
Study of the Spatiotemporal Variability of Oceanographic Parameters and Their Relationship to Holothuria Species Abundance in a Marine Protected Area of the Mediterranean Using Satellite Imagery

Abstract
:1. Introduction
2. Materials and Methods
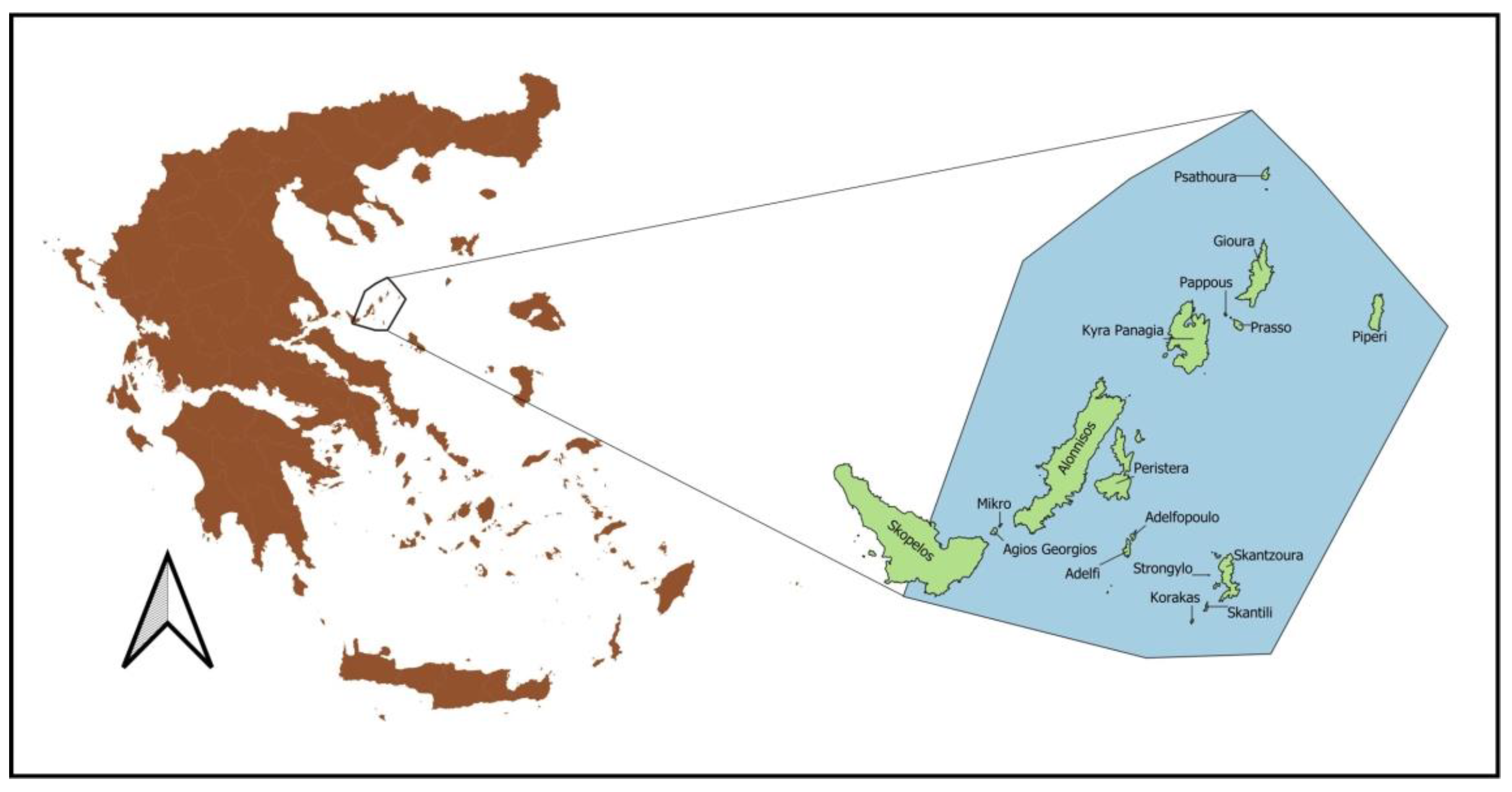
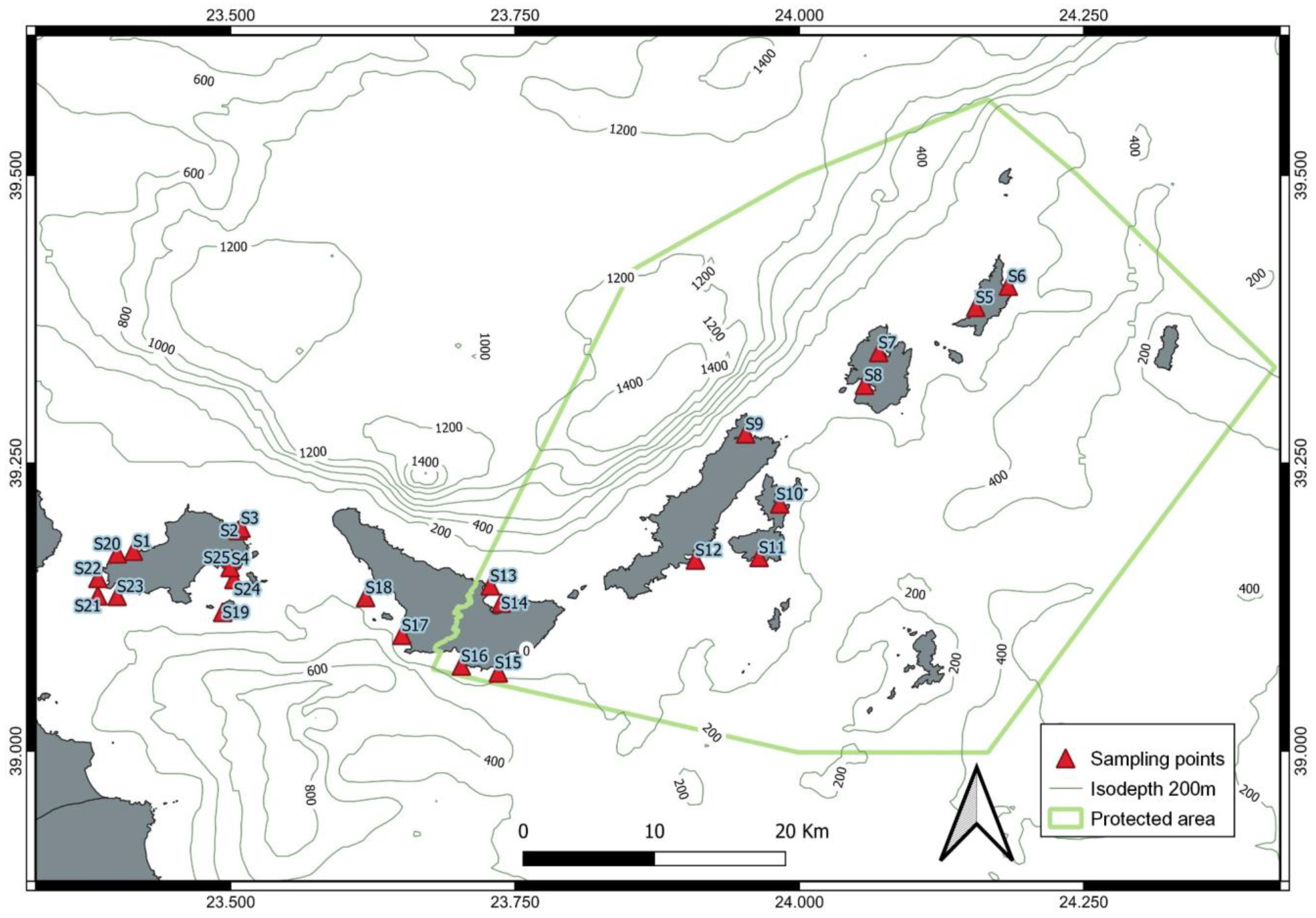
2.1. Study Area
2.2. In Situ Data
2.3. Satellite Database
3. Results
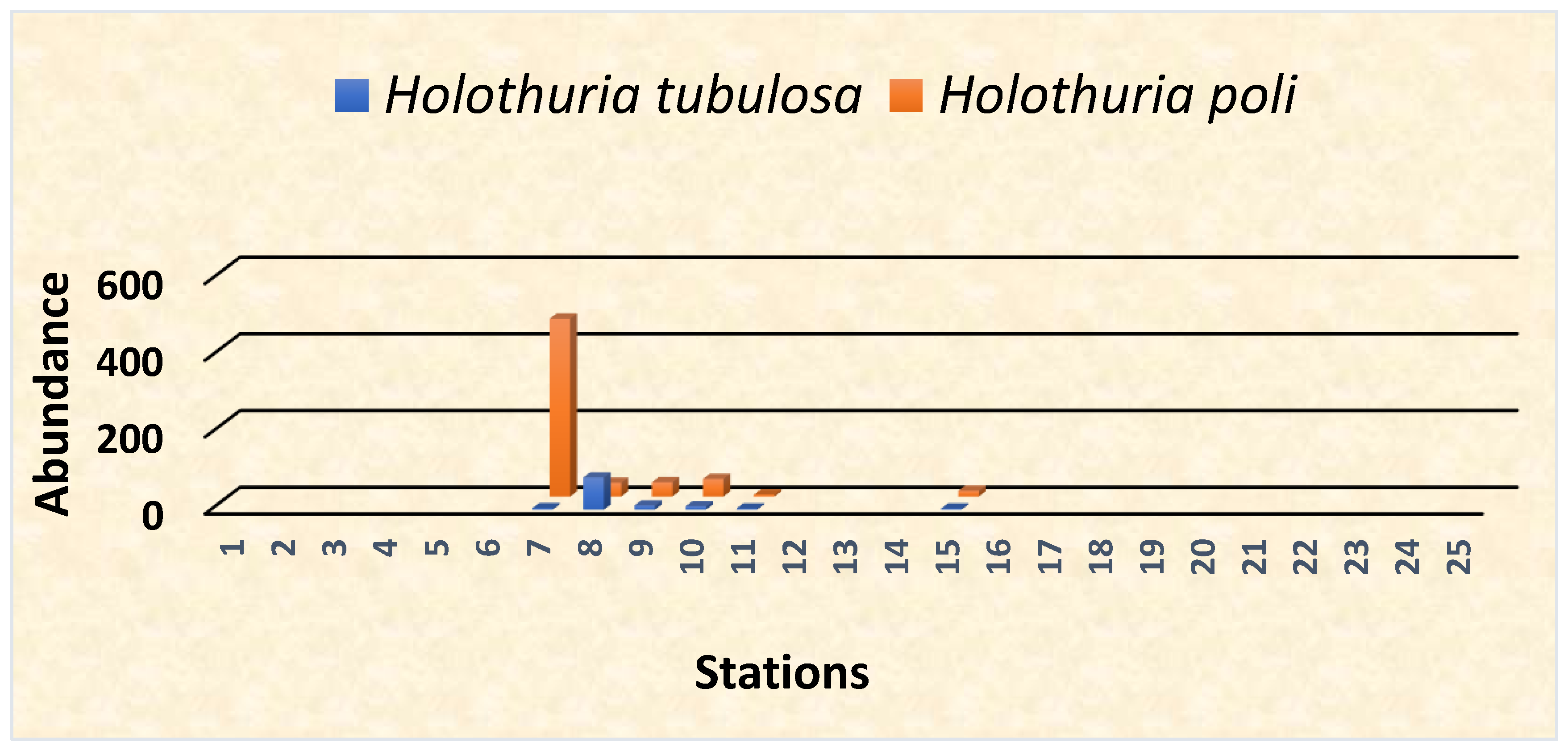
3.1. Species Abundance
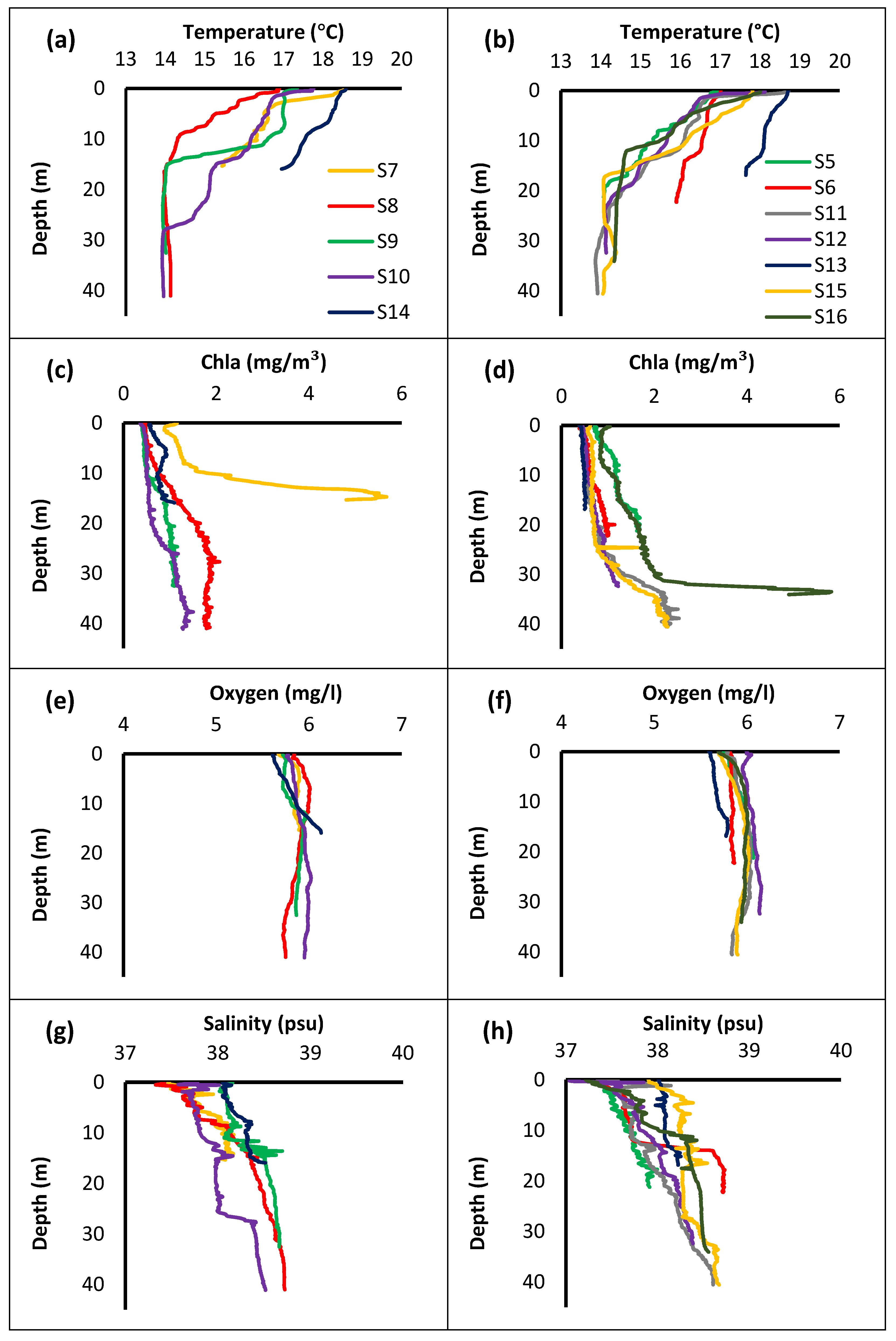
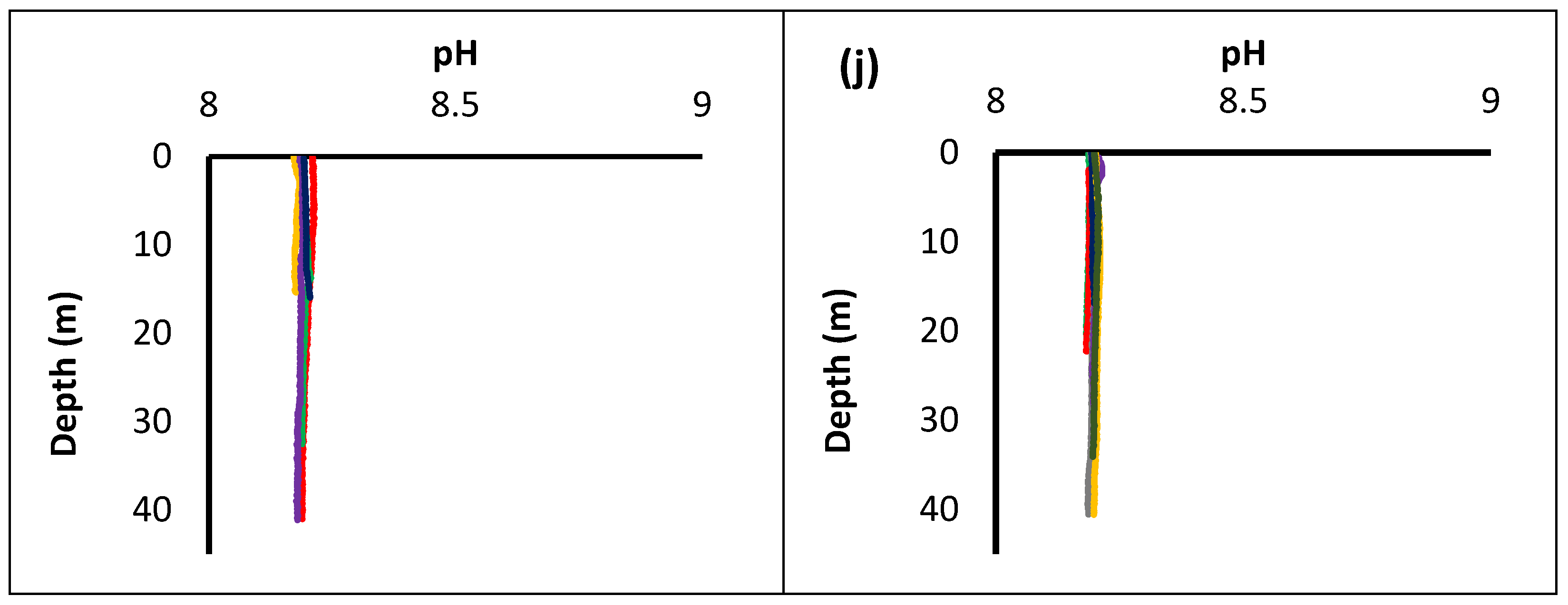
3.2. Physicochemical Parameters
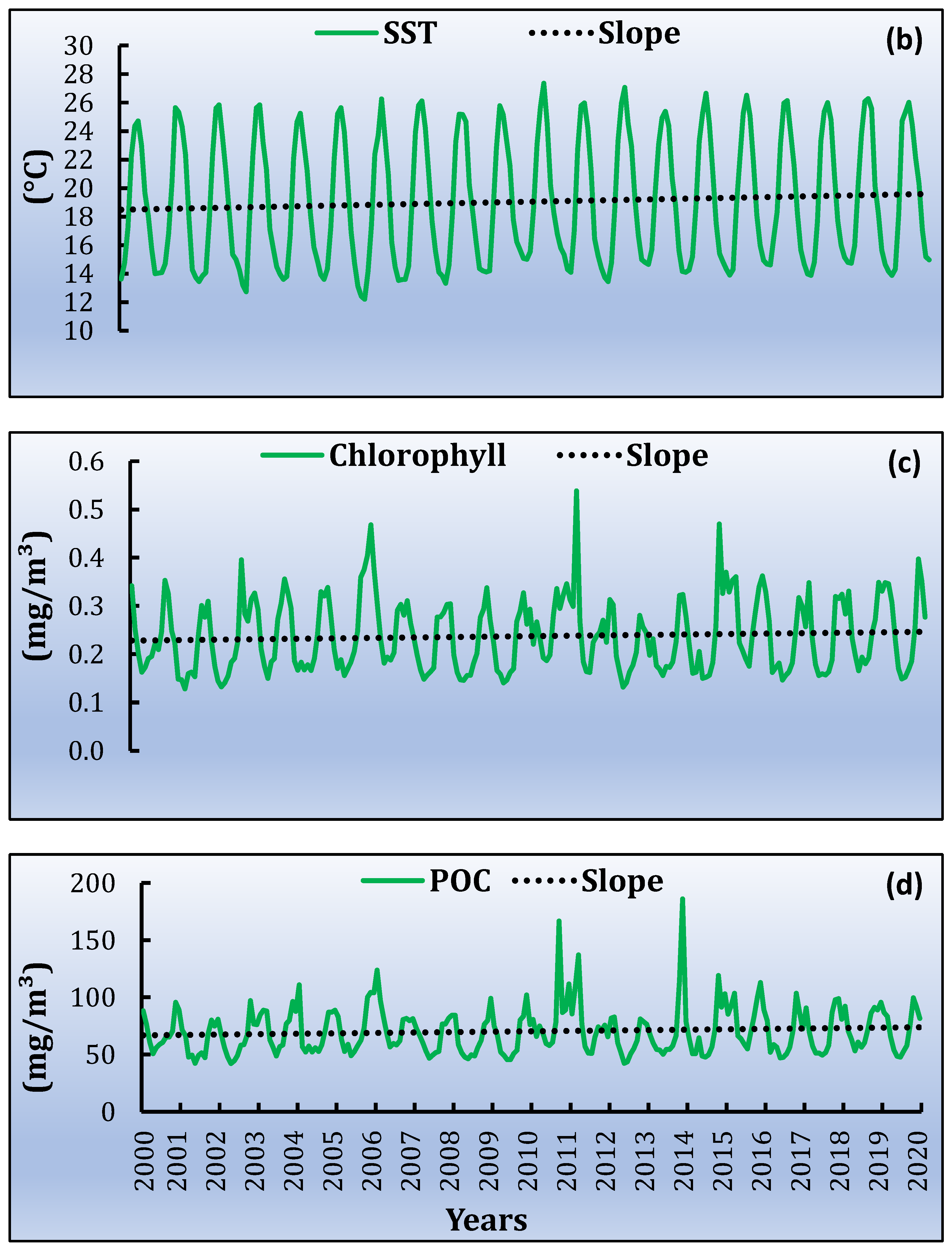
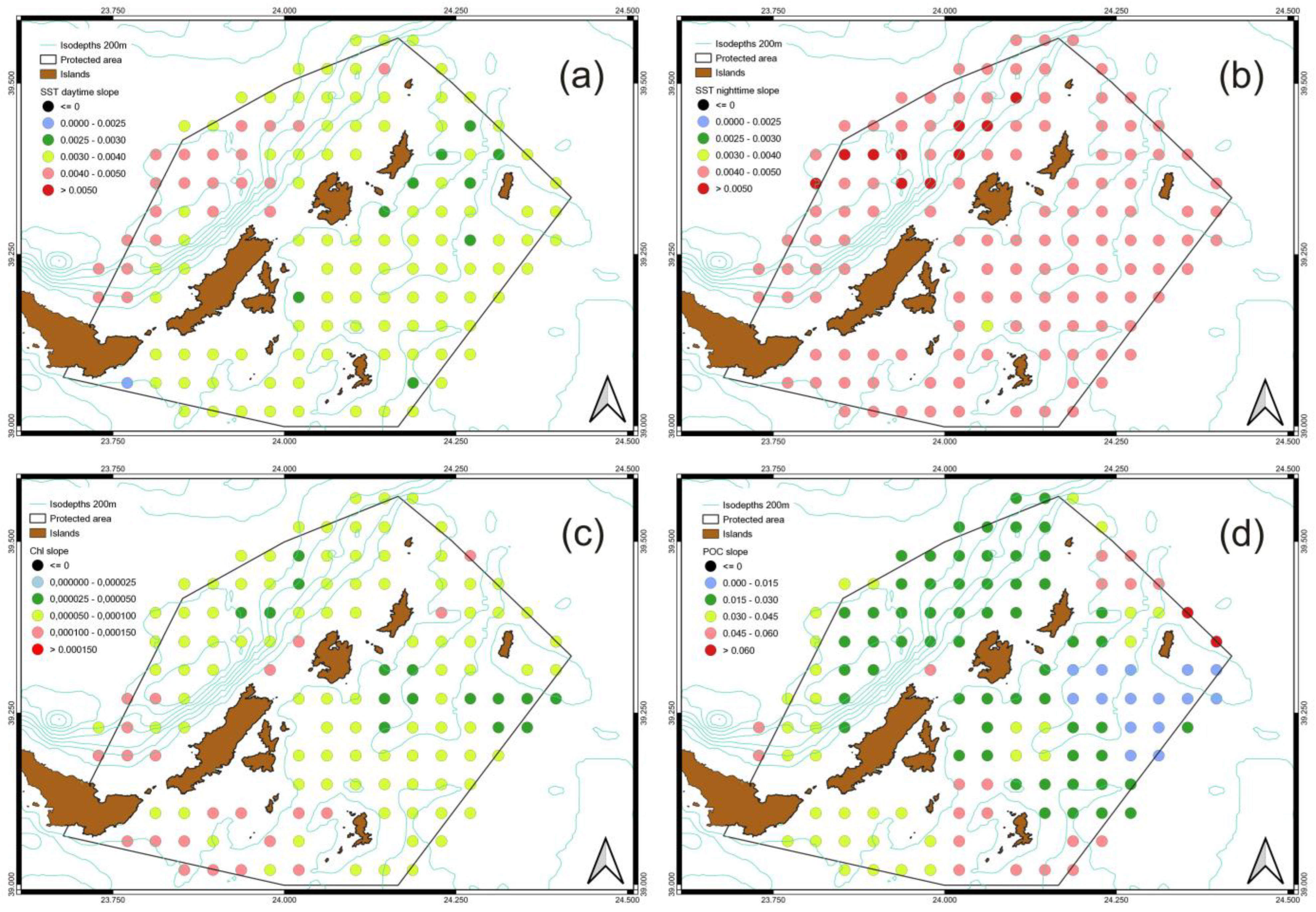
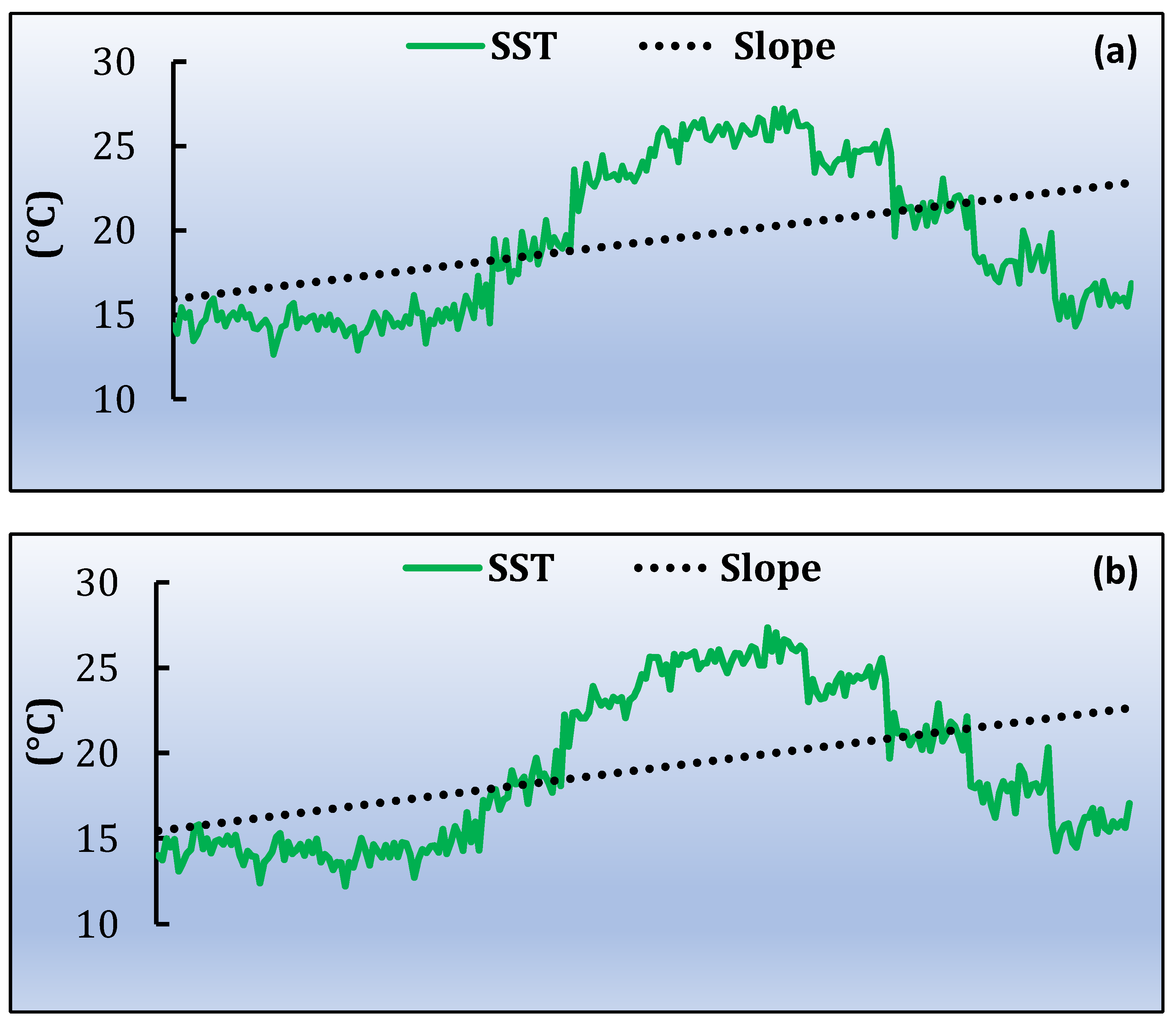
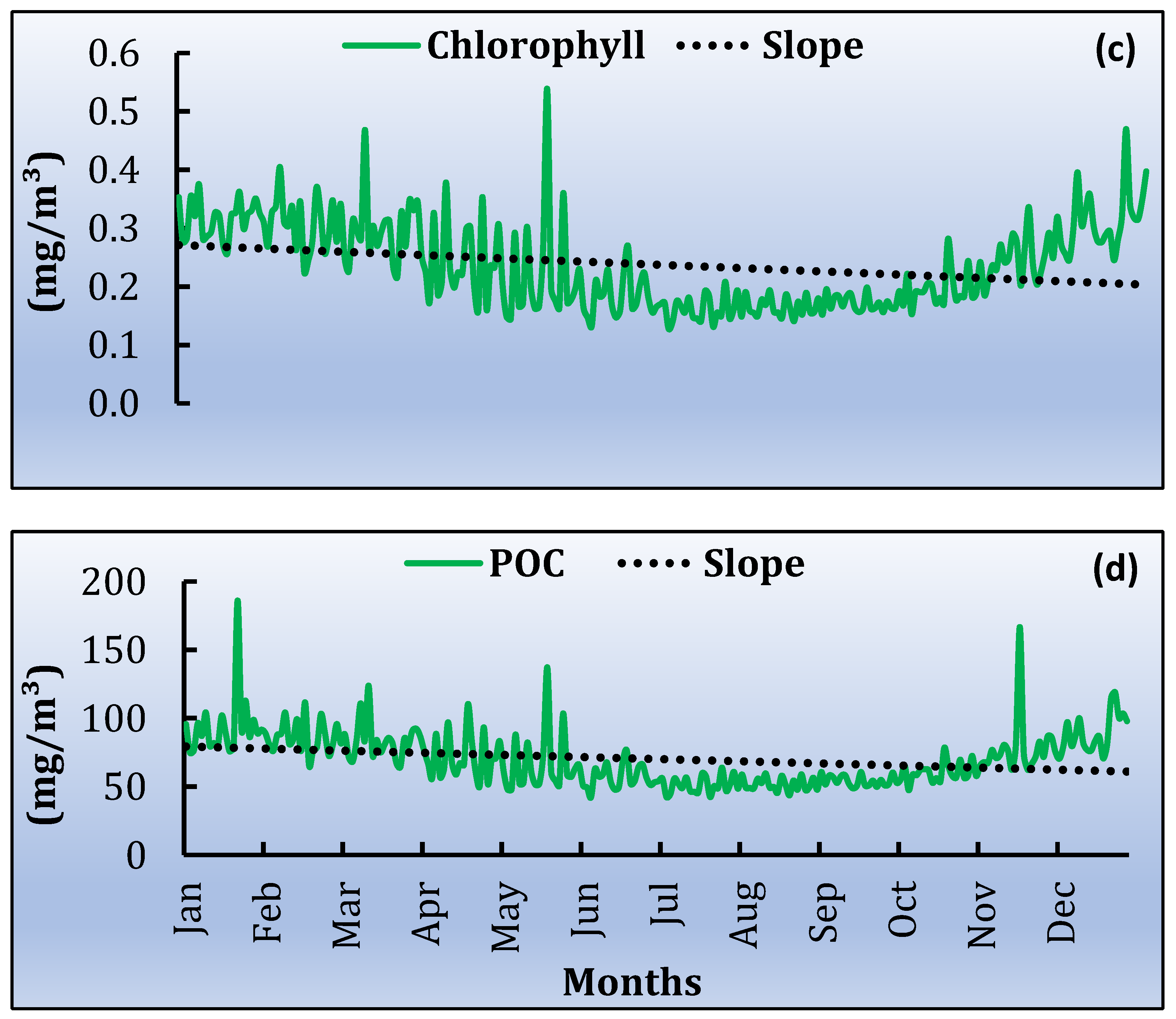
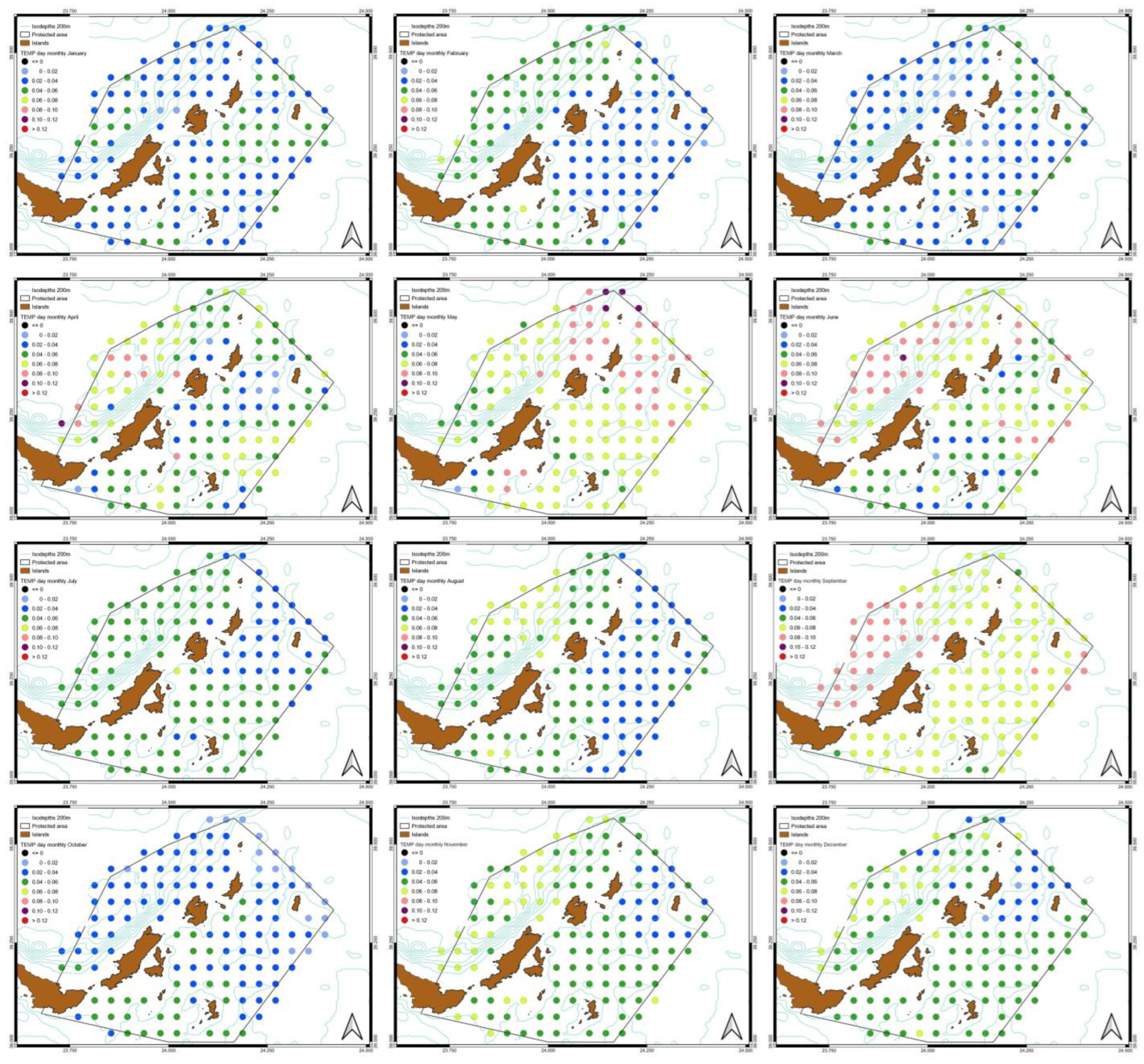
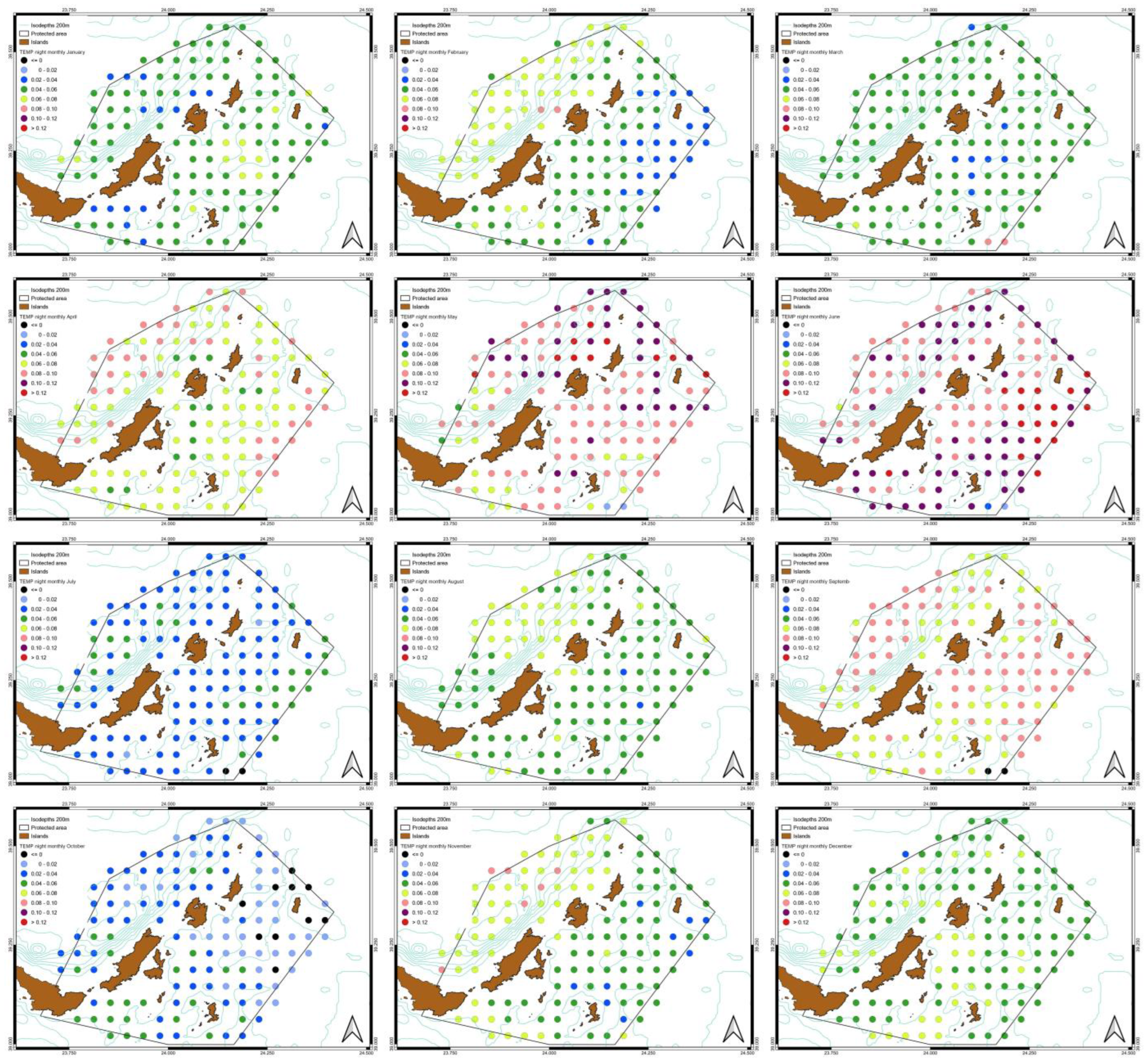
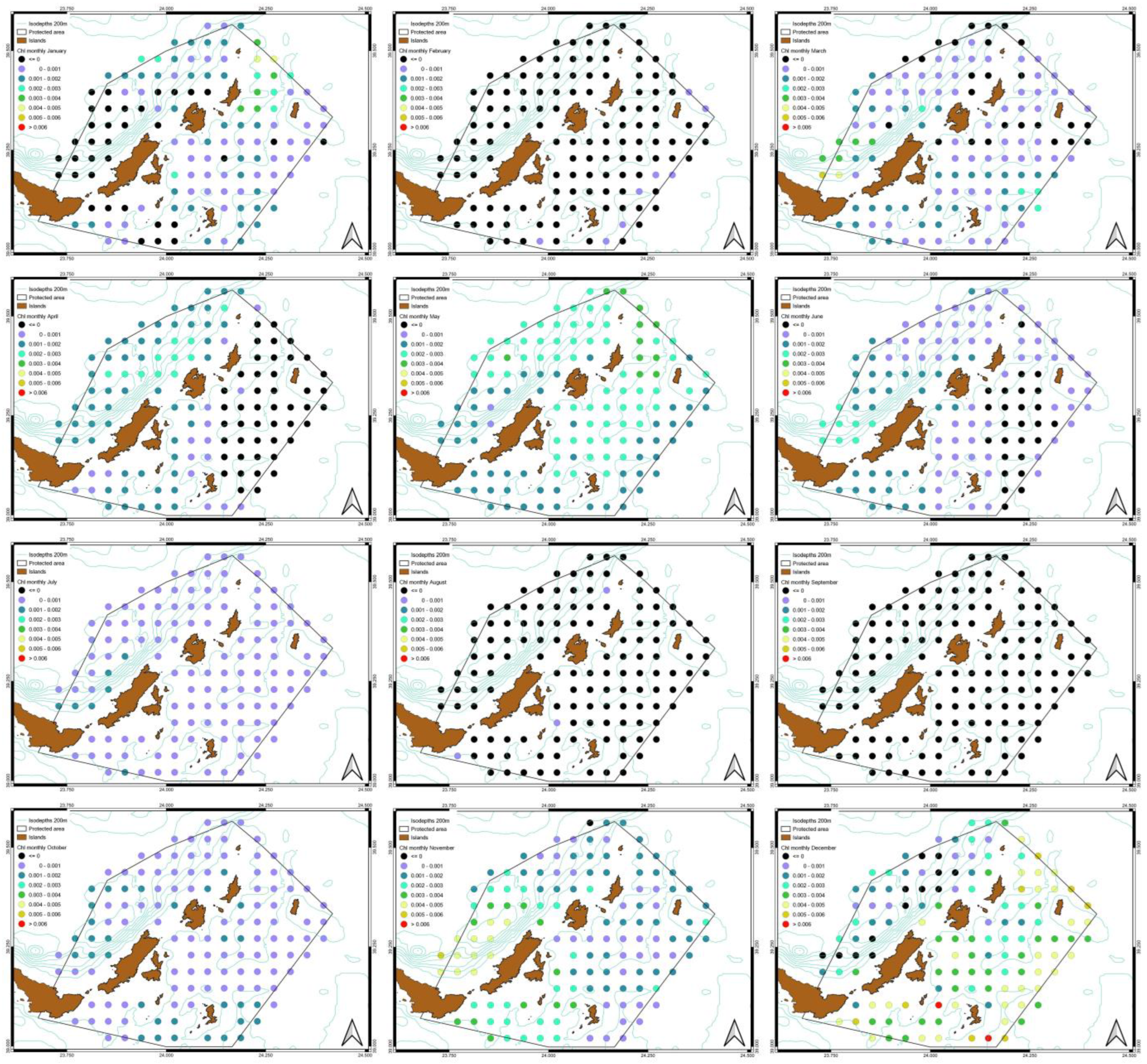
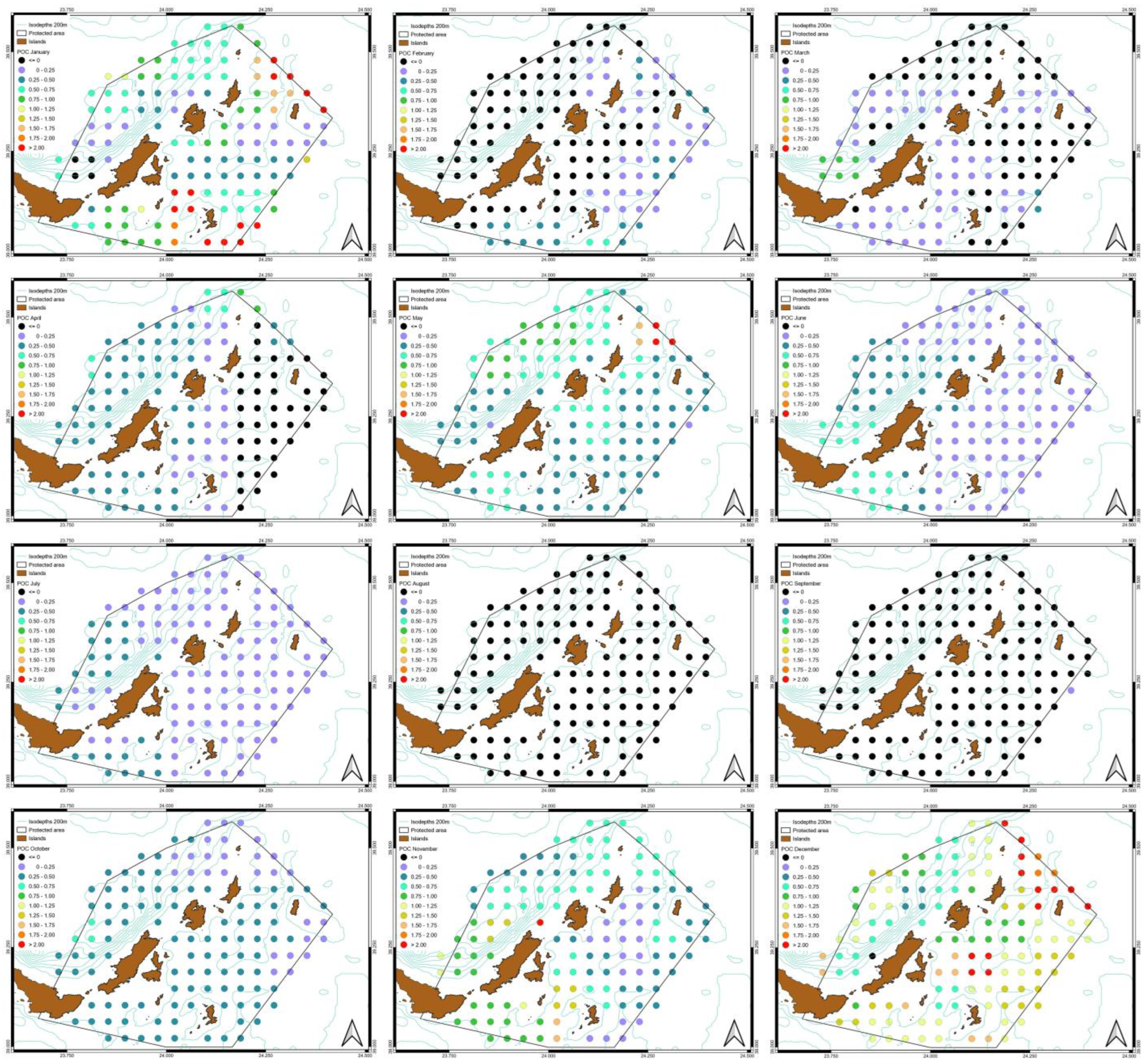
3.3. Spatiotemporal Variati Ons of SST, Chl-a and Particulate Organic Carbon [POC]
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Day, J.; Dudley, N.; Hockings, M.; Holmes, G.; Laffoley, D.; Stolton, S.; Wells, S.; Wenzel, L. Guidelines for Applying the IUCN Protected Area Management Categories to Marine Protected Areas, 2nd ed.; World Comission on Protected Areas: Gland, Switzerland, 2019; ISBN 9782831719412. [Google Scholar]
- Reuchlin-Hugenholtz, E.; McKenzie, E. Marine Protected Areas: Smart Investments in Ocean Health; Tanzer, J., Ed.; WWF: Gland, Switzerland, 2015; ISBN 9782940529216. [Google Scholar]
- Bertzk, B.; Corrigan, C.; Kemsey, J.; Kenney, S.; Ravilious, C.; Besançon, C.; Burgess, N. Protected Planet Report 2012: Tracking Progress Towards Global Targets for Protected Areas; IUCN: Gland, Switzerland, 2012; ISBN 9789280731897. [Google Scholar]
- Sala, E.; Giakoumi, S. No-Take Marine Reserves Are the Most Effective Protected Areas in the Ocean. ICES J. Mar. Sci. 2018, 75, 1166–1168. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Wang, Y.; Chen, Y.; Liu, K. Remote sensing approach for the estimation of particulate organic carbon in coastal waters based on suspended particulate concentration and particle median size. Mar. Pollut. Bull. 2020, 158, 111382. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Minnett, P.J. High latitude sea surface temperatures derived from MODIS infrared measurements. Remote Sens. Environ. 2020, 251, 112094. [Google Scholar] [CrossRef]
- Minnett, P.J.; Alvera-Azcárate, A.; Chin, T.M.; Corlett, G.K.; Gentemann, C.L.; Karagali, I.; Li, X.; Marsouin, A.; Marullo, S.; Maturi, E.; et al. Half a Century of Satellite Remote Sensing of Sea-Surface Temperature. Remote Sens. Environ. 2019, 233, 111366. [Google Scholar] [CrossRef]
- Kilpatrick, K.A.; Podestá, G.; Walsh, S.; Williams, E.; Halliwell, V.; Szczodrak, M.; Brown, O.B.; Minnett, P.J.; Evans, R. A decade of sea surface temperature from MODIS. Remote Sens. Environ. 2015, 165, 27–41. [Google Scholar] [CrossRef]
- Pahlevan, N.; Smith, B.; Binding, C.; Gurlin, D.; Li, L.; Bresciani, M.; Giardino, C. Hyperspectral retrievals of phytoplankton absorption and chlorophyll-a in inland and nearshore coastal waters. Remote Sens. Environ. 2021, 253, 112200. [Google Scholar] [CrossRef]
- Otsuka, A.Y.; Feitosa, F.A.N.; Flores-Montes, M.J.; Silva, A. Dynamics of Chlorophyll a and Oceanographic Parameters in the Coastal Zone: Barra das Jangadas-Pernambuco, Brazil. J. Coast. Res. 2016, 32, 490–499. [Google Scholar] [CrossRef]
- Fan, H.; Wang, X.; Zhang, H.; Yu, Z. Spatial and temporal variations of particulate organic carbon in the Yellow-Bohai Sea over 2002–2016. Sci. Rep. 2018, 8, 7971. [Google Scholar] [CrossRef] [Green Version]
- Guan, Q.; Feng, L.; Hou, X.; Schurgers, G.; Zheng, Y.; Tang, J. Eutrophication changes in fifty large lakes on the Yangtze Plain of China derived from MERIS and OLCI observations. Remote Sens. Environ. 2020, 246, 111890. [Google Scholar] [CrossRef]
- Zhang, L.; Liao, Q.; Gao, R.; Luo, R.; Liu, C.; Zhong, J.; Wang, Z. Spatial variations in diffusive methane fluxes and the role of eutrophication in a subtropical shallow lake. Sci. Total Environ. 2021, 759, 143495. [Google Scholar] [CrossRef]
- Henryson, K.; Kätterer, T.; Tidåker, P.; Sundberg, C. Soil N2O emissions, N leaching and marine eutrophication in life cycle assessment—A comparison of modelling approaches. Sci. Total Environ. 2020, 725, 138332. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.M.; Townsend-Small, A.; Zastepa, A.; Watson, S.B.; Brandes, J.A. Methane and nitrous oxide measured throughout Lake Erie over all seasons indicate highest emissions from the eutrophic Western Basin. J. Great Lakes Res. 2020, 46, 1604–1614. [Google Scholar] [CrossRef]
- Chen, Q.; Huang, M.; Tang, X. Eutrophication assessment of seasonal urban lakes in China Yangtze River Basin using Landsat 8-derived Forel-Ule index: A six-year (2013–2018) observation. Sci. Total Environ. 2020, 745, 135392. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, P.; Zhang, Z. Interaction between carbon dioxide emissions and eutrophication in a drinking water reservoir: A three-dimensional ecological modeling approach. Sci. Total Environ. 2019, 663, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, S.; Vethamony, P.; Sadooni, F.N.; Al-kuwari, H.A.; Al-khayat, J.A.; Seegobin, V.O.; Govil, H.; Nasir, S. Detection of Wakashio oil spill off Mauritius using Sentinel-1 and 2 data: Capability of sensors, image transformation methods and. Environ. Pollut. 2021, 274, 116618. [Google Scholar] [CrossRef]
- Balogun, A.L.; Yekeen, S.T.; Pradhan, B.; Wan Yusof, K.B. Oil spill trajectory modelling and environmental vulnerability mapping using GNOME model and GIS. Environ. Pollut. 2021, 268, 115812. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.R.; Grubesic, T.H. A spatiotemporal analysis of oil spill severity using a multi-criteria decision framework. Ocean Coast. Manag. 2021, 199, 105410. [Google Scholar] [CrossRef]
- Obida, C.B.; Blackburn, G.A.; Whyatt, J.D.; Semple, K.T. Counting the cost of the Niger Delta’s largest oil spills satellite remote sensing reveals extensive environmental damage with 1million people in the impact zone. Sci. Total Environ. 2021, 775, 145854. [Google Scholar] [CrossRef]
- Klemas, V. Remote sensing techniques for studying coastal ecosystems: An overview. J. Coast. Res. 2011, 27, 2–17. [Google Scholar] [CrossRef]
- Ferreira, M.A.; Andrade, F.; Mendes, R.N.; Paula, J. Use of satellite remote sensing for coastal conservation in the eastern african coast: Advantages and shortcomings. Eur. J. Remote Sens. 2012, 45, 293–304. [Google Scholar] [CrossRef]
- Namukose, M.; Msuya, F.E.; Ferse, S.C.A.; Slater, M.J.; Kunzmann, A. Growth performance of the sea cucumber Holothuria scabra and the seaweed Eucheuma denticulatum: Integrated mariculture and effects on sediment organic characteristics. Aquac. Environ. Interactactions 2016, 8, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Parra-luna, M.; Martín-pozo, L.; Hidalgo, F.; Zafra-Gómez, A. Common sea urchin (Paracentrotus lividus) and sea cucumber of the genus Holothuria as bioindicators of pollution in the study of chemical contaminants in aquatic media. A revision. Ecol. Indic. 2020, 113, 106185. [Google Scholar] [CrossRef]
- Mezali, K.; Soualili, D.L. The Ability of Holothurians to Select Sediment Particles and Organic Matter. SPC Beche Mer Inf. Bull. 2013, 1, 38–43. [Google Scholar]
- Kovos, D.; Karagiannis, G. Field study on the “National Marine Park” of Alonissos, Greece. J. Tour. Res. 2018, 19, 243–253. [Google Scholar]
- Karamanlidis, A.A.; Androukaki, E.; Adamantopoulou, S.; Chatzispyrou, A.; Johnson, W.M.; Kotomatas, S.; Papadopoulos, A.; Paravas, V.; Paximadis, G.; Pires, R.; et al. Assessing Accidental Entanglement as a Threat to the Mediterranean Monk Seal Monachus Monachus. Endanger. Species Res. 2008, 5, 205–213. [Google Scholar] [CrossRef]
- Oikonomou, Z.-S.; Dikou, A. Integrating Conservation and Development at the National Marine Park of Alonissos, Northern Sporades, Greece: Perception and Practice. Environ. Manag. 2008, 42, 847–866. [Google Scholar] [CrossRef]
- Konaxis, I. Alonissos Island and the Northern Sporades Marine National Park as a Strategic Socio-Economic Node for The Culture of the Aegean Sea. Am. Res. J. Humanit. Soc. Sci. 2020, 3, 49–53. [Google Scholar]
- Trivourea, M.N.; Karamanlidis, A.A.; Tounta, E.; Dendrinos, P.; Kotomatas, S. People and the Mediterranean Monk Seal (Monachus monachus): A Study of the Socioeconomic Impacts of the National Marine Park of Alonissos, Northern Sporades, Greece. Aquat. Mamm. 2011, 37, 305–318. [Google Scholar] [CrossRef] [Green Version]
- Androulidakis, Y.S.; Krestenitis, Y.N.; Psarra, S. Coastal upwelling over the North Aegean Sea: Observations and simulations. Cont. Shelf Res. 2017, 149, 32–51. [Google Scholar] [CrossRef]
- Poulos, S.E.; Drakopoulos, P.G.; Collins, M.B. Seasonal variability in sea surface oceanographic conditions in the Aegean Sea (Eastern Mediterranean): An overview. J. Mar. Syst. 1997, 13, 225–244. [Google Scholar] [CrossRef]
- Al-Wassai, F.A.; Kalyankar, N.V. Major Limitations of Satellite images. J. Glob. Res. Comput. Sci. 2013, 4, 51–59. [Google Scholar]
- Gao, J. Bathymetric mapping by means of remote sensing: Methods, accuracy and limitations. Prog. Phys. Geogr. 2009, 33, 103–116. [Google Scholar] [CrossRef]
- Lawson, K.; Larson, N.G. CTD. In Encyclopedia of Ocean Sciences; Steele, J.H., Ed.; Sea-Bird Electronics Inc.: Bellevue, DC, USA, 2001; pp. 579–588. ISBN 9780122274305. [Google Scholar]
- Fingas, M. Remote Sensing for Marine Management, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2018; ISBN 9780128050521. [Google Scholar]
- Lotfinasabasl, S.; Gunale, V.R.; Khosroshahi, M. Applying geographic information systems and remote sensing for water quality assessment of mangrove forest. Acta Ecol. Sin. 2018, 38, 135–143. [Google Scholar] [CrossRef]
- Cai, J.-Q.; Liu, X.-M.; Gao, Z.-J.; Li, L.-L.; Wang, H. Chlorophylls derivatives: Photophysical properties, assemblies, nanostructures and biomedical applications. Mater. Today 2021, 45, 77–92. [Google Scholar] [CrossRef]
- Nazeer, M.; Nichol, J.E. Development and application of a remote sensing-based Chlorophyll-a concentration prediction model for complex coastal waters of Hong Kong. J. Hydrol. 2016, 532, 80–89. [Google Scholar] [CrossRef]
- Yu, X.; Shen, J. A data-driven approach to simulate the spatiotemporal variations of chlorophyll-a in Chesapeake Bay. Ocean Model. 2021, 159, 101748. [Google Scholar] [CrossRef]
- Chawla, I.; Karthikeyan, L.; Mishra, A.K. A review of remote sensing applications for water security: Quantity, quality, and extremes. J. Hydrol. 2020, 585, 124826. [Google Scholar] [CrossRef]
- Gholizadeh, M.H.; Melesse, A.M.; Reddi, L. A Comprehensive Review on Water Quality Parameters Estimation Using Remote Sensing Techniques. Sensors 2016, 16, 1298. [Google Scholar] [CrossRef] [Green Version]
- Bennion, D.H.; Warner, D.M.; Esselman, P.C.; Hobson, B.; Kieft, B. A comparison of chlorophyll a values obtained from an autonomous underwater vehicle to satellite-based measures for Lake Michigan. J. Great Lakes Res. 2019, 45, 726–734. [Google Scholar] [CrossRef]
- Wei, G.F.; Tang, D.L.; Wang, S. Distribution of chlorophyll and harmful algal blooms (HABs): A review on space based studies in the coastal environments of Chinese marginal seas. Adv. Space Res. 2008, 41, 12–19. [Google Scholar] [CrossRef]
- Rodríguez-López, L.; Duran-Llacer, I.; González-Rodríguez, L.; Abarca-del-Rio, R.; Cárdenas, R.; Parra, O.; Martínez-Retureta, R.; Urrutia, R. Spectral analysis using LANDSAT images to monitor the chlorophyll-a concentration in Lake Laja in Chile. Ecol. Inform. 2020, 60, 101183. [Google Scholar] [CrossRef]
- Flores-cervantes, D.X.; Plata, D.L.; Macfarlane, J.K.; Reddy, C.M.; Gschwend, P.M. Black carbon in marine particulate organic carbon: Inputs and cycling of highly recalcitrant organic carbon in the Gulf of Maine. Mar. Chem. 2009, 113, 172–181. [Google Scholar] [CrossRef]
- Kharbush, J.J.; Close, H.G.; Van Mooy, B.A.S.; Arnosti, C.; Smittenberg, R.H.; Le Moigne, F.A.C.; Mollenhauer, G.; Scholz-Böttcher, B.; Obreht, I.; Koch, B.P.; et al. Particulate Organic Carbon Deconstructed: Molecular and Chemical Composition of Particulate Organic Carbon in the Ocean. Front. Mar. Sci. 2020, 7, 518. [Google Scholar] [CrossRef]
- McNichol, A.P.; Aluwihare, L.I. The Power of Radiocarbon in Biogeochemical Studies of the Marine Carbon Cycle: Insights from Studies of Dissolved and Particulate Organic Carbon (DOC and POC). Chem. Rev. 2007, 107, 443–466. [Google Scholar] [CrossRef]
- Son, B.Y.; Gardner, W.D.; Mishonov, A.V.; Jo, M. Multispectral remote-sensing algorithms for particulate organic carbon (POC): The Gulf of Mexico. Remote Sens. Environ. 2009, 113, 50–61. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Lei, S.; Bi, S.; Li, Y.; Lyu, H.; Xu, J. Tracking spatio-temporal dynamics of POC sources in eutrophic lakes by remote sensing Tracking spatio-temporal dynamics of POC sources in eutrophic lakes by remote sensing. Water Res. 2019, 168, 115162. [Google Scholar] [CrossRef]
- Yakushev, E.V.; Wallhead, P.; Renaud, P.E.; Ilinskaya, A.; Protsenko, E.; Yakubov, S.; Pakhomova, S.; Sweetman, A.K.; Dunlop, K.; Berezina, A.; et al. Understanding the Biogeochemical Impacts of Fish Farms Using a Benthic-Pelagic Model. Water 2020, 12, 2384. [Google Scholar] [CrossRef]
- Noroi, G.Á.; Glud, R.N.; Gaard, E.; Simonsen, K. Environmental Impacts of Coastal Fish Farming: Carbon and Nitrogen Budgets for Trout Farming in Kaldbaksfjørour (Faroe Islands). Mar. Ecol. Prog. Ser. 2011, 431, 223–241. [Google Scholar]
- Guangjia, J.; Ronghua, M.; Loiselle, S.A.; Duan, H.; Su, W.; Huang, C.; Jie Yang, W.Y. Remote sensing of particulate organic carbon dynamics in a eutrophic lake (Taihu Lake, China). Sci. Total Environ. 2015, 532, 245–254. [Google Scholar] [CrossRef]
- Djurhuus, A.; Read, J.F.; Rogers, A.D. The spatial distribution of particulate organic carbon and microorganisms on seamounts of the South West Indian Ridge. Deep Sea Res. Part II 2017, 136, 73–84. [Google Scholar] [CrossRef]
- Legendre, L.; Michaud, J. Chlorophyll a to estimate the particulate organic carbon available as food to large zooplankton in the euphotic zone of oceans. J. Plankton Res. 1999, 21, 2067–2083. [Google Scholar] [CrossRef] [Green Version]
- Abou Samra, R.M.; El-Gammal, M.; Eissa, R. Oceanographic Factors of Oil Pollution Dispersion Offshore the Nile Delta (Egypt) Using GIS. Environ. Sci. Pollut. Res. 2021, 28, 25830–25843. [Google Scholar] [CrossRef] [PubMed]
- Siepak, J. Total Organic Carbon (TOC) as a Sum Parameter of Water Pollution in Selected Polish Rivers (Vistula, Odra, and Warta). Acta Hydrochim. Hydrobiol. 1999, 27, 282–285. [Google Scholar] [CrossRef]
- Chaithanya, M.S.; Das, B.; Vidya, R. Assessment of metals pollution and subsequent ecological risk in water, sediments and vegetation from a shallow lake: A case study from Ranipet industrial town, Tamil Nadu, India. Int. J. Environ. Anal. Chem. 2021, 1–18. [Google Scholar] [CrossRef]
- Minnett, P.J.; Brown, O.B.; Evans, R.H.; Key, E.L.; Kearns, E.J.; Kilpatrick, K.; Kumar, A.; Maillet, K.A.; Szczodrak, G. Sea-Surface Temperature Measurements from the Moderate-Resolution Imaging Spectroradiometer (MODIS) on Aqua and Terra. Int. Geosci. Remote Sens. Symp. 2004, 7, 4576–4579. [Google Scholar] [CrossRef]
- Gmelin World Register of Marine Species (WoRMS). Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=125182 (accessed on 9 August 2022).
- Rakaj, A.; Fianchini, A.; Boncagni, P.; Scardi, M.; Cataudella, S. Artificial Reproduction of Holothuria Polii: A New Candidate for Aquaculture. Aquaculture 2019, 498, 444–453. [Google Scholar] [CrossRef]
- Tolon, M.T.; Engin, S. Gonadal Development of the Holothurian Holothuria Polii (Delle Chiaje, 1823) in Spawning Period at the Aegean Sea (Mediterranean Sea). Ege J. Fish. Aquat. Sci. 2019, 36, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Toscano, A.; Cirino, P. First Evidence of Artificial Fission in Two Mediterranean Species of Holothurians: Holothuria Tubulosa and Holothuria Polii. Turk. J. Fish. Aquat. Sci. 2018, 18, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
- Slimane-tamacha, F.; Soualili, D.L.; Mezali, K. Reproductive biology of Holothuria (Roweothuria) poli (Holothuroidea: Echinodermata) from Oran Bay, Algeria. SPC Beche Mer Inf. Bull. 2019, 39, 47–53. [Google Scholar]
- Gonzalez-Wanguemert, M.; Valente, S.; Aydin, M. Effects of fishery protection on biometry and genetic structure of two target sea cucumber species from the Mediterranean Sea. Hydrobiologia 2015, 743, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Simunovic, A.; Grubelic, I. A contribution to the knowledge of the species Holothuria tubulosa GMELIN, 1788 (Holothuria, Echinodermata) in the coastal area of the central eastern Adriatic. Acta Adriat. 1998, 39, 13–23. [Google Scholar]
- Neofitou, N.; Lolas, A.; Ballios, I.; Skordas, K.; Tziantziou, L.; Vafidis, D. Contribution of Sea Cucumber Holothuria Tubulosa on Organic Load Reduction from Fish Farming Operation. Aquaculture 2019, 501, 97–103. [Google Scholar] [CrossRef]
- Antoniadou, C.; Vafidis, D. Population structure of the traditionally exploited holothurian Holothuria tubulosa in the south Aegean Sea. Cah. Biol. Mar. 2011, 52, 171–175. [Google Scholar]
- Kazanidis, G.; Lolas, A.; Vafidis, D. Reproductive cycle of the traditionally exploited sea cucumber Holothuria tubulosa (Holothuroidea: Aspidochirotida) in Pagasitikos Gulf, western Aegean Sea, Greece. Turk. J. Zool. 2014, 38, 306–315. [Google Scholar] [CrossRef]
- Kazanidis, G.; Antoniadou, C.; Lolas, A.P.; Neofitou, N.; Vafidis, D.; Chintiroglou, C.; Neofitou, C. Population Dynamics and Reproduction of Holothuria Tubulosa (Holothuroidea: Echinodermata) in the Aegean Sea. J. Mar. Biol. Assoc. UK 2010, 90, 895–901. [Google Scholar] [CrossRef]
- Despalatovic, M.; Grubelic, I.; Simunovic, A.; Antolic, B.; Zuljevic, A. Reproductive biology of the holothurian Holothuria tubulosa (Echinodermata) in the Adriatic Sea. Mar. Biol. Assoc. UK 2004, 84, 409–414. [Google Scholar] [CrossRef]
- Yuval, B.; Sudai, L.; Ziv, Y. Abundance and Diversity of Holothuroids in Shallow Habitats of the Northern Red Sea. J. Mar. Biol. 2014, 2014, 631309. [Google Scholar] [CrossRef] [Green Version]
- Lampe-Ramdoo, K.; Moothien Pillay, R.; Conand, C. An assessment of holothurian diversity, abundance and distribution in the shallow lagoons of Mauritius. SPC Beche Mer Inf. Bull. 2014, 34, 17–24. [Google Scholar]
- Arsad, N.A.; Othman, R.; Raehanah, S.; Shaleh, M.; Abdullah, F.C.; Matsumoto, M.M. Effects of physicochemical parameters on the reproductive pattern of sea cucumber Holothuria scabra in Sabah. Songklanakarin J. Sci. Technol. 2020, 42, 109–116. [Google Scholar]
- Dissanayake, D.C.T.; Stefansson, G. Abundance and distribution of commercial sea cucumber species in the coastal waters of Sri Lanka. Aquat. Living Resour. 2011, 313, 303–313. [Google Scholar] [CrossRef]
- Collard, M.; Eeckhaut, I.; Dehairs, F.; Dubois, P. Acid–Base Physiology Response to Ocean Acidification of Two Ecologically and Economically Important Holothuroids from Contrasting Habitats, Holothuria Scabra and Holothuria Parva. Environ. Sci. Pollut. Res. 2014, 21, 13602–13614. [Google Scholar] [CrossRef]
- Herna, C.; Clemente, S.; Tuya, F. Is there a link between the type of habitat and the patterns of abundance of holothurians in shallow rocky reefs? Hydrobiologia 2006, 571, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Aydin, M. Density and Biomass of Commercial Sea Cucumber Species Relative to Depth in the Northern Aegean Sea. Thalass. Int. J. Mar. Sci. 2019, 35, 541–550. [Google Scholar] [CrossRef]
- Nazari-Sharabian, M.; Ahmad, S.; Moses, K. Climate Change and Eutrophication: A Short Review. Eng. Technol. Appl. Sci. Res. 2018, 8, 3668–3672. [Google Scholar] [CrossRef]
- Nurdin, S.; Mustapha, M.A.; Lihan, T. The Relationship between Sea Surface Temperature and Chlorophyll-a Concentration in Fisheries Aggregation Area in the Archipelagic Waters of Spermonde Using Satellite Images. AIP Conf. Proc. 2013, 1571, 466–472. [Google Scholar] [CrossRef]
- Bulteel, P.; Jangoux, M.; Coulon, P. Biometry, Bathymetric Distribution, and Reproductive Cycle of the Holothuroid from Mediterranean Seagrass Beds. Mar. Ecol. 1992, 13, 53–62. [Google Scholar] [CrossRef]
- Vafeiadou, A.M.; Antoniadou, C.; Vafidis, D.; Fryganiotis, K.; Chintiroglou, C.; Density, K.; Sea, A. Density and Biometry of the Exploited Holothurian Holothuria Tubulosa at the Dodecanese, South Aegean Sea. Rapp. Comm. Int. Mer Medit. 2010, 39, 661. [Google Scholar]
- Lavitra, T.; Fohy, N.; Gestin, P.; Rasolofonirina, R.; Eeckhaut, I. Effect of water temperature on the survival and growth of endobenthic Holothuria scabra (Echinodermata: Holothuroidea) juveniles reared in outdoor ponds. SPC Beche Mer Inf. Bull. 2010, 30, 25–28. [Google Scholar]
- Yang, H.; Yuan, X.; Zhou, Y.; Mao, Y.; Zhang, T.; Liu, Y. Effects of body size and water temperature on food consumption and growth in the sea cucumber Apostichopus japonicus (Selenka) with special reference to aestivation. Aquac. Res. 2005, 36, 1085–1092. [Google Scholar] [CrossRef]
- Marquet, N.; Conand, C.; Power, D.M.; Canário, A.V.M.; González-wangüemert, M. Sea cucumbers, Holothuria arguinensis and H. mammata, from the southern Iberian Peninsula: Variation in reproductive activity between populations from different habitats. Fish. Res. 2017, 191, 120–130. [Google Scholar] [CrossRef]
- Zamora, L.N.; Jeffs, A.G. Feeding, metabolism and growth in response to temperature in juveniles of the Australasian sea cucumber, Australostichopus mollis. Aquaculture 2012, 358–359, 92–97. [Google Scholar] [CrossRef]
- Lampe, K. Holothurian density, distribution and diversity comparing sites with different degrees of exploitation in the shallow lagoons of Mauritius. SPC Beche Mer Inf. Bull. 2013, 33, 23–29. [Google Scholar]
- Seeruttun, R.; Appadoo, C.; Laxminarayana, A.; Codabaccus, B. A Study on the Factors Influencing the Growth and Survival of Juvenile Sea Cucumber, Holothuria atra, under Laboratory Conditions. Univ. Maurit. Res. J. 2008, 14, 1–15. [Google Scholar]
- Tolon, T. Effect of salinity on growth and survival of the juvenile sea cucumbers Holothuria tubulosa (Gmelin, 1788) and Holothuria poli (Delle Chiaje, 1923). Fresenius Environ. Bull. 2017, 26, 3930–3935. [Google Scholar]
- Darmaraki, S.; Somot, S.; Sevault, F.; Nabat, P.; Cabos Narvaez, W.D.; Cavicchia, L.; Djurdjevic, V.; Li, L.; Sannino, G.; Sein, D.V. Future evolution of Marine Heatwaves in the Mediterranean Sea. Clim. Dyn. 2019, 53, 1371–1392. [Google Scholar] [CrossRef]
- Xiong, X.; Salomonson, V.V.; Barnes, W.L.; Guenther, B.; Xie, X.; Sun, J. An overview of terra MODIS reflective solar bands on-orbit calibration. In Proceedings of the 2006 IEEE International Symposium on Geoscience and Remote Sensing, Denver, CO, USA, 31 July–4 August 2006; pp. 1103–1106. [Google Scholar] [CrossRef]
- Pisano, A.; Marullo, S.; Artale, V.; Falcini, F.; Yang, C.; Leonelli, F.E.; Santoleri, R.; Nardelli, B.B. New evidence of Mediterranean climate change and variability from Sea Surface Temperature observations. Remote Sens. 2020, 12, 132. [Google Scholar] [CrossRef] [Green Version]
- Pastor, F.; Valiente, J.A.; Khodayar, S. A warming Mediterranean: 38 years of increasing sea surface temperature. Remote Sens. 2020, 12, 2687. [Google Scholar] [CrossRef]
- Pastor, F.; Valiente, J.A.; Palau, J.L. Sea Surface Temperature in the Mediterranean: Trends and Spatial Patterns (1982–2016). Pure Appl. Geophys. 2018, 175, 4017–4029. [Google Scholar] [CrossRef] [Green Version]
- Tyrlis, E.; Lelieveld, J. Climatology and Dynamics of the Summer Etesian Winds over the Eastern Mediterranean. J. Atmos. Sci. 2013, 70, 3374–3396. [Google Scholar] [CrossRef]
- Poupkou, A.; Zanis, P.; Nastos, P.; Papanastasiou, D.; Melas, D.; Tourpali, K.; Zerefos, C. Present climate trend analysis of the Etesian winds in the Aegean Sea. Theor. Appl. Climatol. 2011, 106, 459–472. [Google Scholar] [CrossRef]
- Anagnostopoulou, C.; Zanis, P.; Katragkou, E.; Tegoulias, I.; Tolika, K. Recent past and future patterns of the Etesian winds based on regional scale climate model simulations. Clim. Dyn. 2014, 42, 1819–1836. [Google Scholar] [CrossRef]
- Coulon, P.; Jangoux, M. Feeding rate and sediment reworking by the holothuroid Holothuria tubulosa (Echinodermata) in a Mediterranean seagrass bed off Ischia Island, Italy. Mar. Ecol. Prog. Ser. 1993, 92, 201–204. [Google Scholar] [CrossRef]
- Ru, X.; Zhang, L.; Liu, S.; Yang, H. Plasticity of Respiratory Function Accommodates High Oxygen Demand in Breeding Sea Cucumbers. Front. Physiol. 2020, 11, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emiroğlu, D.; Günay, D. The effect of sea cucumber Holothuria tubulosa G. 1788 on nutrient and organic matter contents of bottom sediment of oligotrophy and hypereutrophic shores. Fresenius Environ. Bull. 2007, 16, 290–294. [Google Scholar]
- Günay, D.; Emiroğlu, D.; Tolon, T.; Özden, O.; Saygi, H. Growth and Survival Rate of Juvenile Sea Cucumbers (Holothuria Tubulosa, Gmelin, 1788) at Various Temperatures. Turk. J. Fish. Aquat. Sci. 2015, 15, 533–541. [Google Scholar] [CrossRef]
- Gao, Q.F.; Wang, Y.; Dong, S.; Sun, Z.; Wang, F. Absorption of different food sources by sea cucumber Apostichopus japonicus (Selenka) (Echinodermata: Holothuroidea): Evidence from carbon stable isotope. Aquaculture 2011, 319, 272–276. [Google Scholar] [CrossRef]
- Ren, Y.; Dong, S.; Wang, F.; Gao, Q.; Tian, X.; Liu, F. Sedimentation and sediment characteristics in sea cucumber Apostichopus japonicus (Selenka) culture ponds. Aquac. Res. 2010, 42, 14–21. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SST [Daytime] | SST [Night-Time] | Chl-a | POC | |||||
|---|---|---|---|---|---|---|---|---|
| Per Month | °C | Per Month | °C | Per Month | Mg/m3 | Per Month | Mg/m3 | |
| min | 0.002 | 0.47 | 0.0040 | 0.96 | 0.000027 | 0.0064 | 0.0035 | 0.8 |
| max | 0.005 | 1.09 | 0.0053 | 1.28 | 0.000138 | 0.0331 | 0.0722 | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christou, P.; Domenikiotis, C.; Neofitou, N.; Vafidis, D. Study of the Spatiotemporal Variability of Oceanographic Parameters and Their Relationship to Holothuria Species Abundance in a Marine Protected Area of the Mediterranean Using Satellite Imagery. Remote Sens. 2022, 14, 5946. https://doi.org/10.3390/rs14235946
Christou P, Domenikiotis C, Neofitou N, Vafidis D. Study of the Spatiotemporal Variability of Oceanographic Parameters and Their Relationship to Holothuria Species Abundance in a Marine Protected Area of the Mediterranean Using Satellite Imagery. Remote Sensing. 2022; 14(23):5946. https://doi.org/10.3390/rs14235946
Chicago/Turabian StyleChristou, Panteleimon, Christos Domenikiotis, Nikos Neofitou, and Dimitris Vafidis. 2022. "Study of the Spatiotemporal Variability of Oceanographic Parameters and Their Relationship to Holothuria Species Abundance in a Marine Protected Area of the Mediterranean Using Satellite Imagery" Remote Sensing 14, no. 23: 5946. https://doi.org/10.3390/rs14235946