Picea glauca) and trembling aspen (Populus tremuloides). Can. J. For. Res. 2014, 44, 162–171. [Google Scholar] [CrossRef]
- Moore, J.R.; Cown, D.J.; McKinley, R.B. Modelling microfibril angle variation in New Zealand-grown Radiata pine. N. Z. J. For. Sci. 2014, 44, 25. [Google Scholar] [CrossRef] [Green Version]
- Bastien, R.; Bohr, T.; Moulia, B.; Douady, S. Unifying model of shoot gravitropism reveals proprioception as a central feature of posture control in plants. Proc. Natl. Acad. Sci. USA 2013, 110, 755–760. [Google Scholar] [CrossRef] [Green Version]
- Groover, A. Gravitropisms and reaction woods of forest trees—Evolution, functions and mechanisms. New Phytol. 2016, 211, 790–802. [Google Scholar] [CrossRef]
- Bamber, R.K. A general theory for the origin of growth stresses in reaction wood: How trees stay upright. IAWA J. 2001, 22, 205–212. [Google Scholar] [CrossRef]
- Deng, Q.; Li, S.; Chen, Y.P. Mechanical properties and failure mechanism of wood cell wall layers. Comput. Mater. Sci. 2012, 62, 221–226. [Google Scholar] [CrossRef]
- Wang, H.K.; Yu, Z.X.; Zhang, X.X.; Ren, D.; Yu, Y. The combined effects of initial microfibrillar angle and moisture contents on the tensile mechanical properties and angle alteration of wood foils during tension. Holzforschung 2017, 71, 491–497. [Google Scholar] [CrossRef]
- Spokevicius, A.; Southerton, S.G.; MacMillan, C.P.; Qiu, D.; Gan, S.; Tibbits, J.F.G.; Moran, G.F.; Bossinger, G. Beta-tubulin affects cellulose microfibril orientation in plant secondary fibre cell walls. Plant J. 2007, 51, 717–726. [Google Scholar] [CrossRef]
- Roignant, J.; Badel, E.; Leblanc-Fournier, N.; Brunel-Michac, N.; Ruelle, J.; Moulia, B.; Decourteix, M. Feeling stretched or compressed? The multiple mechanosensitive responses of wood formation to bending. Ann. Bot. 2018, 121, 1151–1161. [Google Scholar] [CrossRef]
- Nixon, B.T.; Mansouri, K.; Singh, A.; Du, J.; Davis, J.K.; Lee, J.G.; Slabaugh, E.; Vandavasi, V.G.; O’Neill, H.; Roberts, E.M.; et al. Comparative structural and computational analysis supports eighteen cellulose synthases in the plant cellulose synthesis complex. Sci. Rep. 2016, 6, 28696. [Google Scholar] [CrossRef] [Green Version]
- Persson, S.; Paredez, A.; Carroll, A.; Palsdottir, H.; Doblin, M.; Poindexter, P.; Khitrov, N.; Auer, M.; Somerville, C.R. Genetic evidence for three unique components in primary cell-wall cellulose synthase complexes in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 15566–15571. [Google Scholar] [CrossRef] [Green Version]
- Taylor, N.G.; Howells, R.M.; Huttly, A.K.; Vickers, K.; Turner, S.R. Interactions among three distinct CESA proteins essential for cellulose synthesis. Proc. Natl. Acad. Sci. USA 2003, 100, 1450–1455. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.; Specht, C.D. Understanding plant cellulose synthases through a comprehensive investigation of the cellulose synthase family sequences. Front. Plant Sci. 2011, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.; Mansoori, N.; Li, S.D.; Lei, L.; Vernhettes, S.; Visser, R.G.F.; Somerville, C.; Gu, Y.; Trindade, L.M. Complexes with mixed primary and secondary cellulose synthases are functional in Arabidopsis plants. Plant Physiol. 2012, 160, 726–737. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Dominguez, P.G.; Kumar, M.; Bygdell, J.; Miroshnichenko, S.; Sundberg, B.; Wingsle, G.; Niittyla, T. Cellulose synthase stoichiometry in aspen differs from Arabidopsis and Norway spruce. Plant Physiol. 2018, 177, 1096–1107. [Google Scholar] [CrossRef] [Green Version]
- Schneider, R.; Tang, L.; Lampugnani, E.R.; Barkwill, S.; Lathe, R.; Zhang, Y.; McFarlane, H.E.; Pesquet, E.; Niittyla, T.; Mansfield, S.D.; et al. Two complementary mechanisms underpin cell wall patterning during xylem vessel development. Plant Cell 2017, 29, 2433–2449. [Google Scholar] [CrossRef] [Green Version]
- Turner, S.; Kumar, M. Cellulose synthase complex organization and cellulose microfibril structure. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2018, 376, 20170048. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Mishra, L.; Carr, P.; Pilling, M.; Gardner, P.; Mansfield, S.D.; Turner, S. Exploiting CELLULOSE SYNTHASE (CESA) class specificity to probe cellulose microfibril biosynthesis. Plant Physiol. 2018, 177, 151–167. [Google Scholar] [CrossRef] [Green Version]
- Abe, H.; Funada, R.; Imaizumi, H.; Ohtani, J.; Fukazawa, K. Dynamic changes in the arrangement of cortical microtubules in conifer tracheids during differentiation. Planta 1995, 197, 418–421. [Google Scholar] [CrossRef]
- Chaffey, N.; Barlow, P.; Barnett, J. A cytoskeletal basis for wood formation in angiosperm trees: The involvement of microfilaments. Planta 2000, 210, 890–896. [Google Scholar] [CrossRef]
- Chaffey, N.; Barnett, J.; Barlow, P. A cytoskeletal basis for wood formation in angiosperm trees: The involvement of cortical microtubules. Planta 1999, 208, 19–30. [Google Scholar] [CrossRef]
- Chaffey, N.; Barlow, P.; Sundberg, B. Understanding the role of the cytoskeleton in wood formation in angiosperm trees: Hybrid aspen (Populus tremula × P-tremuloides) as the model species. Tree Physiol. 2002, 22, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Begum, S.; Furusawa, O.; Shibagaki, M.; Nakaba, S.; Yamagishi, Y.; Yoshimoto, J.; Rahman, M.H.; Sano, Y.; Funada, R. Localization of actin filaments and cortical microtubules in wood-forming tissues of conifers. IAWA J. 2019, 40, 703–720. [Google Scholar] [CrossRef]
- Lowe, J.; Li, H.; Downing, K.H.; Nogales, E. Refined structure of alpha beta-tubulin at 3.5 a resolution. J. Mol. Biol. 2001, 313, 1045–1057. [Google Scholar] [CrossRef]
- Heald, R.; Nogales, E. Microtubule dynamics. J. Cell Sci. 2002, 115, 3–4. [Google Scholar]
- Wasteneys, G.O. Microtubule organization in the green kingdom: Chaos or self-order? J. Cell Sci. 2002, 115, 1345–1354. [Google Scholar]
- Mirabet, V.; Krupinski, P.; Hamant, O.; Meyerowitz, E.M.; Jonsson, H.; Boudaoud, A. The self-organization of plant microtubules inside the cell volume yields their cortical localization, stable alignment, and sensitivity to external cues. PLoS Comput. Biol. 2018, 14, e1006011. [Google Scholar] [CrossRef]
- Baskin, T.I.; Beemster, G.T.S.; Judy-March, J.E.; Marga, F. Disorganization of cortical microtubules stimulates tangential expansion and reduces the uniformity of cellulose microfibril alignment among cells in the root of Arabidopsis. Plant Physiol. 2004, 135, 2279–2290. [Google Scholar] [CrossRef] [Green Version]
- Paredez, A.R.; Persson, S.; Ehrhardt, D.W.; Somerville, C.R. Genetic evidence that cellulose synthase activity influences microtubule cortical array organization. Plant Physiol. 2008, 147, 1723–1734. [Google Scholar] [CrossRef] [Green Version]
- Paredez, A.R.; Somerville, C.R.; Ehrhardt, D.W. Visualization of cellulose synthase demonstrates functional association with microtubules. Science 2006, 312, 1491–1495. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Meents, M.J.; McDonnell, L.M.; Barkwill, S.; Sampathkumar, A.; Cartwright, H.N.; Demura, T.; Ehrhardt, D.W.; Samuels, A.L.; Mansfield, S.D. Visualization of cellulose synthases in Arabidopsis secondary cell walls. Science 2015, 350, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Heath, I.B. A unified hypothesis for the role of membrane-bound enzyme complexes and microtubules in plant cell wall synthesis. J. Theor. Biol. 1974, 48, 445–449. [Google Scholar] [CrossRef]
- Giddings, T.H.; Staehelin, L.A. Microtubule-mediated control of microfibril deposition: A re-examination of the hypothesis. In The Cytoskeletal Basis of Plant Growth and Form; Lloyd, C.W., Ed.; Academic Press: London, UK, 1991; pp. 85–99. [Google Scholar]
- Zhong, R.Q.; Burk, D.H.; Morrison, W.H.; Ye, Z.H. A kinesin-like protein is essential for oriented deposition of cellulose microfibrils and cell wall strength. Plant Cell 2002, 14, 3101–3117. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.M.; Dixit, R. Single molecule analysis of the Arabidopsis FRA1 kinesin shows that it is a functional motor protein with unusually high processivity. Mol. Plant 2011, 4, 879–885. [Google Scholar] [CrossRef] [Green Version]
- Bringmann, M.; Li, E.Y.; Sampathkumar, A.; Kocabek, T.; Hauser, M.T.; Persson, S. POM-POM2/cellulose synthase interacting1 is essential for the functional association of cellulose synthase and microtubules in Arabidopsis. Plant Cell 2012, 24, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Lei, L.; Somerville, C.R.; Gu, Y. Cellulose synthase interactive protein 1 (CSI1) links microtubules and cellulose synthase complexes. Proc. Natl. Acad. Sci. USA 2012, 109, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Kesten, C.; Wallmann, A.; Schneider, R.; McFarlane, H.E.; Diehl, A.; Khan, G.A.; van Rossum, B.J.; Lampugnani, E.R.; Szymanski, W.G.; Cremer, N.; et al. The companion of cellulose synthase 1 confers salt tolerance through a tau-like mechanism in plants. Nat. Commun. 2019, 10, 857. [Google Scholar] [CrossRef] [Green Version]
- Woodley, M.; Mulvihill, A.; Fujita, M.; Wasteneys, G.O. Exploring microtubule-dependent cellulose-synthase-complex movement with high precision particle tracking. Plants 2018, 7, 53. [Google Scholar] [CrossRef] [Green Version]
- Oda, Y.; Fukuda, H. Initiation of cell wall pattern by a rho- and microtubule-driven symmetry breaking. Science 2012, 337, 1333–1336. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Nagashima, Y.; Wakazaki, M.; Sato, M.; Toyooka, K.; Fukuda, H.; Oda, Y. A rho-actin signaling pathway shapes cell wall boundaries in Arabidopsis xylem vessels. Nat. Commun. 2019, 10, 468. [Google Scholar] [CrossRef]
- Sasaki, T.; Fukuda, H.; Oda, Y. Cortical microtubule disordering1 is required for secondary cell wall patterning in xylem vessels. Plant Cell 2017, 29, 3123–3139. [Google Scholar] [CrossRef] [Green Version]
- Rajangam, A.S.; Kumar, M.; Aspeborg, H.; Guerriero, G.; Arvestad, L.; Pansri, P.; Brown, C.J.L.; Hober, S.; Blomqvist, K.; Divne, C.; et al. MAP20, a microtubule-associated protein in the secondary cell walls of hybrid aspen, is a target of the cellulose synthesis inhibitor 2,6-dichlorobenzonitrile. Plant Physiol. 2008, 148, 1283–1294. [Google Scholar] [CrossRef] [Green Version]
- Pesquet, E.; Korolev, A.V.; Calder, G.; Lloyd, C.W. The microtubule-associated protein AtMAP70-5 regulates secondary wall patterning in Arabidopsis wood cells. Curr. Biol. 2010, 20, 744–749. [Google Scholar] [CrossRef] [Green Version]
- Szymanski, D.; Staiger, C.J. The actin cytoskeleton: Functional arrays for cytoplasmic organization and cell shape control. Plant Physiol. 2018, 176, 106–118. [Google Scholar] [CrossRef]
- Sampathkumar, A.; Gutierrez, R.; McFarlane, H.E.; Bringmann, M.; Lindeboom, J.; Emons, A.M.; Samuels, L.; Ketelaar, T.; Ehrhardt, D.W.; Persson, S. Patterning and lifetime of plasma membrane-localized cellulose synthase is dependent on actin organization in Arabidopsis interphase cells. Plant Physiol. 2013, 162, 675–688. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, H.; Fukuda, H.; Shibaoka, H. Interrelation between the spatial disposition of actin-filaments and microtubules during the differentiation of tracheary elements in cultured zinnia cells. Protoplasma 1988, 143, 29–37. [Google Scholar] [CrossRef]
- Nebenfuhr, A.; Dixit, R. Kinesins and myosins: Molecular motors that coordinate cellular functions in plants. Annu. Rev. Plant Biol. 2018, 69, 329–361. [Google Scholar] [CrossRef]
- Wightman, R.; Turner, S.R. The roles of the cytoskeleton during cellulose deposition at the secondary cell wall. Plant J. 2008, 54, 794–805. [Google Scholar] [CrossRef]
- Gutierrez, R.; Lindeboom, J.J.; Paredez, A.R.; Emons, A.M.C.; Ehrhardt, D.W. Arabidopsis cortical microtubules position cellulose synthase delivery to the plasma membrane and interact with cellulose synthase trafficking compartments. Nat. Cell Biol. 2009, 11, 797. [Google Scholar] [CrossRef]
- Sampathkumar, A.; Lindeboom, J.J.; Debolt, S.; Gutierrez, R.; Ehrhardt, D.W.; Ketelaar, T.; Persson, S. Live cell imaging reveals structural associations between the actin and microtubule cytoskeleton in Arabidopsis. Plant Cell 2011, 23, 2302–2313. [Google Scholar] [CrossRef] [Green Version]
- Preuss, M.L.; Kovar, D.R.; Lee, Y.R.J.; Staiger, C.J.; Delmer, D.P.; Liu, B. A plant-specific kinesin binds to actin microfilaments and interacts with cortical microtubules in cotton fibers. Plant Physiol. 2004, 136, 3945–3955. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Qu, Z.; Yang, X.Y.; Qin, X.H.; Xiong, J.Y.; Wang, Y.Q.; Ren, D.T.; Liu, G.Q. A cotton kinesin GhKCH2 interacts with both microtubules and microfilaments. Biochem. J. 2009, 421, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Zhang, Y.; Kang, E.F.; Xu, Q.Y.; Wang, M.Y.; Rui, Y.; Liu, B.Q.; Yuan, M.; Fu, Y. MAP18 regulates the direction of pollen tube growth in Arabidopsis by modulating F-actin organization. Plant Cell 2013, 25, 851–867. [Google Scholar] [CrossRef] [Green Version]
- Qin, T.; Liu, X.M.; Li, J.J.; Sun, J.B.; Song, L.N.; Mao, T.L. Arabidopsis microtubule-destabilizing protein 25 functions in pollen tube growth by severing actin filaments. Plant Cell 2014, 26, 325–339. [Google Scholar] [CrossRef] [Green Version]
- Deeks, M.J.; Fendrych, M.; Smertenko, A.; Bell, K.S.; Oparka, K.; Cvrckova, F.; Zarsky, V.; Hussey, P.J. The plant formin AtFH4 interacts with both actin and microtubules, and contains a newly identified microtubule-binding domain. J. Cell Sci. 2010, 123, 1209–1215. [Google Scholar] [CrossRef] [Green Version]
- Rosero, A.; Zarsky, V.; Cvrckova, F. AtFH1 formin mutation affects actin filament and microtubule dynamics in Arabidopsis thaliana. J. Exp. Bot. 2013, 64, 585–597. [Google Scholar] [CrossRef]
- Wang, J.J.; Zhang, Y.; Wu, J.; Meng, L.; Ren, H.Y. AtFH16, an Arabidopsis type II formin, binds and bundles both microfilaments and microtubules, and preferentially binds to microtubules. J. Integr. Plant Biol. 2013, 55, 1002–1015. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Persson, S.; Zhang, Y. The connection of cytoskeletal network with plasma membrane and the cell wall. J. Integr. Plant Biol. 2015, 57, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Tolmie, F.; Poulet, A.; McKenna, J.; Sassmann, S.; Graumann, K.; Deeks, M.; Runions, J. The cell wall of Arabidopsis thaliana influences actin network dynamics. J. Exp. Bot. 2017, 68, 4517–4527. [Google Scholar] [CrossRef]
- Thamarus, K.; Groom, K.; Bradley, A.; Raymond, C.A.; Schimleck, L.R.; Williams, E.R.; Moran, G.F. Identification of quantitative trait loci for wood and fibre properties in two full-sib pedigrees of Eucalyptus globulus. Theor. Appl. Genet. 2004, 109, 856–864. [Google Scholar] [CrossRef]
- Freeman, J.S.; Whittock, S.P.; Potts, B.M.; Vaillancourt, R.E. QTL influencing growth and wood properties in Eucalyptus globulus. Tree Genet. Genomes 2009, 5, 713–722. [Google Scholar] [CrossRef] [Green Version]
- Thumma, B.R.; Southerton, S.G.; Bell, J.C.; Owen, J.V.; Henery, M.L.; Moran, G.F. Quantitative trait locus (QTL) analysis of wood quality traits in Eucalyptus nitens. Tree Genet. Genomes 2010, 6, 305–317. [Google Scholar] [CrossRef]
- Dillon, S.K.; Nolan, M.; Li, W.; Bell, C.; Wu, H.X.; Southerton, S.G. Allelic variation in cell wall candidate genes affecting solid wood properties in natural populations and land races of Pinus radiata. Genetics 2010, 185, 1477–1487. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, J.; Doerksen, T.; Boyle, B.; Clement, S.; Deslauriers, M.; Beauseigle, S.; Blais, S.; Poulin, P.-L.; Lenz, P.; Caron, S.; et al. Association genetics of wood physical traits in the conifer white spruce and relationships with gene expression. Genetics 2011, 188, 197–214. [Google Scholar] [CrossRef] [Green Version]
- Li, X.G.; Wu, H.X.; Southerton, S.G. Identification of putative candidate genes for juvenile wood density in Pinus radiata. Tree Physiol. 2012, 32, 1046–1057. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Martinez, S.C.; Wheeler, N.C.; Ersoz, E.; Nelson, C.D.; Neale, D.B. Association genetics in Pinus taeda L. I. Wood property traits. Genetics 2007, 175, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Pilate, G.; Dejardin, A.; Laurans, F.; Leple, J.C. Tension wood as a model for functional genomics of wood formation. New Phytol. 2004, 164, 63–72. [Google Scholar] [CrossRef]
- Paux, E.; Carocha, V.; Marques, C.; de Sousa, A.M.; Borralho, N.; Sivadon, P.; Grima-Pettenati, J. Transcript profiling of Eucalyptus xylem genes during tension wood formation. New Phytol. 2005, 167, 89–100. [Google Scholar] [CrossRef]
- Li, X.G.; Yang, X.H.; Wu, H.X. Transcriptome profiling of Radiata pine branches reveals new insights into reaction wood formation with implications in plant gravitropism. BMC Genom. 2013, 14, 768. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.H.; Chen, B.B.; Zhang, D.Q. Transcript profiling of Populus tomentosa genes in normal, tension, and opposite wood by RNA-seq. BMC Genom. 2015, 16, 164. [Google Scholar] [CrossRef] [Green Version]
- Lafarguette, F.; Leple, J.C.; Dejardin, A.; Laurans, F.; Costa, G.; Lesage-Descauses, M.C.; Pilate, G. Poplar genes encoding fasciclin-like arabinogalactan proteins are highly expressed in tension wood. New Phytol. 2004, 164, 107–121. [Google Scholar] [CrossRef]
- Andersson-Gunneras, S.; Mellerowicz, E.J.; Love, J.; Segerman, B.; Ohmiya, Y.; Coutinho, P.M.; Nilsson, P.; Henrissat, B.; Moritz, T.; Sundberg, B. Biosynthesis of cellulose-enriched tension wood in Populus: Global analysis of transcripts and metabolites identifies biochemical and developmental regulators in secondary wall biosynthesis. Plant J. 2006, 45, 144–165. [Google Scholar] [CrossRef]
- Qiu, D.; Wilson, I.W.; Gan, S.; Washusen, R.; Moran, G.F.; Southerton, S.G. Gene expression in Eucalyptus branch wood with marked variation in cellulose microfibril orientation and lacking g-layers. New Phytol. 2008, 179, 94–103. [Google Scholar] [CrossRef]
- MacMillan, C.P.; Mansfield, S.D.; Stachurski, Z.H.; Evans, R.; Southerton, S.G. Fasciclin-like arabinogalactan proteins: Specialization for stem biomechanics and cell wall architecture in Arabidopsis and Eucalyptus. Plant J. 2010, 62, 689–703. [Google Scholar] [CrossRef]
- Azri, W.; Ennajah, A.; Nasr, Z.; Woo, S.Y.; Khaldi, A. Transcriptome profiling the basal region of poplar stems during the early gravitropic response. Biol. Plant. 2014, 58, 55–63. [Google Scholar] [CrossRef]
- Bygdell, J.; Srivastava, V.; Obudulu, O.; Srivastava, M.K.; Nilsson, R.; Sundberg, B.; Trygg, J.; Mellerowicz, E.J.; Wingsle, G. Protein expression in tension wood formation monitored at high tissue resolution in Populus. J. Exp. Bot. 2017, 68, 3405–3417. [Google Scholar] [CrossRef] [Green Version]
- Pomies, L.; Decourteix, M.; Franchel, J.; Moulia, B.; Leblanc-Fournier, N. Poplar stem transcriptome is massively remodelled in response to single or repeated mechanical stimuli. BMC Genom. 2017, 18, 300. [Google Scholar] [CrossRef]
- Zinkgraf, M.; Gerttula, S.; Zhao, S.; Filkov, V.; Groover, A. Transcriptional and temporal response of Populus stems to gravi-stimulation. J. Integr. Plant Biol. 2018, 60, 578–590. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, S.; Fujino, T.; Thammanagowda, S.; Zhang, D.Y.; Xu, F.Y.; Joshi, C.P. Xylem-specific and tension stress-responsive coexpression of korrigan endoglucanase and three secondary wall-associated cellulose synthase genes in aspen trees. Planta 2006, 224, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Li, X.G.; Wu, H.X.; Southerton, S.G. Transcriptome profiling of Pinus radiata juvenile wood with contrasting stiffness identifies putative candidate genes involved in microfibril orientation and cell wall mechanics. BMC Genom. 2011, 12, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wu, H.X.; Southerton, S.G. Transcriptome profiling of wood maturation in Pinus radiata identifies differentially expressed genes with implications in juvenile and mature wood variation. Gene 2011, 487, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sederoff, R.R.; Allona, I. Differential expression of genes encoding cell wall proteins in vascular tissues from vertical and bent Loblolly pine trees. Tree Physiol. 2000, 20, 457–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whetten, R.; Sun, Y.H.; Zhang, Y.; Sederoff, R. Functional genomics and cell wall biosynthesis in Loblolly pine. Plant Mol. Biol. 2001, 47, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Yoshida, M.; Yamamoto, H.; Okuyama, T. Screening genes that change expression during compression wood formation in Chamaecyparis obtusa. Tree Physiol. 2008, 28, 1331–1340. [Google Scholar] [CrossRef] [Green Version]
- Le Provost, G.; Paiva, J.; Pot, D.; Brach, J.; Plomion, C. Seasonal variation in transcript accumulation in wood-forming tissues of maritime pine (Pinus pinaster Ait.) with emphasis on a cell wall glycine-rich protein. Planta 2003, 217, 820–830. [Google Scholar] [CrossRef]
- Showalter, A.M. Arabinogalactan-proteins: Structure, expression and function. Cell. Mol. Life Sci. 2001, 58, 1399–1417. [Google Scholar] [CrossRef]
- Driouich, A.; Baskin, T.I. Intercourse between cell wall and cytoplasma exemplified by arabinogalactan proteins and cortical microtubules. Am. J. Bot. 2008, 95, 1491–1497. [Google Scholar] [CrossRef] [Green Version]
- Putoczki, T.L.; Pettolino, F.; Griffin, M.D.W.; Moller, R.; Gerrard, J.A.; Bacic, A.; Jackson, S.L. Characterization of the structure, expression and function of Pinus radiata D. Don arabinogalactan-proteins. Planta 2007, 226, 1131–1142. [Google Scholar] [CrossRef]
- Andeme-Onzighi, C.; Sivaguru, M.; Judy-March, J.; Baskin, T.I.; Driouich, A. The reb1-1 mutation of Arabidopsis alters the morphology of trichoblasts, the expression of arabinogalactan-proteins and the organization of cortical microtubules. Planta 2002, 215, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Sardar, H.S.; Yang, J.; Showalter, A.M. Molecular interactions of arabinogalactan proteins with cortical microtubules and F-actin in bright yellow-2 tobacco cultured cells. Plant Physiol. 2006, 142, 1469–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguema-Ona, E.; Bannigan, A.; Chevalier, L.; Baskin, T.I.; Driouich, A. Disruption of arabinogalactan proteins disorganizes cortical microtubules in the root of Arabidopsis thaliana. Plant J. 2007, 52, 240–251. [Google Scholar] [CrossRef] [PubMed]
- DeBolt, S.; Gutierrez, R.; Ehrhardt, D.W.; Melo, C.V.; Ross, L.; Cutler, S.R.; Somerville, C.; Bonetta, D. Morlin, an inhibitor of cortical microtubule dynamics and cellulose synthase movement. Proc. Natl. Acad. Sci. USA 2007, 104, 5854–5859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Stegmann, M.; Han, Z.F.; DeFalco, T.A.; Parys, K.; Xu, L.; Belkhadir, Y.; Zipfel, C.; Chai, J.J. Mechanisms of RALF peptide perception by a heterotypic receptor complex. Nature 2019, 572, 270–274. [Google Scholar] [CrossRef]
- Moody, R.G.; Williamson, M.P. Structure and function of a bacterial Fasciclin I domain protein elucidates function of related cell adhesion proteins such as TGFBIp and periostin. FEBS Open Bio 2013, 3, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Gaspar, Y.; Johnson, K.L.; McKenna, J.A.; Bacic, A.; Schultz, C.J. The complex structures of arabinogalactan-proteins and the journey towards understanding function. Plant Mol. Biol. 2001, 47, 161–176. [Google Scholar] [CrossRef]
- Seifert, G.J.; Roberts, K. The biology of arabinogalactan proteins. Annu. Rev. Plant Biol. 2007, 58, 137–161. [Google Scholar] [CrossRef]
- Johnson, K.L.; Jones, B.J.; Bacic, A.; Schultz, C.J. The fasciclin-like arabinogalactan proteins of Arabidopsis. A multigene family of putative cell adhesion molecules. Plant Physiol. 2003, 133, 1911–1925. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.L.; Zhao, J. Genome-wide identification, classification, and expression analysis of the arabinogalactan protein gene family in rice (Oryza sativa L.). J. Exp. Bot. 2010, 61, 2647–2668. [Google Scholar] [CrossRef]
- Huang, G.Q.; Xu, W.L.; Gong, S.Y.; Li, B.; Wang, X.L.; Xu, D.; Li, X.B. Characterization of 19 novel cotton FLA genes and their expression profiling in fiber development and in response to phytohormones and salt stress. Physiol. Plant. 2008, 134, 348–359. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, C.P.; Taylor, L.; Bi, Y.D.; Southerton, S.G.; Evans, R.; Spokevicius, A. The fasciclin-like arabinogalactan protein family of Eucalyptus grandis contains members that impact wood biology and biomechanics. New Phytol. 2015, 206, 1314–1327. [Google Scholar] [CrossRef] [PubMed]
- Persson, S.; Wei, H.R.; Milne, J.; Page, G.P.; Somerville, C.R. Identification of genes required for cellulose synthesis by regression analysis of public microarray data sets. Proc. Natl. Acad. Sci. USA 2005, 102, 8633–8638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahiya, P.; Findlay, K.; Roberts, K.; McCann, M.C. A fasciclin-domain containing gene, ZeFLA11, is expressed exclusively in xylem elements that have reticulate wall thickenings in the stem vascular system of Zinnia elegans cv Envy. Planta 2006, 223, 1281–1291. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.L.; Rahman, A.; Baskin, T.I.; Kieber, J.J. Two leucine-rich repeat receptor kinases mediate signaling, linking cell wall biosynthesis and ACC synthase in Arabidopsis. Plant Cell 2008, 20, 3065–3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Zhang, R.; Gui, J.; Zhong, Y.; Li, L. The receptor-like kinase AtVRLK1 regulates secondary cell wall thickening. Plant Physiol. 2018, 177, 671–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.H.; Jiang, C.M.; Wang, C.T.; Yang, Y.; Yang, L.; Gao, X.Y.; Zhang, H.X. Antisense expression of the fasciclin-like arabinogalactan protein FLA6 gene in Populus inhibits expression of its homologous genes and alters stem biomechanics and cell wall composition in transgenic trees. J. Exp. Bot. 2015, 66, 1291–1302. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.H.; Jin, Y.L.; Wang, C.T.; Li, B.; Jiang, C.M.; Sun, Z.C.; Zhang, Z.P.; Kong, F.J.; Zhang, H.X. Fasciclin-like arabinogalactan proteins, PtFLAs, play important roles in ga-mediated tension wood formation in Populus. Sci. Rep. 2017, 7, 6182. [Google Scholar] [CrossRef]
- Oakley, R.V.; Wang, Y.S.; Ramakrishna, W.; Harding, S.A.; Tsai, C.J. Differential expansion and expression of alpha- and beta-tubulin gene families in Populus. Plant Physiol. 2007, 145, 961–973. [Google Scholar] [CrossRef] [Green Version]
- Breviario, D.; Giani, S.; Morello, L. Multiple tubulins: Evolutionary aspects and biological implications. Plant J. 2013, 75, 202–218. [Google Scholar] [CrossRef]
- Cheng, Z.G.; Snustad, D.P.; Carter, J.V. Temporal and spatial expression patterns of TUB9, a beta-tubulin gene of Arabidopsis thaliana. Plant Mol. Biol. 2001, 47, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Snustad, D.P.; Haas, N.A.; Kopczak, S.D.; Silflow, C.D. The small genome of Arabidopsis contains at least 9 expressed beta-tubulin genes. Plant Cell 1992, 4, 549–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, M.; Yang, G.X.; Kawaguchi, K.; Komatsu, S. Expression analyses of beta-tubulin isotype genes in rice. Plant Cell Physiol. 2003, 44, 1202–1207. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, D.J.; Triplett, B.A. Gene-specific changes in alpha-tubulin transcript accumulation in developing cotton fibers. Plant Physiol. 1999, 121, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, P.G.; Borisy, G.G. Evolution of the multi-tubulin hypothesis. Bioessays 1997, 19, 451–454. [Google Scholar] [CrossRef] [PubMed]
- Parrotta, L.; Cresti, M.; Cai, G. Accumulation and post-translational modifications of plant tubulins. Plant Biol. 2014, 16, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Swamy, P.S.; Hu, H.; Pattathil, S.; Maloney, V.J.; Xiao, H.; Xue, L.J.; Chung, J.D.; Johnson, V.E.; Zhu, Y.Y.; Peter, G.F.; et al. Tubulin perturbation leads to unexpected cell wall modifications and affects stomatal behaviour in Populus. J. Exp. Bot. 2015, 66, 6507–6518. [Google Scholar] [CrossRef] [Green Version]
- Nicol, F.; His, I.; Jauneau, A.; Vernhettes, S.; Canut, H.; Hofte, H. A plasma membrane-bound putative endo-1,4-beta-d-glucanase is required for normal wall assembly and cell elongation in Arabidopsis. EMBO J. 1998, 17, 5563–5576. [Google Scholar] [CrossRef] [Green Version]
- Maloney, V.J.; Mansfield, S.D. Characterization and varied expression of a membrane-bound endo-beta-1,4-glucanase in hybrid poplar. Plant Biotechnol. J. 2010, 8, 294–307. [Google Scholar] [CrossRef]
- His, I.; Driouich, A.; Nicol, F.; Jauneau, A.; Hofte, H. Altered pectin composition in primary cell walls of korrigan, a dwarf mutant of Arabidopsis deficient in a membrane-bound endo-1,4-beta-glucanase. Planta 2001, 212, 348–358. [Google Scholar] [CrossRef]
- Szyjanowicz, P.M.J.; McKinnon, I.; Taylor, N.G.; Gardiner, J.; Jarvis, M.C.; Turner, S.R. The irregular xylem 2 mutant is an allele of korrigan that affects the secondary cell wall of Arabidopsis thaliana. Plant J. 2004, 37, 730–740. [Google Scholar] [CrossRef] [PubMed]
- Hertzberg, M.; Aspeborg, H.; Schrader, J.; Andersson, A.; Erlandsson, R.; Blomqvist, K.; Bhalerao, R.; Uhlen, M.; Teeri, T.T.; Lundeberg, J.; et al. A transcriptional roadmap to wood formation. Proc. Natl. Acad. Sci. USA 2001, 98, 14732–14737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, H.; Kawahara, A.; Inoue, Y. Ethylene promotes the induction by auxin of the cortical microtubule randomization required for low-ph-induced root hair initiation in lettuce (Lactuca sativa L.) seedlings. Plant Cell Physiol. 2003, 44, 932–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Derba-Maceluch, M.; Awano, T.; Takahashi, J.; Lucenius, J.; Ratke, C.; Kontro, I.; Busse-Wicher, M.; Kosik, O.; Tanaka, R.; Winzell, A.; et al. Suppression of xylan endotransglycosylase PtxtXyn10A affects cellulose microfibril angle in secondary wall in aspen wood. New Phytol. 2015, 205, 666–681. [Google Scholar] [CrossRef] [Green Version]
- Mareri, L.; Romi, M.; Cai, G. Arabinogalactan proteins: Actors or spectators during abiotic and biotic stress in plants? Plant Biosyst. 2019, 153, 173–185. [Google Scholar] [CrossRef]
- Landrein, B.; Hamant, O. How mechanical stress controls microtubule behavior and morphogenesis in plants: History, experiments and revisited theories. Plant J. 2013, 75, 324–338. [Google Scholar] [CrossRef]
- Monshausen, G.B.; Haswell, E.S. A force of nature: Molecular mechanisms of mechanoperception in plants. J. Exp. Bot. 2013, 64, 4663–4680. [Google Scholar] [CrossRef] [Green Version]
- Showalter, A.M.; Basu, D. Extensin and arabinogalactan-protein biosynthesis: Glycosyltransferases, research challenges, and biosensors. Front. Plant Sci. 2016, 7, 814. [Google Scholar] [CrossRef] [Green Version]
- Lamport, D.T.A.; Varnai, P. Periplasmic arabinogalactan glycoproteins act as a calcium capacitor that regulates plant growth and development. New Phytol. 2013, 197, 58–64. [Google Scholar] [CrossRef]
- Li, M.; Hong, Y.; Wang, X. Phospholipase d- and phosphatidic acid-mediated signaling in plants. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2009, 1791, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Smertenko, A.P.; Chang, H.Y.; Sonobe, S.; Fenyk, S.I.; Weingartner, M.; Bogre, L.; Hussey, P.J. Control of the AtMAP65-1 interaction with microtubules through the cell cycle. J. Cell Sci. 2006, 119, 3227–3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Lin, F.; Mao, T.L.; Nie, J.N.; Yan, M.; Yuan, M.; Zhang, W.H. Phosphatidic acid regulates microtubule organization by interacting with MAP65-1 in response to salt stress in Arabidopsis. Plant Cell 2012, 24, 4555–4576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoppin-Mellet, V.; Fache, V.; Portran, D.; Martiel, J.L.; Vantard, M. MAP65 coordinate microtubule growth during bundle formation. PLoS ONE 2013, 8, e56808. [Google Scholar] [CrossRef]
- Wang, C.; Li, J.J.; Yuan, M. Salt tolerance requires cortical microtubule reorganization in Arabidopsis. Plant Cell Physiol. 2007, 48, 1534–1547. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.X.; Nick, P. Cold sensing in grapevine-which signals are upstream of the microtubular “thermometer”. Plant Cell Environ. 2017, 40, 2844–2857. [Google Scholar] [CrossRef]
- Endler, A.; Kesten, C.; Schneider, R.; Zhang, Y.; Ivakov, A.; Froehlich, A.; Funke, N.; Persson, S. A mechanism for sustained cellulose synthesis during salt stress. Cell 2015, 162, 1353–1364. [Google Scholar] [CrossRef] [Green Version]
- Lillo, C.; Kataya, A.R.A.; Heidari, B.; Creighton, M.T.; Nemie-Feyissa, D.; Ginbot, Z.; Jonassen, E.M. Protein phosphatases PP2A, PP4 and PP6: Mediators and regulators in development and responses to environmental cues. Plant Cell Environ. 2014, 37, 2631–2648. [Google Scholar] [CrossRef]
- Heisler, M.G.; Hamant, O.; Krupinski, P.; Uyttewaal, M.; Ohno, C.; Jonsson, H.; Traas, J.; Meyerowitz, E.M. Alignment between PIN1 polarity and microtubule orientation in the shoot apical meristem reveals a tight coupling between morphogenesis and auxin transport. PLoS. Biol. 2010, 8, e1000516. [Google Scholar] [CrossRef] [Green Version]
- Hellgren, J.M.; Olofsson, K.; Sundberg, B. Patterns of auxin distribution during gravitational induction of reaction wood in poplar and pine. Plant Physiol. 2004, 135, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.D.; Wen, M.Z.; Nagawa, S.; Fu, Y.; Chen, J.G.; Wu, M.J.; Perrot-Rechenmann, C.; Friml, J.; Jones, A.M.; Yang, Z.B. Cell surface- and rho gtpase-based auxin signaling controls cellular interdigitation in Arabidopsis. Cell 2010, 143, 99–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.; Cao, L.; Zhou, Z.; Zhu, L.; Ehrhardt, D.; Yang, Z.; Fu, Y. Rho GTPase signaling activates microtubule severing to promote microtubule ordering in Arabidopsis. Curr. Biol. 2013, 23, 290–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uyttewaal, M.; Burian, A.; Alim, K.; Landrein, B.T.; Borowska-Wykret, D.; Dedieu, A.; Peaucelle, A.; Ludynia, M.; Traas, J.; Boudaoud, A.; et al. Mechanical stress acts via katanin to amplify differences in growth rate between adjacent cells in Arabidopsis. Cell 2012, 149, 439–451. [Google Scholar] [CrossRef] [Green Version]
- Li, C.J.; Lu, H.M.; Li, W.; Yuan, M.; Fu, Y. A ROP2-RIC1 pathway fine-tunes microtubule reorganization for salt tolerance in Arabidopsis. Plant Cell Environ. 2017, 40, 1127–1142. [Google Scholar] [CrossRef]
- Lindeboom, J.J.; Nakamura, M.; Hibbel, A.; Shundyak, K.; Gutierrez, R.; Ketelaar, T.; Emons, A.M.C.; Mulder, B.M.; Kirik, V.; Ehrhardt, D.W. A mechanism for reorientation of cortical microtubule arrays driven by microtubule severing. Science 2013, 342, 1245533. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tobias, L.M.; Spokevicius, A.V.; McFarlane, H.E.; Bossinger, G. The Cytoskeleton and Its Role in Determining Cellulose Microfibril Angle in Secondary Cell Walls of Woody Tree Species. Plants 2020, 9, 90. https://doi.org/10.3390/plants9010090
Tobias LM, Spokevicius AV, McFarlane HE, Bossinger G. The Cytoskeleton and Its Role in Determining Cellulose Microfibril Angle in Secondary Cell Walls of Woody Tree Species. Plants. 2020; 9(1):90. https://doi.org/10.3390/plants9010090
Chicago/Turabian StyleTobias, Larissa Machado, Antanas V. Spokevicius, Heather E. McFarlane, and Gerd Bossinger. 2020. "The Cytoskeleton and Its Role in Determining Cellulose Microfibril Angle in Secondary Cell Walls of Woody Tree Species" Plants 9, no. 1: 90. https://doi.org/10.3390/plants9010090