
Towards Sustainable Livestock Production: Estimation of Methane Emissions and Dietary Interventions for Mitigation

 , ,
, ,  and
and
Abstract
:
1. Introduction
2. Global Climate Change and the Role of Methane
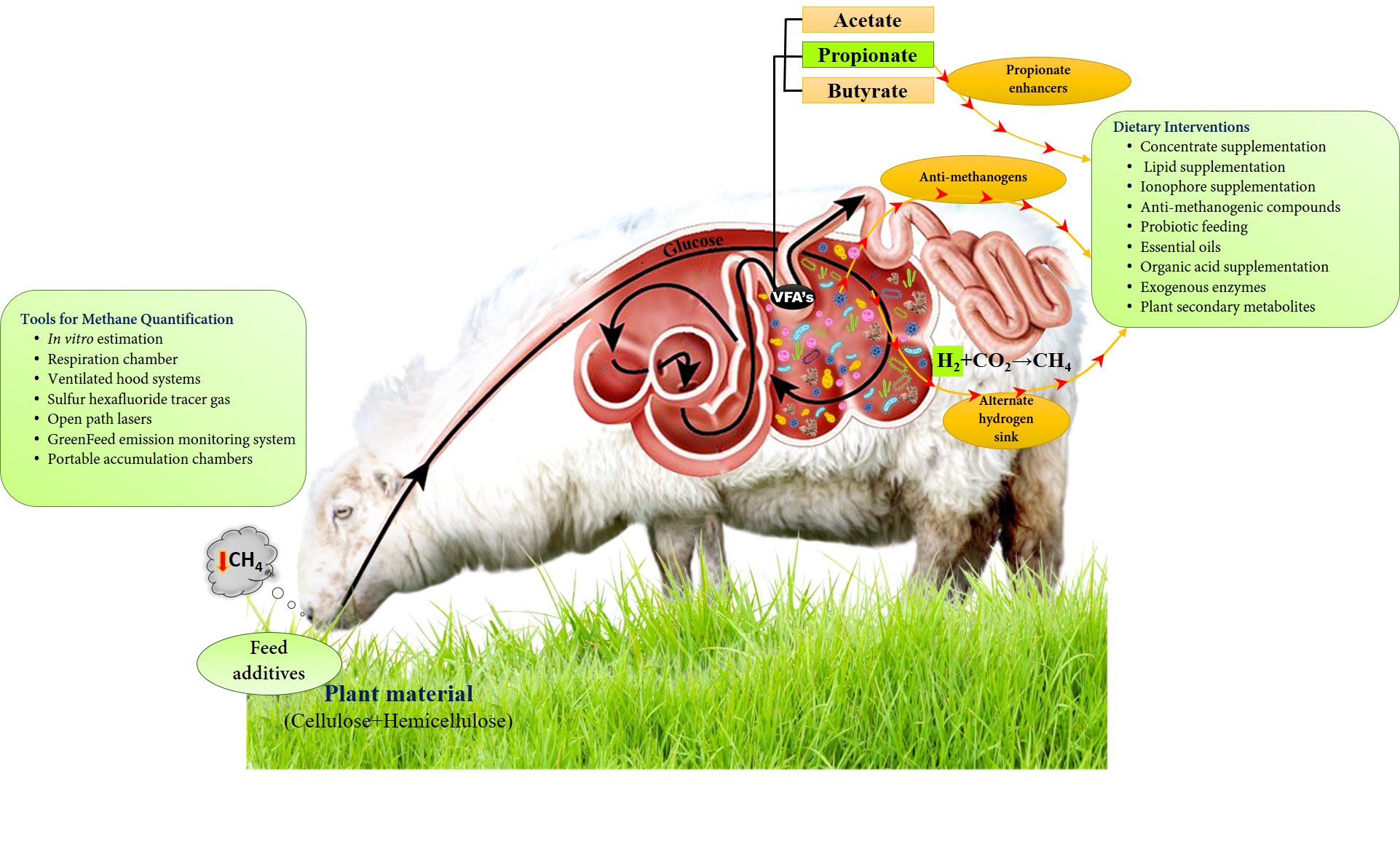
3. Mechanism of Methane Production in the Rumen
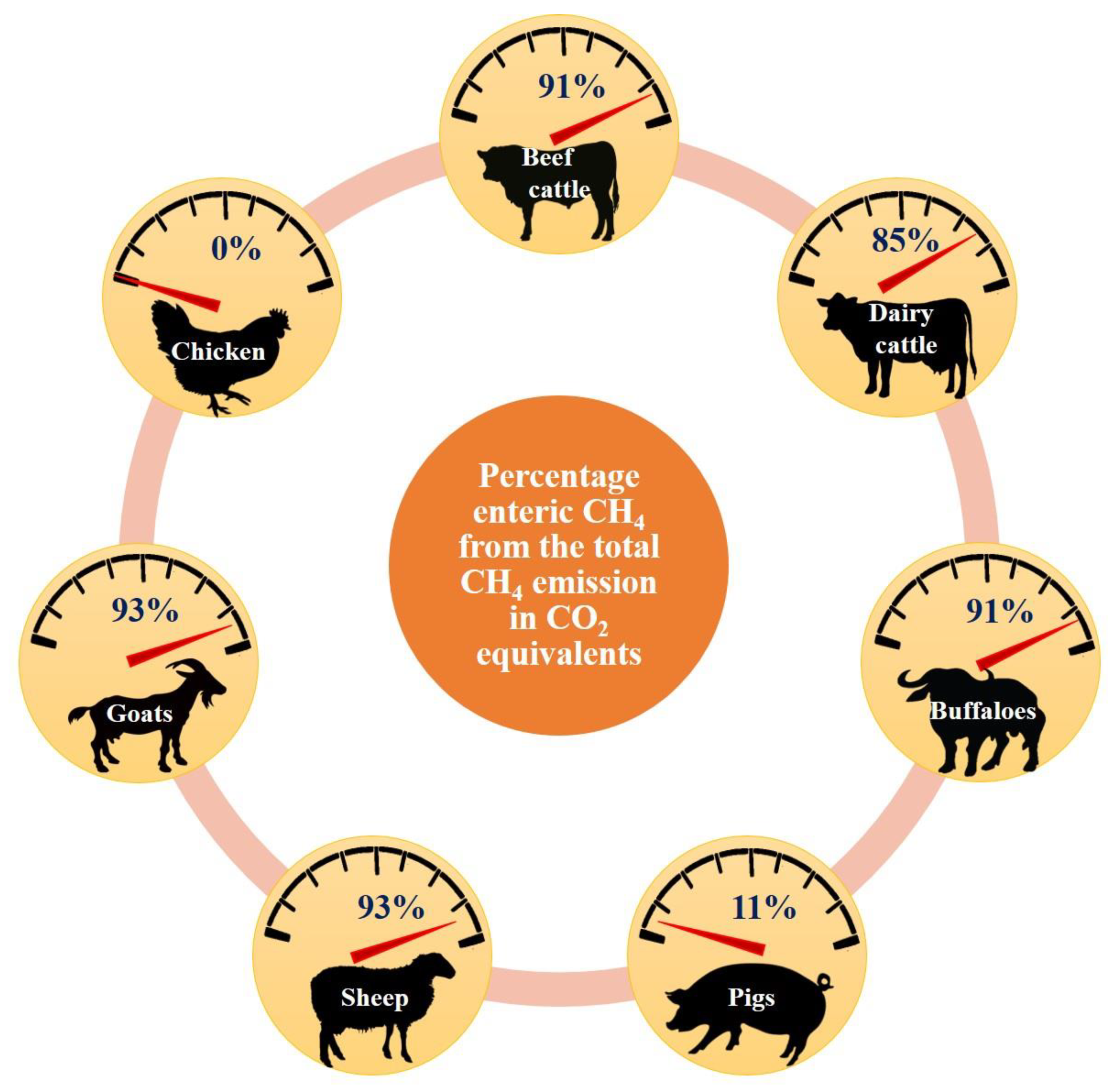
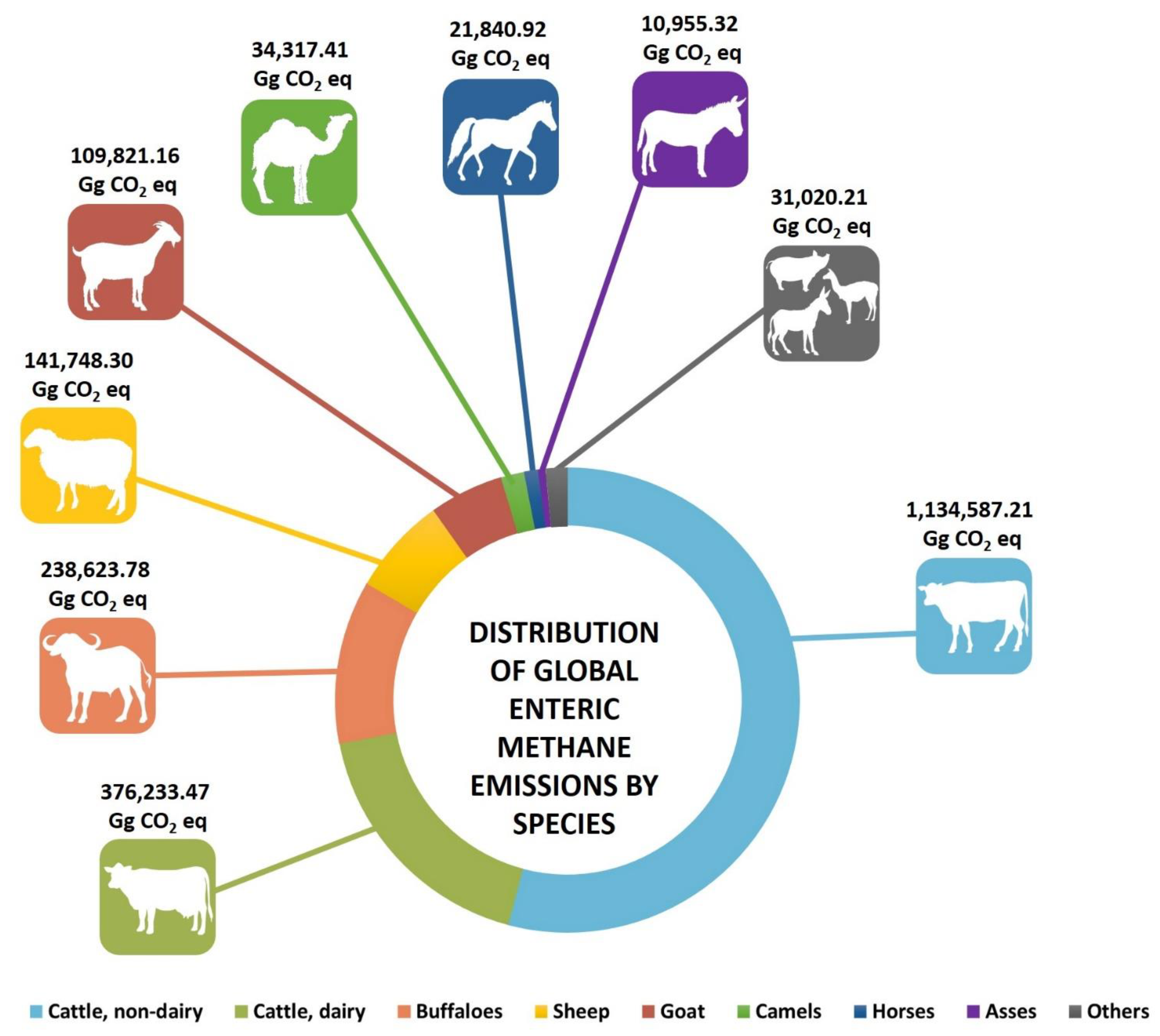
4. Estimates of Enteric Methane Production: How Much Does Each Species Contribute?
5. Methods Used to Quantify Methane Production
5.1. In Vitro Estimation
5.2. Respiration Chamber
5.3. Ventilated Hood Systems
5.4. Sulfur Hexafluoride Tracer Gas
5.5. Open-Path Lasers
5.6. GreenFeed Emission Monitoring System
5.7. Portable Accumulation Chambers
6. Nutritional Interventions as One of the Important Methane Mitigation Options
6.1. Concentrate Supplementation
6.2. Lipid Supplementation
6.3. Ionophore Supplementation
6.4. Anti-Methanogenic Compounds
6.5. Probiotic Feeding
6.6. Essential Oils
6.7. Organic Acid Supplementation
6.8. Exogenous Enzymes
6.9. Plant Secondary Metabolites

{kind=link}
{kind=link}
{kind=link}
| Feed Additives | Dosage | Species | Impact on CH4 | Current Feasibility | Reference |
|---|---|---|---|---|---|
| Concentrate feed containing vitamins and mineral supplement | 6 kg/day | Cow | ↑ CH4 (g/day) | feasible | [176] |
| Ground corn | 3.2 kg/day | Cow | ↓ CH4 yield by 7.3 g/kg DMI | feasible | [177] |
| Concentrate feed containing maize, rapeseed meal, soybean, Molaferm, and Megalac, etc. | 6 kg/day | Cow | No effect (g/kgDMI) but ↓ CH4 (g/kg Energy corrected milk) | feasible | [178] |
| Concentrate feed containing barley, beet pulp, soybean meal, maize meal, molasses, vitamins, and minerals | 0.5 kg/day | Lamb | No effect (CH4 g/kg DMI) | feasible | [179] |
| Concentrate feed mixture | 18.1% of DM | Cow | ↓ CH4 (g/day and g/kg DMI) | feasible | [180] |
| Concentrate feed containing maize, deoiled mustard cake, soybean meal, wheat bran, rice bran, mineral mixture, and salt etc. | 15% high ME content (2.82 Mcal/kg) | buffalo | ↓ CH4 g/day, g/kg DM intake) | feasible | [181] |
| Coconut oil | 4% | Goat | 34% ↓ CH4 emission | Somewhat feasible | [182] |
| Coconut oil | 2% | Goat | More than 50% ↓ CH4 emission | Somewhat feasible | [183] |
| Soybean oil | 4% | Goat | 32% ↓ CH4 emission | Somewhat feasible | [182] |
| Soybean oil | 50 g/kg DM | Sheep | 35.8% ↓ CH4 emission | Somewhat feasible | [184] |
| Corn oil | 30 g/kg DM | Goat | 15.1% ↓ CH4 emission (g/kg DMI) | feasible | [185] |
| Corn oil | 5% | Cattle | ~30% ↓ CH4 emission | feasible | [186] |
| Lasalocid | 200 mg/hd/d | Cattle | 30.91% ↓ CH4 production (g/kg DMI) | feasible | [187] |
| Monensin | 30 mg/kg | Steer | 16.67% ↓ CH4 (MJ/100 MJ GE intake) | Currently not feasible | [188] |
| Monensin | 22 mg/kg | Goat | 28% ↓ CH4 emission | Currently not feasible | [182] |
| Monensin | 0.6 mg/kg of body weight | Buffalo | 8–9%↓ CH4 emission | Currently not feasible | [118] |
| Nitrate | 11 g/kg DM | Cow | ↓ CH4 by 8% | Not permitted in some countries | [101] |
| Nitrate | 23 g/kg DM | Cow | ↓ CH4 by 15% | Not permitted in some countries | [101] |
| Ethyl-3-NOP | 50 and 500 mg/animal per day | Sheep | ↓ CH4 by 29% (L/kg of DMI) | Not permitted in some countries | [125] |
| 3NOP | 60 mg of 3NOP/kg DM | Cow | ↓ CH4 by 31% | Not permitted in some countries | [124] |
| Bacillus licheniformis | 2.5 × 108 colony forming units (CFU) | Sheep | ↓ CH4 by 6% | Currently not economically feasible | [136] |
| Bacillus licheniformis | 2.5 × 109 CFUs | Sheep | ↓ CH4 by 12% | Currently not economically feasible | [136] |
| Saccharomyces cerevisiae | (1.2–2.3) × 107 CFU/g | Sheep | ↓ CH4 by 10% (L/day) | Currently not economically feasible | [189] |
| Leuconostoc mesenteroides | (1.5–1.8) × 109 CFU/g | Sheep | No effect | Currently not economically feasible | [189] |
| Orange leaves | TMR | Goat | ↓ CH4 by 32% (g/day) | feasible | [190] |
| Citrus essential oil | 0. 0.8 and 1.6 mL/L | sheep | ↓ CH4 | feasible | [144] |
| Commercial essential oil blend | 1 g/day | Cow | ↓ CH4 by 6% (g/day) | feasible | [145] |
| Encapsulated fumaric acid | 117 g EFA/kg | Lamb | ↓ CH4 by 76% (L/day) | feasible | [191] |
| Fumaric acid | 100 g FA and 17 g partially hydrogenated vegetable oil/kg | Lamb | ↓ CH4 by 62% (L/day) | Currently not economically feasible | [191] |
| Dl-malic acid | 7.5% on a DM basis | Beef cattle | ↓ CH4 by 9% (L/day) | Currently not economically feasible | [154] |
| Cellulase | 10,000 IU/g | Goat | No effect | Currently not economically possible | [192] |
| Cellulose/xylanase | 7000 IU/g of cellulase and 5000 IU/g of xylanase | Goat | No effect | Currently not economically feasible | [192] |
| Leucaena leucocephala | 350 g/kg DM | sheep | ↓ CH4 by 14.1% g/kg DMI | [193] | |
| White grape marc | 5.0 kg DM | Cow | ↓ CH4 by 15% g/kg DMI | feasible | [194] |
| Red grape marc | 5.0 kg DM | Cow | ↓ CH4 by 15% g/kg DMI | feasible | [194] |
| Willow fodder (Salix spp.) | 12 g CT kg/DMI | Sheep | ↓ CH4 by 19% (g/kg BW0.75/day) | feasible | [195] |
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UN-EPA. Global Anthropogenic Non-CO2 Greenhouse Gas Emissions: 1990–2030; Climate Change Division U.S. Environmental Protection Agency 1200 Pennsylvania Avenue NW: Washington, DC, USA, 2012.
- IPCC. Climate Change and Land, an Ipcc Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Inter Governmental Panel on Climate Change: Geneva, Switzerland, 2019; p. 41.
- Opio, C.; Gerber, P.; Mottet, A.; Falcucci, A.; Tempio, G.; MacLeod, M.; Vellinga, T.; Henderson, B.; Steinfeld, H. Greenhouse Gas Emissions from Ruminant Supply Chains–A Global Life Cycle Assessment; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Davison, T.M.; Black, J.L.; Moss, J.F. Red meat—An essential partner to reduce global greenhouse gas emissions. Anim. Front. 2020, 10, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Ammar, H.; Abidi, S.; Ayed, M.; Moujahed, N.; de Haro Martí, M.E.; Chahine, M.; Bouraoui, R.; López, S.; Cheikh M’hamed, H.; Hechlef, H. Estimation of tunisian greenhouse gas emissions from different livestock species. Agriculture 2020, 10, 562. [Google Scholar] [CrossRef]
- Giraldo, P.; Marín, A.; Giraldo, L.A. Effects of different feed additives on methane emissions from beef cattle. In Impacts on and Adaptation of Livestock Production Systems to Climate Change; Universidad Nacional de Colombia: Cundinamarca, Colombia, 2013; pp. 1386–1387. [Google Scholar]
- Stewart, E.K.; Beauchemin, K.A.; Dai, X.; MacAdam, J.W.; Christensen, R.G.; Villalba, J.J. Effect of tannin-containing hays on enteric methane emissions and nitrogen partitioning in beef cattle1. J. Anim. Sci. 2019, 97, 3286–3299. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Lee, C.; Pechtl, H.A.; Hettick, J.M.; Campler, M.R.; Pairis-Garcia, M.D.; Beauchemin, K.A.; Celi, P.; Duval, S.M. Effects of 3-nitrooxypropanol on enteric methane production, rumen fermentation, and feeding behavior in beef cattle fed a high-forage or high-grain diet. J. Anim. Sci. 2019, 97, 2687–2699. [Google Scholar] [CrossRef]
- DISER-Department of Industry; Energy and Resources. National greenhouse gas inventory quarterly update: September 2020. In Quarterly; Commonwealth of Australia: Sydney, Australia, 2021. [Google Scholar]
- Bhatta, R.; Malik, P.K.; Sejian, V. Enteric methane emission and reduction strategies in sheep. In Sheep Production Adapting to Climate Change; Sejian, V., Bhatta, R., Gaughan, J., Malik, P.K., Naqvi, S.M.K., Lal, R., Eds.; Springer: Singapore, 2017; pp. 291–305. [Google Scholar]
- Hammond, K.J.; Crompton, L.A.; Bannink, A.; Dijkstra, J.; Yáñez-Ruiz, D.R.; O’Kiely, P.; Kebreab, E.; Eugène, M.; Yu, Z.; Shingfield, K.J. Review of current in vivo measurement techniques for quantifying enteric methane emission from ruminants. Anim. Feed Sci. Technol. 2016, 219, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K. Recent advances in measurement and dietary mitigation of enteric methane emissions in ruminants. Front. Vet. Sci. 2016, 3, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pragna, P.; Chauhan, S.S.; Sejian, V.; Leury, B.J.; Dunshea, F.R. Climate change and goat production: Enteric methane emission and its mitigation. Animals 2018, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Min, B.R.; Solaiman, S.; Waldrip, H.M.; Parker, D.; Todd, R.W.; Brauer, D. Dietary mitigation of enteric methane emissions from ruminants: A review of plant tannins mitigation options☆. Anim. Nutr. 2020, 6, 231–246. [Google Scholar] [CrossRef]
- McCauley, J.I.; Labeeuw, L.; Jaramillo-Madrid, A.C.; Nguyen, L.N.; Nghiem, L.D.; Chaves, A.V.; Ralph, P.J. Management of enteric methanogenesis in ruminants by algal-derived feed additives. Curr. Pollut. Rep. 2020, 6, 1–18. [Google Scholar] [CrossRef]
- Baca-González, V.; Asensio-Calavia, P.; González-Acosta, S.; Pérez de la Lastra, J.M.; Morales de la Nuez, A. Are vaccines the solution for methane emissions from ruminants? A systematic review. Vaccines 2020, 8, 460. [Google Scholar] [CrossRef]
- IPCC. Global Warming of 1.5 °C-An Ipcc Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; IPCC: Geneva, Switzerland, 2018. [Google Scholar]
- IPCC. Synthesis report: Summary for policy makers. In Fifth Assessment Report (ar5); Pachauri, R.K., Riahi, K., Eds.; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014; p. 31. [Google Scholar]
- IPCC. Summary for policymakers ipcc special report on the ocean and cryosphere in a changing climate. In Ipcc Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.O., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Nicolai, M., Okem, A., Petzold, J., Rama, B., et al., Eds.; IPCC: Geneva, Switzerland, 2019; in press. [Google Scholar]
- Steffen, W.; Hughes, L.; Perkins, S. Heatwaves: Hotter, Longer, More Often; Climate Council of Australia: Sydney, Australia, 2014. [Google Scholar]
- Wuebbles, D.J.; Hayhoe, K. Atmospheric methane and global change. Earth-Sci. Rev. 2002, 57, 177–210. [Google Scholar] [CrossRef]
- Malyan, S.K.; Bhatia, A.; Kumar, A.; Gupta, D.K.; Singh, R.; Kumar, S.S.; Tomer, R.; Kumar, O.; Jain, N. Methane production, oxidation and mitigation: A mechanistic understanding and comprehensive evaluation of influencing factors. Sci. Total Environ. 2016, 572, 874–896. [Google Scholar] [CrossRef]
- Malyan, S.K.; Bhatia, A.; Kumar, S.S.; Fagodiya, R.K.; Pugazhendhi, A.; Duc, P.A. Mitigation of greenhouse gas intensity by supplementing with azolla and moderating the dose of nitrogen fertilizer. Biocatal. Agric. Biotechnol. 2019, 20, 101266. [Google Scholar] [CrossRef]
- EPA. Importance of Methane. In Global Methane Initiative; United States Environmental Protection Agency: Washington, DC, USA, 2016. Available online: https://www.epa.gov/gmi/importance-methane#:~:text=Methane%20is%20also%20a%20greenhouse,%2Dinfluenced)%20and%20natural%20sources.&text=Methane%20is%20more%20than%2025,trapping%20heat%20in%20the%20atmosphere (accessed on 15 July 2020).
- Brasseur, G.P.; Schultz, M.; Granier, C.; Saunois, M.; Diehl, T.; Botzet, M.; Roeckner, E.; Walters, S. Impact of climate change on the future chemical composition of the global troposphere. J. Clim. 2006, 19, 3932–3951. [Google Scholar] [CrossRef] [Green Version]
- Hook, S.E.; Wright, A.D.; McBride, B.W. Methanogens: Methane producers of the rumen and mitigation strategies. Archaea 2010, 2010, 945785. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.E.; Ward, G.M. Estimates of animal methane emissions. Environ. Monit. Assess. 1996, 42, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Huws, S.A.; Creevey, C.J.; Oyama, L.B.; Mizrahi, I.; Denman, S.E.; Popova, M.; Muñoz-Tamayo, R.; Forano, E.; Waters, S.M.; Hess, M. Addressing global ruminant agricultural challenges through understanding the rumen microbiome: Past, present, and future. Front. Microbiol. 2018, 9, 2161. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2019, 10, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Fagodiya, R.K.; Hiloidhari, M.; Dahiya, R.P.; Kumar, A. Methane production and estimation from livestock husbandry: A mechanistic understanding and emerging mitigation options. Sci. Total Environ. 2020, 709, 136135. [Google Scholar] [CrossRef] [PubMed]
- Moss, A.R.; Jouany, J.-P.; Newbold, J. Methane Production by Ruminants: Its Contribution to Global Warming; Annales de zootechnie, 2000; EDP Science: London, UK, 2000; pp. 231–253. [Google Scholar]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 1–15. [Google Scholar] [CrossRef]
- Tapio, I.; Snelling, T.J.; Strozzi, F.; Wallace, R.J. The ruminal microbiome associated with methane emissions from ruminant livestock. J. Anim. Sci. Biotechnol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Van Lingen, H.J.; Plugge, C.M.; Fadel, J.G.; Kebreab, E.; Bannink, A.; Dijkstra, J. Thermodynamic driving force of hydrogen on rumen microbial metabolism: A theoretical investigation. PLoS ONE 2016, 11, e0161362. [Google Scholar]
- FAO. Five Practical Actions towards Low-Carbon Livestock; Animal Production and Health Division: Rome, Italy, 2019. [Google Scholar]
- FAOSTAT. Fao Statistical Data Base; FAOSTAT: Rome, Italy, 2018. [Google Scholar]
- Gerber, P.J.; Steinfeld, H.; Henderson, B.; Mottet, A.; Opio, C.; Dijkman, J.; Falcucci, A.; Tempio, G. Tackling Climate Change through Livestock: A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013. [Google Scholar]
- Grossi, G.; Goglio, P.; Vitali, A.; Williams, A.G. Livestock and climate change: Impact of livestock on climate and mitigation strategies. Anim. Front. 2019, 9, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Global Livestock Environmental Assessment Model; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018; Volume 2, p. 109. [Google Scholar]
- Goopy, J.P.; Chang, C.; Tomkins, N. A comparison of methodologies for measuring methane emissions from ruminants. In Methods for Measuring Greenhouse Gas Balances and Evaluating Mitigation Options in Smallholder Agriculture; Rosenstock, T.S., Rufino, M.C., Butterbach-Bahl, K., Wollenberg, L., Richards, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 97–117. [Google Scholar]
- Brouček, J. Methods of methane measurement in ruminants. Slovak J. Anim. Sci. 2014, 47, 51–60. [Google Scholar]
- Alvarez Hess, P.S.; Eckard, R.J.; Jacobs, J.L.; Hannah, M.C.; Moate, P.J. Comparison of five methods for the estimation of methane production from vented in vitro systems. J. Sci. Food Agric. 2019, 99, 109–116. [Google Scholar] [CrossRef]
- Russo, V.; Jacobs, J.; Hannah, M.; Moate, P.; Dunshea, F.; Leury, B. In vitro evaluation of the methane mitigation potential of a range of grape marc products. Anim. Prod. Sci. 2017, 57, 1437–1444. [Google Scholar] [CrossRef]
- Gonzalez-Rivas, P.; DiGiacomo, K.; Russo, V.; Leury, B.; Cottrell, J.; Dunshea, F. Feeding slowly fermentable grains has the potential to ameliorate heat stress in grain-fed wethers. J. Anim. Sci. 2016, 94, 2981–2991. [Google Scholar] [CrossRef]
- Hess, P.A.; Giraldo, P.; Williams, R.; Moate, P.; Beauchemin, K.; Eckard, R. A novel method for collecting gas produced from the in vitro ankom gas production system. J. Anim. Sci. 2016, 94, 570. [Google Scholar] [CrossRef]
- Ramin, M.; Huhtanen, P. Development of an in vitro method for determination of methane production kinetics using a fully automated in vitro gas system—A modelling approach. Anim. Feed Sci. Technol. 2012, 174, 190–200. [Google Scholar] [CrossRef]
- Dubois, B.; Tomkins, N.W.; Kinley, R.D.; Bai, M.; Seymour, S.; Paul, N.A.; de Nys, R. Effect of tropical algae as additives on rumen in vitro gas production and fermentation characteristics. Am. J. Plant Sci. 2013, 4, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Ramin, M.; Krizsan, S.; Jančík, F.; Huhtanen, P. Measurements of methane emissions from feed samples in filter bags or dispersed in the medium in an in vitro gas production system. J. Dairy Sci. 2013, 96, 4643–4646. [Google Scholar] [CrossRef]
- Bhatta, R.; Tajima, K.; Takusari, N.; Higuchi, K.; Enishi, O.; Kurihara, M. Comparison of Sulfur Hexafluoride Tracer Technique, Rumen Simulation Technique and In Vitro Gas Production Techniques for Methane Production from Ruminant Feeds; International Congress Series; Elsevier: Amsterdam, The Netherlands, 2006; pp. 58–61. [Google Scholar]
- Czerkawski, J.; Breckenridge, G. Design and development of a long-term rumen simulation technique (rusitec). Br. J. Nutr. 1977, 38, 371–384. [Google Scholar] [CrossRef] [Green Version]
- Kellner, O.J.; Goodwin, W. The Scientific Feeding of Animals; Duckworth: Richmond-upon-Thames, UK, 1913. [Google Scholar]
- Muñoz, C.; Yan, T.; Wills, D.; Murray, S.; Gordon, A. Comparison of the sulfur hexafluoride tracer and respiration chamber techniques for estimating methane emissions and correction for rectum methane output from dairy cows. J. Dairy Sci. 2012, 95, 3139–3148. [Google Scholar] [CrossRef] [PubMed]
- Schwarm, A.; Schweigel-Röntgen, M.; Kreuzer, M.; Ortmann, S.; Gill, F.; Kuhla, B.; Meyer, U.; Lohölter, M.; Derno, M. Methane emission, digestive characteristics and faecal archaeol in heifers fed diets based on silage from brown midrib maize as compared to conventional maize. Arch. Anim. Nutr. 2015, 69, 159–176. [Google Scholar] [CrossRef] [Green Version]
- Hammond, K.; Hoskin, S.; Burke, J.; Waghorn, G.; Koolaard, J.; Muetzel, S. Effects of feeding fresh white clover (trifolium repens) or perennial ryegrass (lolium perenne) on enteric methane emissions from sheep. Anim. Feed Sci. Technol. 2011, 166, 398–404. [Google Scholar] [CrossRef]
- Vanlierde, A.; Soyeurt, H.; Gengler, N.; Colinet, F.G.; Froidmont, E.; Kreuzer, M.; Grandl, F.; Bell, M.; Lund, P.; Olijhoek, D.W.; et al. Short communication: Development of an equation for estimating methane emissions of dairy cows from milk fourier transform mid-infrared spectra by using reference data obtained exclusively from respiration chambers. J. Dairy Sci. 2018, 101, 7618–7624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Green-Miller, A.R.; Shike, D.W. Integrity assessment of open-circuit respiration chambers for ruminant animal indirect calorimetry. Trans. Am. Soc. Agric. Biol. Eng. 2019, 62, 1185–1193. [Google Scholar] [CrossRef]
- Takahashi, J.; Chaudhry, A.; Beneke, R.; Young, B. An open-circuit hood system for gaseous exchange measurements in small ruminants. Small Rumin. Res. 1999, 32, 31–36. [Google Scholar] [CrossRef]
- Fernández Martínez, C.J.; López Luján, M.; Lachica, M. Low-cost mobile open-circuit hood system for measuring gas exchange in small ruminants: From manual to automatic recording. J. Agric. Sci. 2015, 153, 1302–1309. [Google Scholar] [CrossRef]
- Place, S.E.; Pan, Y.; Zhao, Y.; Mitloehner, F.M. Construction and operation of a ventilated hood system for measuring greenhouse gas and volatile organic compound emissions from cattle. Animals 2011, 1, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Fernández, C.; Gomis-Tena, J.; Hernández, A.; Saiz, J. An open-circuit indirect calorimetry head hood system for measuring methane emission and energy metabolism in small ruminants. Animals 2019, 9, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troy, S.; Rooke, J.; Duthie, C.; Ross, D.; Hyslop, J.; Roehe, R.; Waterhouse, T. Measurement of methane from finishing cattle fed either a forage-based or high concentrate diet from both feeder-mounted samplers and respiration chambers. In Proceedings of the Greenhouse Gases and Animal Agriculture Conference, Dublin, Ireland, 23–26 June 2013; Volume 4, p. 551. [Google Scholar]
- Hill, J.; McSweeney, C.; Wright, A.-D.G.; Bishop-Hurley, G.; Kalantar-Zadeh, K. Measuring methane production from ruminants. Trends Biotechnol. 2016, 34, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; McCrabb, G.; Nishida, T.; Indramanee, S.; Kurihara, M. Construction and operation of ventilated hood-type respiration calorimeters for in vivo measurement of methane production and energy partition in ruminants. In Measuring Methane Production from Ruminants; Springer: Berlin/Heidelberg, Germany, 2007; pp. 125–135. [Google Scholar]
- Kebreab, E.; Clark, K.; Wagner-Riddle, C.; France, J. Methane and nitrous oxide emissions from canadian animal agriculture: A review. Can. J. Anim. Sci. 2006, 86, 135–157. [Google Scholar] [CrossRef]
- Boadi, D.; Wittenberg, K.; Kennedy, A. Validation of the sulphur hexafluoride (sf6) tracer gas technique for measurement of methane and carbon dioxide production by cattle. Can. J. Anim. Sci. 2002, 82, 125–131. [Google Scholar] [CrossRef]
- Deighton, M.H.; Williams, S.R.O.; Hannah, M.C.; Eckard, R.J.; Boland, T.M.; Wales, W.J.; Moate, P.J. A modified sulphur hexafluoride tracer technique enables accurate determination of enteric methane emissions from ruminants. Anim. Feed Sci. Technol. 2014, 197, 47–63. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.; Kennedy, P.; O’Neill, C.; Toovey, A.; Popovski, S.; Rea, S.; Pimm, C.; Klein, L. Reducing methane emissions in sheep by immunization against rumen methanogens. Vaccine 2004, 22, 3976–3985. [Google Scholar] [CrossRef] [PubMed]
- Grainger, C.; Clarke, T.; McGinn, S.; Auldist, M.; Beauchemin, K.; Hannah, M.; Waghorn, G.; Clark, H.; Eckard, R. Methane emissions from dairy cows measured using the sulfur hexafluoride (sf6) tracer and chamber techniques. J. Dairy Sci. 2007, 90, 2755–2766. [Google Scholar] [CrossRef]
- Lassey, K.; Pinares-Patiño, C.; Martin, R.; Molano, G.; McMillan, A. Enteric methane emission rates determined by the sf6 tracer technique: Temporal patterns and averaging periods. Anim. Feed Sci. Technol. 2011, 166, 183–191. [Google Scholar] [CrossRef]
- Pinares-Patiño, C.; Clark, H. Reliability of the sulfur hexafluoride tracer technique for methane emission measurement from individual animals: An overview. Aust. J. Exp. Agric. 2008, 48, 223–229. [Google Scholar] [CrossRef]
- Pinares-Patiño, C.; Lassey, K.; Martin, R.; Molano, G.; Fernandez, M.; MacLean, S.; Sandoval, E.; Luo, D.; Clark, H. Assessment of the sulphur hexafluoride (sf6) tracer technique using respiration chambers for estimation of methane emissions from sheep. Anim. Feed Sci. Technol. 2011, 166, 201–209. [Google Scholar] [CrossRef]
- Van Well, B.; Murray, S.; Hodgkinson, J.; Pride, R.; Strzoda, R.; Gibson, G.; Padgett, M. An open-path, hand-held laser system for the detection of methane gas. J. Opt. A Pure Appl. Opt. 2005, 7, S420. [Google Scholar] [CrossRef] [Green Version]
- Detto, M.; Verfaillie, J.; Anderson, F.; Xu, L.; Baldocchi, D. Comparing laser-based open-and closed-path gas analyzers to measure methane fluxes using the eddy covariance method. Agric. For. Meteorol. 2011, 151, 1312–1324. [Google Scholar] [CrossRef]
- Laubach, J.; Kelliher, F.M. Methane emissions from dairy cows: Comparing open-path laser measurements to profile-based techniques. Agric. For. Meteorol. 2005, 135, 340–345. [Google Scholar] [CrossRef]
- Desjardins, R.; Denmead, O.; Harper, L.; McBain, M.; Massé, D.; Kaharabata, S. Evaluation of a micrometeorological mass balance method employing an open-path laser for measuring methane emissions. Atmos. Environ. 2004, 38, 6855–6866. [Google Scholar] [CrossRef]
- Gao, Z.; Desjardins, R.L.; Flesch, T.K. Assessment of the uncertainty of using an inverse-dispersion technique to measure methane emissions from animals in a barn and in a small pen. Atmos. Environ. 2010, 44, 3128–3134. [Google Scholar] [CrossRef]
- McGinn, S.; Turner, D.; Tomkins, N.; Charmley, E.; Bishop-Hurley, G.; Chen, D. Methane emissions from grazing cattle using point-source dispersion. J. Environ. Qual. 2011, 40, 22–27. [Google Scholar] [CrossRef]
- Charmley, E.; McSweeney, C.; Eady, S. Strategies for measuring and reducing methane emissions from beef cattle in northern australia. In Proceedings of the Northern Beef Research Update Conference, Darwin, Austrilia, 2 August 2011; pp. 73–80. [Google Scholar]
- Hammond, K.; Waghorn, G.; Hegarty, R. The greenfeed system for measurement of enteric methane emission from cattle. Anim. Prod. Sci. 2016, 56, 181–189. [Google Scholar] [CrossRef]
- C-Lock. Greenfeed Large Animals. Available online: https://www.c-lockinc.com/researchers/products/greenfeed-pasture-system (accessed on 23 May 2020).
- Huhtanen, P.; Ramin, M.; Hristov, A.N. Enteric methane emission can be reliably measured by the greenfeed monitoring unit. Livest. Sci. 2019, 222, 31–40. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Giallongo, F.; Frederick, T.; Weeks, H.; Zimmerman, P.R.; Harper, M.T.; Hristova, R.A.; Zimmerman, R.S.; Branco, A.F. The use of an automated system (greenfeed) to monitor enteric methane and carbon dioxide emissions from ruminant animals. JoVE 2015, 103, e52904. [Google Scholar] [CrossRef] [Green Version]
- Waghorn, G.; Jonker, A.; Macdonald, K. Measuring methane from grazing dairy cows using greenfeed. Anim. Prod. Sci. 2016, 56, 252–257. [Google Scholar] [CrossRef]
- Hammond, K.; Humphries, D.; Crompton, L.; Green, C.; Reynolds, C.J.A.F.S. Methane emissions from cattle: Estimates from short-term measurements using a greenfeed system compared with measurements obtained using respiration chambers or sulphur hexafluoride tracer. Anim. Feed Sci. Technol. 2015, 203, 41–52. [Google Scholar] [CrossRef]
- Lind, V. Ruminants and Methane 2: 4 Methods and Techniques for Measuring Ghg Emissions from Ruminants; 2464-1170; Norsk Institutt for Bioøkonomi: Høgskoleveien, Norway, 2020; pp. 1–4. [Google Scholar]
- Goopy, J.; Robinson, D.L.; Woodgate, R.; Donaldson, A.; Oddy, V.; Vercoe, P.; Hegarty, R. Estimates of repeatability and heritability of methane production in sheep using portable accumulation chambers. Anim. Prod. Sci. 2016, 56, 116–122. [Google Scholar] [CrossRef]
- Robinson, D.L.; Goopy, J.P.; Hegarty, R.; Oddy, V. Comparison of repeated measurements of methane production in sheep over 5 years and a range of measurement protocols. J. Anim. Sci. 2015, 93, 4637–4650. [Google Scholar] [CrossRef] [PubMed]
- Goopy, J.P.; Woodgate, R.; Donaldson, A.; Robinson, D.L.; Hegarty, R. Validation of a short-term methane measurement using portable static chambers to estimate daily methane production in sheep. Anim. Feed Sci. Technol. 2011, 166, 219–226. [Google Scholar] [CrossRef]
- Robinson, D.; Dominik, S.; Donaldson, A.; Oddy, V. Repeatabilities, heritabilities and correlations of methane and feed intake of sheep in respiration and portable chambers. Anim. Prod. Sci. 2020, 60, 880–892. [Google Scholar] [CrossRef]
- Robinson, D.L.; Goopy, J.P.; Donaldson, A.; Woodgate, R.; Oddy, V.; Hegarty, R. Sire and liveweight affect feed intake and methane emissions of sheep confined in respiration chambers. Animal 2014, 8, 1935–1944. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.J. Measuring enteric methane emissions from individual ruminant animals in their natural environment. In Greenhouse Gas Emissions; Springer: Berlin/Heidelberg, Germany, 2019; pp. 111–126. [Google Scholar]
- Jayanegara, A. Reducing Methane Emissions from Livestock: Nutritional Approaches. In Proceedings of the Indonesian Students Scientific Meeting (ISSM), Manchester, UK, 25–26 August 2011; pp. 18–21. [Google Scholar]
- Beauchemin, K.A.; Kreuzer, M.; O’Mara, F.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27. [Google Scholar] [CrossRef]
- Sejian, V.; Lal, R.; Lakritz, J.; Ezeji, T. Measurement and prediction of enteric methane emission. Int. J. Biometeorol. 2011, 55, 1–16. [Google Scholar] [CrossRef]
- Pesta, A.C. Dietary strategies for mitigation of methane production by growing and finishing cattle. Ph.D. Thesis, University of Nebraska, Lincoln, NE, USA, 2015. [Google Scholar]
- Haque, M. Dietary manipulation: A sustainable way to mitigate methane emissions from ruminants. J. Anim. Sci. Technol. 2018, 60, 15. [Google Scholar] [CrossRef] [Green Version]
- Nampoothiri, V.M.; Mohini, M.; Malla, B.A.; Mondal, G.; Pandita, S. Growth performance, and enteric and manure greenhouse gas emissions from murrah calves fed diets with different forage to concentrate ratios. Anim. Nutr. 2018, 4, 215–221. [Google Scholar] [CrossRef]
- Jiao, H.P.; Dale, A.J.; Carson, A.F.; Murray, S.; Gordon, A.W.; Ferris, C.P. Effect of concentrate feed level on methane emissions from grazing dairy cows. J. Dairy Sci. 2014, 97, 7043–7053. [Google Scholar] [CrossRef] [PubMed]
- Lovett, D.; Lovell, S.; Stack, L.; Callan, J.; Finlay, M.; Conolly, J.; O’Mara, F. Effect of forage/concentrate ratio and dietary coconut oil level on methane output and performance of finishing beef heifers. Livest. Prod. Sci. 2003, 84, 135–146. [Google Scholar] [CrossRef]
- Sabia, E.; Kühl, S.; Flach, L.; Lambertz, C.; Gauly, M. Effect of feed concentrate intake on the environmental impact of dairy cows in an alpine mountain region including soil carbon sequestration and effect on biodiversity. Sustainability 2020, 12, 2128. [Google Scholar] [CrossRef] [Green Version]
- Van Wyngaard, J.D.V.; Meeske, R.; Erasmus, L.J. Effect of concentrate level on enteric methane emissions, production performance, and rumen fermentation of jersey cows grazing kikuyu-dominant pasture during summer. J. Dairy Sci. 2018, 101, 9954–9966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguerre, M.J.; Wattiaux, M.A.; Powell, J.; Broderick, G.A.; Arndt, C. Effect of forage-to-concentrate ratio in dairy cow diets on emission of methane, carbon dioxide, and ammonia, lactation performance, and manure excretion. J. Dairy Sci. 2011, 94, 3081–3093. [Google Scholar] [CrossRef]
- Muñoz, C.; Hube, S.; Morales, J.M.; Yan, T.; Ungerfeld, E.M. Effects of concentrate supplementation on enteric methane emissions and milk production of grazing dairy cows. Livest. Sci. 2015, 175, 37–46. [Google Scholar] [CrossRef]
- Dohme, F.; Machmüller, A.; Wasserfallen, A.; Kreuzer, M. Comparative efficiency of various fats rich in medium-chain fatty acids to suppress ruminal methanogenesis as measured with rusitec. Can. J. Anim. Sci. 2000, 80, 473–484. [Google Scholar] [CrossRef]
- Grainger, C.; Beauchemin, K. Can enteric methane emissions from ruminants be lowered without lowering their production? Anim. Feed Sci. Technol. 2011, 166, 308–320. [Google Scholar] [CrossRef]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef] [Green Version]
- Machmüller, A.; Kreuzer, M. Methane suppression by coconut oil and associated effects on nutrient and energy balance in sheep. Can. J. Anim. Sci. 1999, 79, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Pinares-Patiño, C.S.; Franco, F.E.; Molano, G.; Kjestrup, H.; Sandoval, E.; MacLean, S.; Battistotti, M.; Koolaard, J.; Laubach, J. Feed intake and methane emissions from cattle grazing pasture sprayed with canola oil. Livest. Sci. 2016, 184, 7–12. [Google Scholar] [CrossRef]
- Mao, H.-L.; Wang, J.-K.; Zhou, Y.-Y.; Liu, J.-X. Effects of addition of tea saponins and soybean oil on methane production, fermentation and microbial population in the rumen of growing lambs. Livest. Sci. 2010, 129, 56–62. [Google Scholar] [CrossRef]
- Chuntrakort, P.; Otsuka, M.; Hayashi, K.; Takenaka, A.; Udchachon, S.; Sommart, K. The effect of dietary coconut kernels, whole cottonseeds and sunflower seeds on the intake, digestibility and enteric methane emissions of zebu beef cattle fed rice straw based diets. Livest. Sci. 2014, 161, 80–89. [Google Scholar] [CrossRef]
- Machmüller, A.; Ossowski, D.; Kreuzer, M. Comparative evaluation of the effects of coconut oil, oilseeds and crystalline fat on methane release, digestion and energy balance in lambs. Anim. Feed Sci. Technol. 2000, 85, 41–60. [Google Scholar] [CrossRef]
- Cosgrove, G.; Waghorn, G.; Anderson, C.; Peters, J.; Smith, A.; Molano, G.; Deighton, M. The effect of oils fed to sheep on methane production and digestion of ryegrass pasture. Aust. J. Exp. Agric. 2008, 48, 189–192. [Google Scholar] [CrossRef]
- Hersom, M.; Thrift, T. Application of Ionophores in Cattle Diets; University of Florida: Gainesville, FL, USA, 2013; Volume 14. [Google Scholar]
- Valli, C. Mitigating enteric methane emission from livestock through farmer-friendly practices. In Global Climate Change and Environmental Policy; Springer: Berlin/Heidelberg, Germany, 2020; pp. 257–273. [Google Scholar]
- Perna Junior, F.; Vásquez, D.C.Z.; Gardinal, R.; Meyer, P.M.; Berndt, A.; Friguetto, R.T.S.; Demarchi, J.A.; Rodrigues, P.H.M. Short-term use of monensin and tannins as feed additives on digestibility and methanogenesis in cattle. Rev. Bras. Zootec. 2020, 49. [Google Scholar] [CrossRef]
- Guan, H.; Wittenberg, K.; Ominski, K.; Krause, D. Efficacy of ionophores in cattle diets for mitigation of enteric methane. J. Anim. Sci. Technol. 2006, 84, 1896–1906. [Google Scholar]
- Li, Z.; Ren, H.; Liu, S.; Cai, C.; Han, J.; Li, F.; Yao, J. Dynamics of methanogenesis, ruminal fermentation, and alfalfa degradation during adaptation to monensin supplementation in goats. J. Dairy Sci. 2018, 101, 1048–1059. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Mohini, M.; Malla, B.A.; Mondal, G.; Pandita, S. Effects of monensin feeding on performance, nutrient utilisation and enteric methane production in growing buffalo heifers. Trop. Anim. Health Prod. 2019, 51, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Kunkle, W.; Johns, J.; Poore, M.; Herd, D. Designing supplementation programs for beef cattle fed forage-based diets. J. Anim. Sci. 2000, 77, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lascano, C.E.; Cárdenas, E. Alternatives for methane emission mitigation in livestock systems. Rev. Bras. Zootecn 2010, 39, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Matsui, H.; Tajima, K.; Itabashi, H. Diversity of prokaryotes in the rumen of steers fed a diet supplemented with or without bromochloromethane, an anti-methanogenic compound. Jpn. Agric. Res. Q. JARQ 2020, 54, 179–183. [Google Scholar] [CrossRef]
- Lalu, K.; Bhar, R.; Das, A.; Mandal, A. Effect of bromochloromethane supplementation and dietary energy restriction on methane production and efficiency of energy utilization in rams. Indian J. Anim. Nutr. 2009, 26, 97–102. [Google Scholar]
- Romero-Perez, A.; Okine, E.K.; McGinn, S.M.; Guan, L.L.; Oba, M.; Duval, S.M.; Kindermann, M.; Beauchemin, K.A. Sustained reduction in methane production from long-term addition of 3-nitrooxypropanol to a beef cattle diet. J. Anim. Sci. 2015, 93, 1780–1791. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.; de Matos, L.; Harper, M.; Giallongo, F.; Oh, J.; Gruen, D.; Ono, S.; Kindermann, M.; Duval, S.; Hristov, A.N. Effect of 3-nitrooxypropanol on methane and hydrogen emissions, methane isotopic signature, and ruminal fermentation in dairy cows. J. Dairy Sci. 2016, 99, 5335–5344. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Fernández, G.; Abecia, L.; Arco, A.; Cantalapiedra-Hijar, G.; Martín-García, A.I.; Molina-Alcaide, E.; Kindermann, M.; Duval, S.; Yáñez-Ruiz, D.R. Effects of ethyl-3-nitrooxy propionate and 3-nitrooxypropanol on ruminal fermentation, microbial abundance, and methane emissions in sheep. J. Dairy Sci. 2014, 97, 3790–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, L.; Magnusson, M.; Paul, N.A.; Kinley, R.; de Nys, R.; Tomkins, N. Identification of bioactives from the red seaweed asparagopsis taxiformis that promote antimethanogenic activity in vitro. J. Appl. Phycol. 2016, 28, 3117–3126. [Google Scholar] [CrossRef]
- Paul, N.A.; de Nys, R.; Steinberg, P. Chemical defence against bacteria in the red alga asparagopsis armata: Linking structure with function. Mar. Ecol. Prog. Ser. 2006, 306, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Norman, H.C.; Kinley, R.D.; Laurence, M.; Wilmot, M.; Bender, H.; de Nys, R.; Tomkins, N. Asparagopsis taxiformis decreases enteric methane production from sheep. Anim. Prod. Sci. 2018, 58, 681–688. [Google Scholar] [CrossRef]
- Kinley, R.D.; de Nys, R.; Vucko, M.J.; Machado, L.; Tomkins, N. The red macroalgae asparagopsis taxiformis is a potent natural antimethanogenic that reduces methane production during in vitro fermentation with rumen fluid. Anim. Prod. Sci. 2016, 56, 282–289. [Google Scholar] [CrossRef]
- Roque, B.M.; Salwen, J.K.; Kinley, R.; Kebreab, E. Inclusion of asparagopsis armata in lactating dairy cows’ diet reduces enteric methane emission by over 50 percent. J. Clean. Prod. 2019, 234, 132–138. [Google Scholar] [CrossRef]
- Martínez, G.; Abecia, L.; Martín-García, A.; Ramos-Morales, E.; Molina-Alcaide, E.; Yáñez-Ruiz, D. Effect of antimethanogenic garlic-derived compounds on amylolytic and xylanolytic activities in the rumen. Options Méditerranéennes. Série A Séminaires Méditerranéens 2013, 107, 277–282. [Google Scholar]
- Klevenhusen, F.; Duval, S.; Zeitz, J.O.; Kreuzer, M.; Soliva, C.R. Diallyl disulphide and lovastatin: Effects on energy and protein utilisation in, as well as methane emission from, sheep. Arch. Anim. Nutr. 2011, 65, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Bidarkar, V.K.; Swain, P.S.; Ray, S.; Dominic, G. Probiotics: Potential alternative to antibiotics in ruminant feeding. Trends Vet. Anim. Sci. 2014, 1, 1–4. [Google Scholar]
- Hassan, A.; Gado, H.; Anele, U.Y.; Berasain, M.A.; Salem, A.Z. Influence of dietary probiotic inclusion on growth performance, nutrient utilization, ruminal fermentation activities and methane production in growing lambs. Anim. Biotechnol. 2019, 31, 1–8. [Google Scholar] [CrossRef]
- Arowolo, M.A.; He, J. Use of probiotics and botanical extracts to improve ruminant production in the tropics: A review. Anim. Nutr. 2018, 4, 241–249. [Google Scholar] [CrossRef]
- Deng, K.D.; Xiao, Y.; Ma, T.; Tu, Y.; Diao, Q.Y.; Chen, Y.H.; Jiang, J.J. Ruminal fermentation, nutrient metabolism, and methane emissions of sheep in response to dietary supplementation with bacillus licheniformis. Anim. Feed Sci. Technol. 2018, 241, 38–44. [Google Scholar] [CrossRef]
- Latham, E.A.; Pinchak, W.E.; Trachsel, J.; Allen, H.K.; Callaway, T.R.; Nisbet, D.J.; Anderson, R.C. Paenibacillus 79r4, a potential rumen probiotic to enhance nitrite detoxification and methane mitigation in nitrate-treated ruminants. Sci. Total Environ. 2019, 671, 324–328. [Google Scholar] [CrossRef]
- Suryani, H.; Zain, M.; Ningrat, R.; Jamarun, N. Effect of dietary supplementation based on an ammoniated palm frond with direct fed microbials and virgin coconut oil on the growth performance and methane production of bali cattle. Pak. J. Nutr. 2017, 16, 599–604. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Harstad, O.M.; McAllister, T.; Dörsch, P.; Holo, H. Propionic acid bacteria enhance ruminal feed degradation and reduce methane production in vitro. Acta Agric. Scand. Sect. A Anim. Sci. 2020, 69, 1–7. [Google Scholar] [CrossRef] [Green Version]
- De Raphelis-Soissan, V.; Li, L.; Godwin, I.; Barnett, M.; Perdok, H.; Hegarty, R. Use of nitrate and propionibacterium acidipropionici to reduce methane emissions and increase wool growth of merino sheep. Anim. Prod. Sci. 2014, 54, 1860–1866. [Google Scholar] [CrossRef]
- Cobellis, G.; Trabalza-Marinucci, M.; Yu, Z. Critical evaluation of essential oils as rumen modifiers in ruminant nutrition: A review. Sci. Total Environ. 2016, 545, 556–568. [Google Scholar] [CrossRef] [PubMed]
- Benchaar, C.; Chaves, A.; Fraser, G.; Beauchemin, K.; McAllister, T. Effects of essential oils and their components on in vitro rumen microbial fermentation. Can. J. Anim. Sci. 2007, 87, 413–419. [Google Scholar] [CrossRef]
- Benchaar, C.; Greathead, H. Essential oils and opportunities to mitigate enteric methane emissions from ruminants. Anim. Feed Sci. Technol. 2011, 166–167, 338–355. [Google Scholar] [CrossRef]
- Wu, P.; Liu, Z.B.; He, W.F.; Yu, S.B.; Gao, G.; Wang, J.K. Intermittent feeding of citrus essential oils as a potential strategy to decrease methane production by reducing microbial adaptation. J. Clean. Prod. 2018, 194, 704–713. [Google Scholar] [CrossRef]
- Hart, K.J.; Jones, H.G.; Waddams, K.E.; Worgan, H.J.; Zweifel, B.; Newbold, C.J. An essential oil blend decreases methane emissions and increases milk yield in dairy cows. Open J. Anim. Sci. 2019, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Laabouri, F.; Guerouali, A.; Alali, S.; Remmal, A.; Ajbilou, M. Effect of a natural food additive rich in thyme essential oil on methane emissions in dairy cows. Rev. Maroc. Des Sci. Agron. Et Vétérinaires 2017, 5, 287–292. [Google Scholar]
- Soltan, Y.A.; Natel, A.S.; Araujo, R.; Morsy, A.S.; Abdalla, A.L. Progressive adaptation of sheep to a microencapsulated blend of essential oils: Ruminal fermentation, methane emission, nutrient digestibility, and microbial protein synthesis. Anim. Feed Sci. Technol. 2018, 237, 8–18. [Google Scholar] [CrossRef]
- Sallama, S.; Nassera, M.; Araujoc, R.; Abdallab, A. Methane production by sheep consuming diets with different levels of eucalyptus essential oil. In Proceedings of the FAO/IAEA International Symposium on Sustainable Improvement of Animal Production and Health, Vienna, Austria, 8–11 June 2009; Available online: https://inis.iaea.org/search/search.aspx?orig_q=RN:41037857 (accessed on 18 June 2020).
- Sahoo, A.; Jena, B. Organic acids as rumen modifiers. Int. J. Sci. Res. 2014, 3, 2262–2266. [Google Scholar]
- Newbold, C.J.; Rode, L.M. Dietary additives to control methanogenesis in the rumen. Int. Congr. Ser. 2006, 1293, 138–147. [Google Scholar] [CrossRef]
- Bayaru, E.; Kanda, S.; Kamada, T.; Itabashi, H.; Andoh, S.; Nishida, T.; Ishida, M.; Itoh, T.; Nagara, K.; Isobe, Y. Effect of fumaric acid on methane production, rumen fermentation and digestibility of cattle fed roughage alone. Anim. Sci. J. 2001, 72, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Wallace, R.J.; Wood, T.A.; Rowe, A.; Price, J.; Yanez, D.R.; Williams, S.P.; Newbold, C.J. Encapsulated fumaric acid as a means of decreasing ruminal methane emissions. Int. Congr. Ser. 2006, 1293, 148–151. [Google Scholar] [CrossRef]
- Ma, T.; Chen, D.; Tu, Y.; Zhang, N.; Si, B.; Deng, K.; Diao, Q. Effect of supplementation of allicin on methanogenesis and ruminal microbial flora in dorper crossbred ewes. J. Anim. Sci. Biotechnol. 2016, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Foley, P.A.; Kenny, D.A.; Callan, J.J.; Boland, T.M.; O’Mara, F.P. Effect of dl-malic acid supplementation on feed intake, methane emission, and rumen fermentation in beef cattle. J. Anim. Sci. 2009, 87, 1048–1057. [Google Scholar] [CrossRef]
- Jalč, D.; Čerešňáková, Z. Effect of plant oils and aspartate on rumen fermentation in vitro. J. Anim. Physiol. Anim. Nutr. 2001, 85, 378–384. [Google Scholar] [CrossRef]
- McAllister, T.; Hristov, A.; Beauchemin, K.; Rode, L.; Cheng, K. Enzymes in ruminant diets. In Enzymes in Farm Animal Nutrition; Bedford, M.R., Partridge, G.G., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 273–298. [Google Scholar]
- Zhou, M.; Chung, Y.H.; Beauchemin, K.; Holtshausen, L.; Oba, M.; McAllister, T.; Guan, L. Relationship between rumen methanogens and methane production in dairy cows fed diets supplemented with a feed enzyme additive. J. Appl. Microbiol. 2011, 111, 1148–1158. [Google Scholar] [CrossRef] [PubMed]
- Arriola, K.G.; Kim, S.C.; Staples, C.R.; Adesogan, A.T. Effect of fibrolytic enzyme application to low- and high-concentrate diets on the performance of lactating dairy cattle. J. Dairy Sci. 2011, 94, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Peng, Y.; Wang, J.; Liu, J. Effects of exogenous fibrolytic enzyme on in vitro ruminal fiber digestion and methane production of corn stover and corn stover based mixed diets. Life Sci. J. 2015, 12, 1–9. [Google Scholar] [CrossRef]
- McGinn, S.M.; Beauchemin, K.A.; Coates, T.; Colombatto, D. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. J. Anim. Sci. 2004, 82, 3346–3356. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Harper, M.; Melgar, A.; Compart, D.M.P.; Hristov, A.N. Effects of saccharomyces cerevisiae-based direct-fed microbial and exogenous enzyme products on enteric methane emission and productivity in lactating dairy cows. J. Dairy Sci. 2019, 102, 6065–6075. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Jiao, J.; Tang, S.; He, Z.; Zhou, C.; Han, X.; Wang, M.; Kang, J.; Odongo, N.; Tan, Z. Effects of dietary cellulase and xylanase addition on digestion, rumen fermentation and methane emission in growing goats. Arch. Anim. Nutr. 2015, 69, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.; Siddhuraju, P.; Becker, K. Preface. In Plant Secondary Metabolites; Humana Press: Totowa, NJ, USA, 2007; pp. i–xi. [Google Scholar]
- Rochfort, S.; Parker, A.J.; Dunshea, F.R. Plant bioactives for ruminant health and productivity. Phytochemistry 2008, 69, 299–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodas, R.; Prieto, N.; García-González, R.; Andrés, S.; Giráldez, F.J.; López, S. Manipulation of rumen fermentation and methane production with plant secondary metabolites. Anim. Feed Sci. Technol. 2012, 176, 78–93. [Google Scholar] [CrossRef]
- Malik, P.; Uyeno, Y.; Kolte, A.; Kumar, R.; Trivedi, S.; Bhatta, R. Screening of phyto-sources from foothill of himalayan mountain for livestock methane reduction. Appl. Sci. 2019, 1, 232. [Google Scholar] [CrossRef]
- Baruah, L.; Malik, P.K.; Kolte, A.P.; Goyal, P.; Dhali, A.; Bhatta, R. Rumen methane amelioration in sheep using two selected tanniferous phyto-leaves. Carbon Manag. 2019, 10, 299–308. [Google Scholar] [CrossRef]
- Moreira, G.D.; Lima, P.d.M.T.; Borges, B.O.; Primavesi, O.; Longo, C.; McManus, C.; Abdalla, A.; Louvandini, H. Tropical tanniniferous legumes used as an option to mitigate sheep enteric methane emission. Trop. Anim. Health Prod. 2013, 45, 879–882. [Google Scholar] [CrossRef]
- Śliwiński, B.; Kreuzer, M.; Wettstein, H.-R.; Machmüller, A. Rumen fermentation and nitrogen balance of lambs fed diets containing plant extracts rich in tannins and saponins, and associated emissions of nitrogen and methane. Arch. Anim. Nutr. 2002, 56, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Carulla, J.; Kreuzer, M.; Machmüller, A.; Hess, H. Supplementation of acacia mearnsii tannins decreases methanogenesis and urinary nitrogen in forage-fed sheep. Aust. J. Agric. Res. 2005, 56, 961–970. [Google Scholar] [CrossRef]
- Malik, P.K.; Kolte, A.P.; Bakshi, B.; Baruah, L.; Dhali, A.; Bhatta, R. Effect of tamarind seed husk supplementation on ruminal methanogenesis, methanogen diversity and fermentation characteristics. Carbon Manag. 2017, 8, 319–329. [Google Scholar] [CrossRef]
- Wann, C.; Wanapat, M.; Mapato, C.; Ampapon, T.; Huang, B.-z. Effect of bamboo grass (tiliacora triandra, diels) pellet supplementation on rumen fermentation characteristics and methane production in thai native beef cattle. Asian Australas. J. Anim. Sci. 2019, 32, 1153. [Google Scholar] [CrossRef]
- Liu, H.; Vaddella, V.; Zhou, D. Effects of chestnut tannins and coconut oil on growth performance, methane emission, ruminal fermentation, and microbial populations in sheep. J. Dairy Sci. 2011, 94, 6069–6077. [Google Scholar] [CrossRef] [PubMed]
- Albores-Moreno, S.; Alayón-Gamboa, J.; Ayala-Burgos, A.; Solorio-Sánchez, F.; Aguilar-Pérez, C.; Olivera-Castillo, L.; Ku-Vera, J. Effects of feeding ground pods of enterolobium cyclocarpum jacq. Griseb on dry matter intake, rumen fermentation, and enteric methane production by pelibuey sheep fed tropical grass. Trop. Anim. Health Prod. 2017, 49, 857–866. [Google Scholar] [CrossRef]
- Váradyová, Z.; Zeleňák, I.; Siroka, P. In vitro study of the rumen and hindgut fermentation of fibrous materials (meadow hay, beech sawdust, wheat straw) in sheep. Anim. Feed Sci. Technol. 2000, 83, 127–138. [Google Scholar] [CrossRef]
- Lovett, D.K.; Stack, L.J.; Lovell, S.; Callan, J.; Flynn, B.; Hawkins, M.; O’Mara, F.P. Manipulating enteric methane emissions and animal performance of late-lactation dairy cows through concentrate supplementation at pasture. J. Dairy Sci. 2005, 88, 2836–2842. [Google Scholar] [CrossRef]
- Dall-Orsoletta, A.C.; Oziemblowski, M.M.; Berndt, A.; Ribeiro-Filho, H.M.N. Enteric methane emission from grazing dairy cows receiving corn silage or ground corn supplementation. Anim. Feed Sci. Technol. 2019, 253, 65–73. [Google Scholar] [CrossRef]
- Ferris, C.P.; Jiao, H.; Murray, S.; Gordon, A.; Laidlaw, S. Effect of dairy cow genotype and concentrate feed level on cow performance and enteric methane emissions during grazing. Agric. Food Sci. 2020, 29, 130–138. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, Y.; Aubry, A.; Arnott, G.; Hou, F.; Yan, T. Effects of concentrate input on nutrient utilization and methane emissions of two breeds of ewe lambs fed fresh ryegrass. Transl. Anim. Sci. 2019, 3, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Hynes, D.N.; Stergiadis, S.; Gordon, A.; Yan, T. Effects of concentrate crude protein content on nutrient digestibility, energy utilization, and methane emissions in lactating dairy cows fed fresh-cut perennial grass. J. Dairy Sci. 2016, 99, 8858–8866. [Google Scholar] [CrossRef] [Green Version]
- Talukdar, P.; Kundu, S.S.; Mondal, G. Quantification of methane emissions from murrah buffaloes fed different energy diets during various temperature humidity index periods in a tropical environment. Anim. Prod. Sci. 2019, 59, 169–176. [Google Scholar] [CrossRef]
- Puchala, R.; LeShure, S.; Gipson, T.A.; Tesfai, K.; Flythe, M.D.; Goetsch, A.L. Effects of different levels of lespedeza and supplementation with monensin, coconut oil, or soybean oil on ruminal methane emission by mature boer goat wethers after different lengths of feeding. J. Appl. Anim. Res. 2018, 46, 1127–1136. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.T.K.; Van Thu, N. Dietary supplementation of coconut oil markedly suppressed enteric methane production without compromising growth performance in bach thao goats. In Strengthening Development of Dairy Goat Production Adapting to Climate Change, Proceedings of the 4th Asian-Australasian Dairy Goat Conference, Tra Vinh, Viet Nam, 17–19 October 2018; Van Thu, N., Liang, J.B., Eds.; Abstract Number 54; Available online: https://www.researchgate.net/profile/Aliah-Mohsin/publication/333773806_4thAADC_Proceedings_TVU_2018/links/5d03284e92851c874c65066b/4thAADC-Proceedings-TVU-2018.pdf#page=404 (accessed on 22 March 2020).
- Lima, P.R.; Apdini, T.; Freire, A.S.; Santana, A.S.; Moura, L.M.L.; Nascimento, J.C.S.; Rodrigues, R.T.S.; Dijkstra, J.; Garcez Neto, A.F.; Queiroz, M.A.Á.; et al. Dietary supplementation with tannin and soybean oil on intake, digestibility, feeding behavior, ruminal protozoa and methane emission in sheep. Anim. Feed Sci. Technol. 2019, 249, 10–17. [Google Scholar] [CrossRef]
- Zhang, X.M.; Medrano, R.F.; Wang, M.; Beauchemin, K.A.; Ma, Z.Y.; Wang, R.; Wen, J.N.; Lukuyu, B.A.; Tan, Z.L.; He, J.H. Corn oil supplementation enhances hydrogen use for biohydrogenation, inhibits methanogenesis, and alters fermentation pathways and the microbial community in the rumen of goats. J. Anim. Sci. 2019, 97, 4999–5008. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Coppa, M.; Fougère, H.; Bougouin, A.; Baumont, R.; Eugène, M.; Bernard, L. Diets supplemented with corn oil and wheat starch, marine algae, or hydrogenated palm oil modulate methane emissions similarly in dairy goats and cows, but not feeding behavior. Anim. Feed Sci. Technol. 2021, 272, 114783. [Google Scholar] [CrossRef]
- Pickett, A.T. Effects Of Lasalocid and Energy Supplementation on Forage Intake, Energy Metabolism, and Performance of Cattle Grazing Wheat Pasture; University of Arkansas: Fayetteville, AR, USA, 2020. [Google Scholar]
- Mwenya, B.; Sar, C.; Santoso, B.; Kobayashi, T.; Morikawa, R.; Takaura, K.; Umetsu, K.; Kogawa, S.; Kimura, K.; Mizukoshi, H.; et al. Comparing the effects of β1-4 galacto-oligosaccharides and l-cysteine to monensin on energy and nitrogen utilization in steers fed a very high concentrate diet. Anim. Feed Sci. Technol. 2005, 118, 19–30. [Google Scholar] [CrossRef]
- Mwenya, B.; Santoso, B.; Sar, C.; Gamo, Y.; Kobayashi, T.; Arai, I.; Takahashi, J. Effects of including β1–4 galacto-oligosaccharides, lactic acid bacteria or yeast culture on methanogenesis as well as energy and nitrogen metabolism in sheep. Anim. Feed Sci. Technol. 2004, 115, 313–326. [Google Scholar] [CrossRef]
- Fernández, C.; Pérez-Baena, I.; Marti, J.V.; Palomares, J.L.; Jorro-Ripoll, J.; Segarra, J.V. Use of orange leaves as a replacement for alfalfa in energy and nitrogen partitioning, methane emissions and milk performance of murciano-granadina goats. Anim. Feed Sci. Technol. 2019, 247, 103–111. [Google Scholar] [CrossRef]
- Wood, T.A.; Wallace, R.J.; Rowe, A.; Price, J.; Yáñez-Ruiz, D.R.; Murray, P.; Newbold, C.J. Encapsulated fumaric acid as a feed ingredient to decrease ruminal methane emissions. Anim. Feed Sci. Technol. 2009, 152, 62–71. [Google Scholar] [CrossRef]
- Wang, L.; Xue, B. Effects of cellulase supplementation on nutrient digestibility, energy utilization and methane emission by boer crossbred goats. Asian Australas. J. Anim. Sci. 2016, 29, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Soltan, Y.A.; Morsy, A.S.; Sallam, S.M.; Lucas, R.C.; Louvandini, H.; Kreuzer, M.; Abdalla, A.L. Contribution of condensed tannins and mimosine to the methane mitigation caused by feeding leucaena leucocephala. Arch. Anim. Nutr. 2013, 67, 169–184. [Google Scholar] [CrossRef]
- Moate, P.J.; Jacobs, J.L.; Hixson, J.L.; Deighton, M.H.; Hannah, M.C.; Morris, G.L.; Ribaux, B.E.; Wales, W.J.; Williams, S.R.O. Effects of feeding either red or white grape marc on milk production and methane emissions from early-lactation dairy cows. Animals 2020, 10, 976. [Google Scholar] [CrossRef]
- Ramírez-Restrepo, C.A.; Barry, T.N.; Marriner, A.; López-Villalobos, N.; McWilliam, E.L.; Lassey, K.R.; Clark, H. Effects of grazing willow fodder blocks upon methane production and blood composition in young sheep. Anim. Feed Sci. Technol. 2010, 155, 33–43. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prathap, P.; Chauhan, S.S.; Leury, B.J.; Cottrell, J.J.; Dunshea, F.R. Towards Sustainable Livestock Production: Estimation of Methane Emissions and Dietary Interventions for Mitigation. Sustainability 2021, 13, 6081. https://doi.org/10.3390/su13116081
Prathap P, Chauhan SS, Leury BJ, Cottrell JJ, Dunshea FR. Towards Sustainable Livestock Production: Estimation of Methane Emissions and Dietary Interventions for Mitigation. Sustainability. 2021; 13(11):6081. https://doi.org/10.3390/su13116081
Chicago/Turabian StylePrathap, Pragna, Surinder Singh Chauhan, Brian Joseph Leury, Jeremy James Cottrell, and Frank Rowland Dunshea. 2021. "Towards Sustainable Livestock Production: Estimation of Methane Emissions and Dietary Interventions for Mitigation" Sustainability 13, no. 11: 6081. https://doi.org/10.3390/su13116081