
Foliar Application of Gibberellin Alleviates Adverse Impacts of Drought Stress and Improves Growth, Physiological and Biochemical Attributes of Canola (Brassica napus L.)
 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
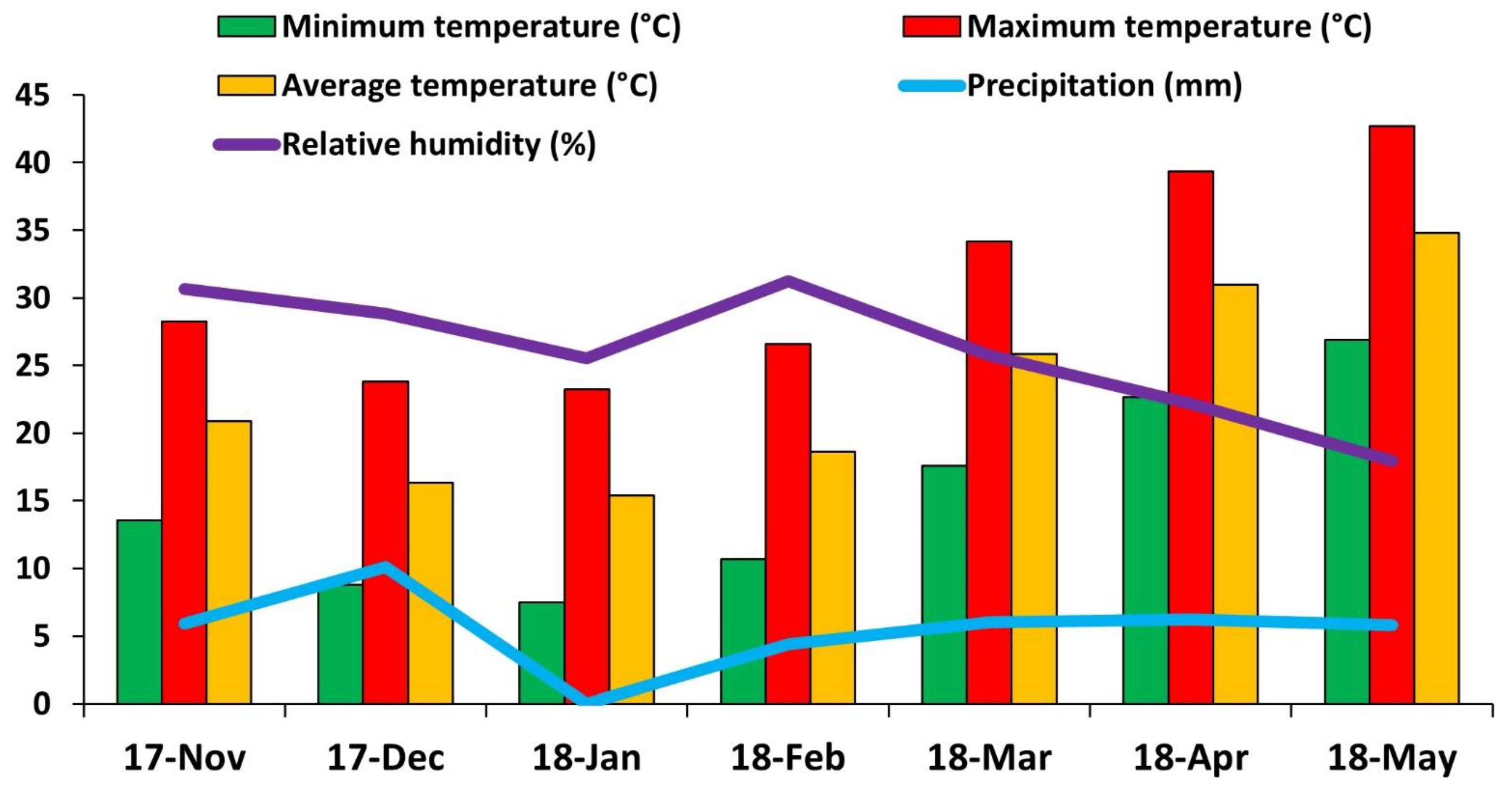
2.1. Soil Analysis
2.2. Pot Experiment
2.3. Data Collection
2.4. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Global Forest Resources Assessment, 2010—Main Report; FAO Forestry Paper 163; FAO: Rome, Italy, 2010. [Google Scholar]
- Finance Division Economic Adviser’s Wing, Government of Pakistan. Pakistan Economic Survey, 2017–18; Finance Division Economic Adviser’s Wing, Government of Pakistan: Islamabad, Pakistan, 2018. [Google Scholar]
- Mishra, S.; Manchanda, S.C. Cooking oils for heart health. J. Prev. Cardiol. 2012, 1, 123–131. [Google Scholar]
- Shahzadi, H.; Sheikh, M.A.; Hameed, A.; Jamil, A.; Rehman, S.U. Comparative antioxidant potential and bioactivity of maize (Zea mays) ear tissues from different genotypes. Int. J. Agric. Biol. 2015, 17, 539–546. [Google Scholar] [CrossRef]
- Harvey, B.; Downey, R. The inheritance of erucic acid content in rapeseed (Brassica napus L.). Can. J. Plant Sci. 1964, 44, 104–111. [Google Scholar] [CrossRef]
- Dupont, J.; White, P.; Johnston, K.; Heggtveit, H.; McDonald, B.; Grundy, S.M.; Bonanome, A. Food safety and health effects of canola oil. J. Am. Coll. Nutr. 1989, 8, 360–375. [Google Scholar] [CrossRef]
- Lin, L.; Allemekinders, H.; Dansby, A.; Campbell, L.; Durance-Tod, S.; Berger, A.; Jones, P.J. Evidence of health benefits of canola oil. Nutr. Rev. 2013, 71, 370–385. [Google Scholar] [CrossRef] [Green Version]
- Moghadam, H.R.T.; Zahedi, H.; Ghooshchi, F. Oil quality of canola cultivars in response to water stress and super absorbent polymer application. Pesqui. Agropecu. Trop. 2011, 41, 579–586. [Google Scholar]
- Trenberth, K.E.; Dai, A.; van der Schrier, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffeld, J. Global warming, and changes in drought. Nat. Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Ullah, N.; Ditta, A.; Imtiaz, M.; Li, X.; Jan, A.U.; Mehmood, S.; Rizwan, M.S.; Rizwan, M. Appraisal for organic amendments and plant growth-promoting rhizobacteria to enhance crop productivity under drought stress: A review. J. Agron. Crop Sci. 2021, 207, 783–802. [Google Scholar] [CrossRef]
- Ahmad, R.; Hadi, F.; Jan, A.U.; Ditta, A. Straw incorporation enhances drought stress tolerance but at the same time increases bioaccumulation of heavy metals under contaminated soil in Oryza sativa L. Sustainability 2022, 14, 10578. [Google Scholar] [CrossRef]
- Jan, A.U.; Hadi, F.; Ditta, A.; Suleman, M.; Ullah, M. Zinc-induced anti-oxidative defense and osmotic adjustments to enhance drought stress tolerance in sunflower (Helianthus annuus L.). Environ. Exp. Bot. 2022, 193, 104682. [Google Scholar] [CrossRef]
- Nath, R.; Nath, D.; Li, Q.; Chen, W.; Cui, X.F. Impact of drought on agriculture in the Indo-Gangetic Plain, India. Adv. Atmos. Sci. 2016, 34, 335–346. [Google Scholar] [CrossRef]
- Ahmad, H.T.; Hussain, A.; Aimen, A.; Jamshaid, M.U.; Ditta, A.; Asghar, H.N.; Zahir, Z.A. Improving resilience against drought stress among crop plants through inoculation of plant growth-promoting rhizobacteria. In Harsh Environment and Plant Resilience: Molecular and Functional Aspects; Husen, A., Jawaid, M., Eds.; Springer: Cham, Switzerland, 2021; pp. 387–408. [Google Scholar]
- Urban, M.O.; Vašek, J.; Klíma, M.; Krtková, J.; Kosová, K.; Prášil, I.T.; Vítámvás, P. Proteomic and physiological approach reveals drought-induced changes in rapeseeds: Water-saver and water-spender strategy. J. Proteom. 2017, 152, 188–205. [Google Scholar] [CrossRef]
- Pillai, B.V.S.; Kagale, S.; Chellamma, S. Enhancing productivity and performance of oil seed crops under environmental stresses. In Crop Stress and Its Management: Perspectives and Strategies; Venkateswarlu, B., Shanker, A.K., Shanker, C., Maheswari, M., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 139–161. [Google Scholar]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Ahmadi, M.; Bahrani, M.J. Yield and yield components of rapeseed as influenced by water stress at different growth stages and nitrogen levels. Am. Eurasian J. Agric. Environ. Sci. 2009, 5, 755–761. [Google Scholar]
- Tinker, N.A. Why quantitative geneticists should care about bioinformatics. In Quantitative Genetics, Genomics, and Plant Breeding; CABI: Wallingford, UK, 2002; pp. 33–44. [Google Scholar]
- Parveen, A.; Ashraf, M.A.; Hussain, I.; Perveen, S.; Rasheed, R.; Mahmood, Q.; Hussain, S.; Ditta, A.; Hashim, A.; Al-Arjani, A.-B.F.; et al. Promotion of Growth and Physiological Characteristics in Water Stressed Triticum aestivum Consequent to Foliar-application of Salicylic Acid. Water 2021, 13, 1316. [Google Scholar] [CrossRef]
- Whitehead, A. The Effects of Gibberellic Acid on Wild Type and Rosette Plants of the Species, Brassica rapa. University of Alabama at Birmingham. Lab Handout, 2008. Available online: www.regional.org.au/au/gcirc/canola/p-07.htm#P698_75989 (accessed on 16 September 2022).
- Rady, M.M.; Boriek, S.H.K.; El-Mageed, T.A.A.; El-Yazal, M.A.S.; Ali, E.F.; Hassan, F.A.S.; Abdelkhalik, A. Exogenous gibberellic acid or dilute bee honey boosts drought stress tolerance in Vicia faba by rebalancing osmoprotectants, antioxidants, nutrients, and phytohormones. Plants 2021, 10, 748. [Google Scholar] [CrossRef]
- Chunthaburee, S.; Sanitchon, J.; Pattanagul, W.; Theerakulpisut, P. Alleviation of salt stress in seedlings of black glutinous rice by seed priming with spermidine and gibberellic acid. Not. Bot. Horti Agrobot. Cluj-Napoca 2014, 42, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Estefan, G.; Sommer, R.; Ryan, J. Methods of Soil, Plant, and Water Analysis: A Manual for the West Asia and North Africa Region, 3rd ed.; ICARDA: Beirut, Lebanon, 2013; pp. 65–119. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.; Qayyum, A.; Razzaq, A.; Ahmad, M.; Mahmood, I.; Sher, A. Role of foliar application of salicylic acid and L-tryptophan in drought tolerance of maize. J. Anim. Plant Sci. 2012, 22, 768–772. [Google Scholar]
- Tkachuk, R. Oil and protein analysis of whole rapeseed kernels by near-infrared reflectance spectroscopy. J. Am. Oil Chem. Soc. 1981, 58, 817–822. [Google Scholar] [CrossRef]
- Thomas, C.R.; Maurice, S.C. Statistix 9, Managerial Economics; McGraw-Hill/Irwin: Columbus, OH, USA, 2008. [Google Scholar]
- Kwon, S.H.; Torrie, J.H. Heritability and interrelationship among traits of two soybean populations. Crop Sci. 1964, 4, 196–198. [Google Scholar] [CrossRef] [Green Version]
- Achard, P.; Cheng, H.; De Grauwe, L.; Decat, J.; Schoutteten, H.; Moritz, T.; Van Der Straeten, D.; Peng, J.; Harberd, N.P. Integration of plant responses to environmentally activated Phytohormonal signals. Science 2006, 311, 91–94. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Kurepin, L.V.; Reid, D.M. Growth and physiological responses of canola (Brassica napus) to three components of global climate change: Temperature, carbon dioxide, and drought. Physiol. Plant. 2006, 128, 710–721. [Google Scholar] [CrossRef]
- Rodríguez, A.A.; Stella, A.M.; Storni, M.M.; Zulpa, G.; Zaccaro, M.C. Effects of cyanobacterial extracellular products and gibberellic acid on salinity tolerance in Oryza sativa L. Saline Syst. 2006, 2, 1–4. [Google Scholar] [CrossRef]
- Rahman, M.; Nahar, M.; Sahariar, M.; Karim, M. Plant growth regulators promote growth and yield of summer tomato (Lycopersicon esculentum Mill.). Progress. Agric. 2015, 26, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Panda, B.; Bandyopadhyay, S.; Shivay, Y. Effect of irrigation level, sowing dates, and varieties on yield attributes, yield, consumptive water use, and water-use efficiency of Indian mustard (Brassica juncea). Indian J. Agric. Sci. 2004, 74, 339–342. [Google Scholar]
- Germchi, S.; Shekari, F.; Hassanpooraghdam, M.B.; Benam, M.; Shekari, F. Water deficit stress affects growth and some biochemical characteristics of rapeseed (Brassica napus L.). J. Food Agric. Environ. 2010, 8, 1127. [Google Scholar]
- Morrison, M.J.; Gutknecht, A.; Chan, J.; Miller, S.S. Characterizing canola pollen germination across a temperature gradient. Crop Pasture Sci. 2016, 67, 317. [Google Scholar] [CrossRef]
- Wu, W.; Ma, B.L.; Whalen, J.K. Enhancing rapeseed tolerance to heat and drought stresses in a changing climate: Perspectives for stress adaptation from root system architecture. Adv. Agron. 2018, 151, 87–157. [Google Scholar]
- Sánchez-Blanco, M.J.; Rodrıguez, P.; Morales, M.A.; Ortuño, M.; Torrecillas, A. Comparative growth and water relations of Cistusalbidus and Cistusmonspeliensis plants during water deficit conditions and recovery. Plant Sci. 2002, 162, 107–113. [Google Scholar] [CrossRef]
- Valentovic, P.; Luxova, M.; Kolarovic, L.; Gasparikova, O. Effect of osmotic stress on compatible solutes content, membrane stability and water relations in two maize cultivars. Plant Soil Environ. 2006, 52, 184. [Google Scholar]
- Mondal, S.K.; Khajuria, M.R. Genetic analysis for yield attributes in mustard. Environ. Ecol. 2000, 18, 1–5. [Google Scholar]
- Kamara, A.Y.; Menkir, A.; Badu–Apraku, B.; Ibikunle, O. The influence of drought stress on growth, yield, and yield components of selected maize genotypes. J. Agric. Sci. 2003, 141, 43–50. [Google Scholar] [CrossRef]
- Monneveux, P.; Sanchez, C.; Beck, D.; Edmeades, G. Drought tolerance improvement in tropical maize source populations. Crop Sci. 2006, 46, 180–191. [Google Scholar] [CrossRef]
- Schussler, J.; Westgate, M. Assimilate flux determines kernel set at low water potential in maize. Crop Sci. 1995, 35, 1074–1080. [Google Scholar] [CrossRef]
- Kamkar, B.; Daneshmand, A.; Ghooshchi, F.; Shiranirad, A.; Langeroudi, A.S. The effects of irrigation regimes and nitrogen rates on some agronomic traits of canola under a semiarid environment. Agric. Water Manag. 2011, 98, 1005–1012. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, J.; Duan, A.; Wang, J.; Shen, X.; Liu, X. Effects of different planting patterns on water use and yield performance of winter wheat in the Huang-Huai-Hai plain of China. Agric. Water Manag. 2007, 92, 41–47. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Abreu, M.E.; Munné-Bosch, S. Salicylic acid may be involved in the regulation of drought-induced leaf senescence in perennials: A case study in field-grown (Salvia officinalis L.) plants. Environ. Exp. Bot. 2008, 64, 105–112. [Google Scholar] [CrossRef]
- Korkmaz, A.; Uzunlu, M.; Demirkiran, A.R. Treatment with acetylsalicylic acid protects muskmelon seedlings against drought stress. Acta Physiol. Plant. 2007, 29, 503–508. [Google Scholar] [CrossRef]
- Habibi, G. Contrastive response of Brassica napus L. to exogenous salicylic acid, selenium and silicon supplementation under water stress. Arch. Biol. Sci. 2015; 67, 397–404. [Google Scholar]
- Si, P.; Mailer, R.J.; Galwey, N.; Turner, D.W. Influence of genotype and environment on oil and protein concentrations of canola (Brassica napus L.) grown across southern Australia. Aust. J. Agric. Res. 2003, 54, 397–407. [Google Scholar] [CrossRef]
- Champolivier, L.; Merrien, A. Effects of water stress applied at different growth stages to Brassica napus L. var. oleifera on yield, yield components, and seed quality. Eur. J. Agron. 1996, 5, 153–160. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M.; Anwar, F. Physicochemical attributes of seed oil from drought-stressed sunflower (Helianthus annuus L.) plants. Grasas Y aceites 2009, 60, 477–483. [Google Scholar]
- Dwivedi, S.; Nigam, S.; Rao, R.N.; Singh, U.; Rao, K. Effect of drought on oil, fatty acids, and protein contents of groundnut (Arachis hypogaea L.) seeds. Field Crop Res. 1996, 48, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Talaat, I.; El-Din, K. Physiological effect of putrescine and heat hardening on Nigella sativa L. plants. Int. J. Agric. Biol. 2005, 7, 358–362. [Google Scholar]
- Serafini-Fracassini, D. Cell cycle-dependent changes in plant polyamine metabolism. In Biochemistry and Physiology of Polyamines in Plants; CRC Press: Boca Raton, FL, USA, 1991; pp. 159–173. [Google Scholar]
- Brown, J.; Paliyath, G.; Thompson, J. Physiological mechanisms of plant senescence. In Plant Physiology; Academic Press: Cambridge, MA, USA, 1991; pp. 227–275. [Google Scholar]
- Khan, N.A.; Mir, R.; Khan, M.; Javid, S.; Samiullah. Effects of gibberellic acid spray on nitrogen yield efficiency of mustard grown with different nitrogen levels. Plant Growth Regul. 2002, 38, 243–247. [Google Scholar] [CrossRef]
- Wright, P.R.; Morgan, J.M.; Jessop, R.S.; Cass, A. Comparative adaptation of canola (Brassica napus) and Indian mustard (Brassica Juncea) to soil water deficit: Yields and yield components. Field Crop. Res. 1995, 42, 1–13. [Google Scholar] [CrossRef]
- Enferad, A.; Poustini, K.; MajnounHosseini, N.; Taleei, A.; Khajeh-Ahmad-Attari, A. Physiological responses of Rapeseed (Brassica napus L.) varieties to salinity stress in the Vegetative Growth Phase. J. Sci. Technol. Agric. Natur. Resour. 2004, 4, 103–114. [Google Scholar]
- Ozer, H.; Oral, E.; Dogru, U. Relationship between yield and yield components on currently improved spring rapeseed cultivars. Turk. J. Agric. For. 1999, 23, 603–607. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Replication | Variety (V) | Foliar (F) | Drought (D) | V × F | V × D | F × D | V × F × D |
|---|---|---|---|---|---|---|---|---|
| DF | 2.00 | 1.00 | 2.00 | 1.00 | 2.00 | 1.00 | 2.00 | 2.00 |
| PH | 0.25 | 11.67 ** | 760.08 ** | 2.01 * | 0.36 ns | 57.51 ** | 25.69 ** | 61.69 ** |
| CHA | 0.00 | 0.02 ** | 0.19 ** | 0.01 ** | 0 ns | 0.02 ** | 0 ns | 0 ns |
| CHB | 0.00 | 0.03 ** | 0.39 ** | 0.49 ** | 0.00 * | 1.38 ** | 0.01 ** | 0.03 * |
| TCH | 0.00 | 2.11 ** | 0.55 ** | 1.42 ** | 0.02 ** | 3.03 ** | 0.1 ** | 0.03 ** |
| NSPP | 0.27 | 2.51 ** | 56.9 ** | 6.67 ** | 0.09 ns | 5.06 ** | 0.76 * | 0.56 * |
| SPS | 0.01 | 3.58 ** | 43.1 ** | 0.23 * | 0.13 ns | 10.84 ** | 0.55 ** | 0.13 ns |
| SL | 0.01 | 0.0 ns | 9.01 ** | 0.0 ns | 0.02 ns | 0.03 ns | 0.04 ns | 0.17 * |
| CARC | 0.19 | 31.51 ** | 498.39 ** | 58.93 ** | 9.86 ** | 37.5 ** | 4.39 ** | 8.05 ** |
| RWC | 0.07 | 1854.74 ** | 941.13 ** | 7.77 ** | 0.13 ns | 295.73 ** | 77.06 ** | 5.17 ** |
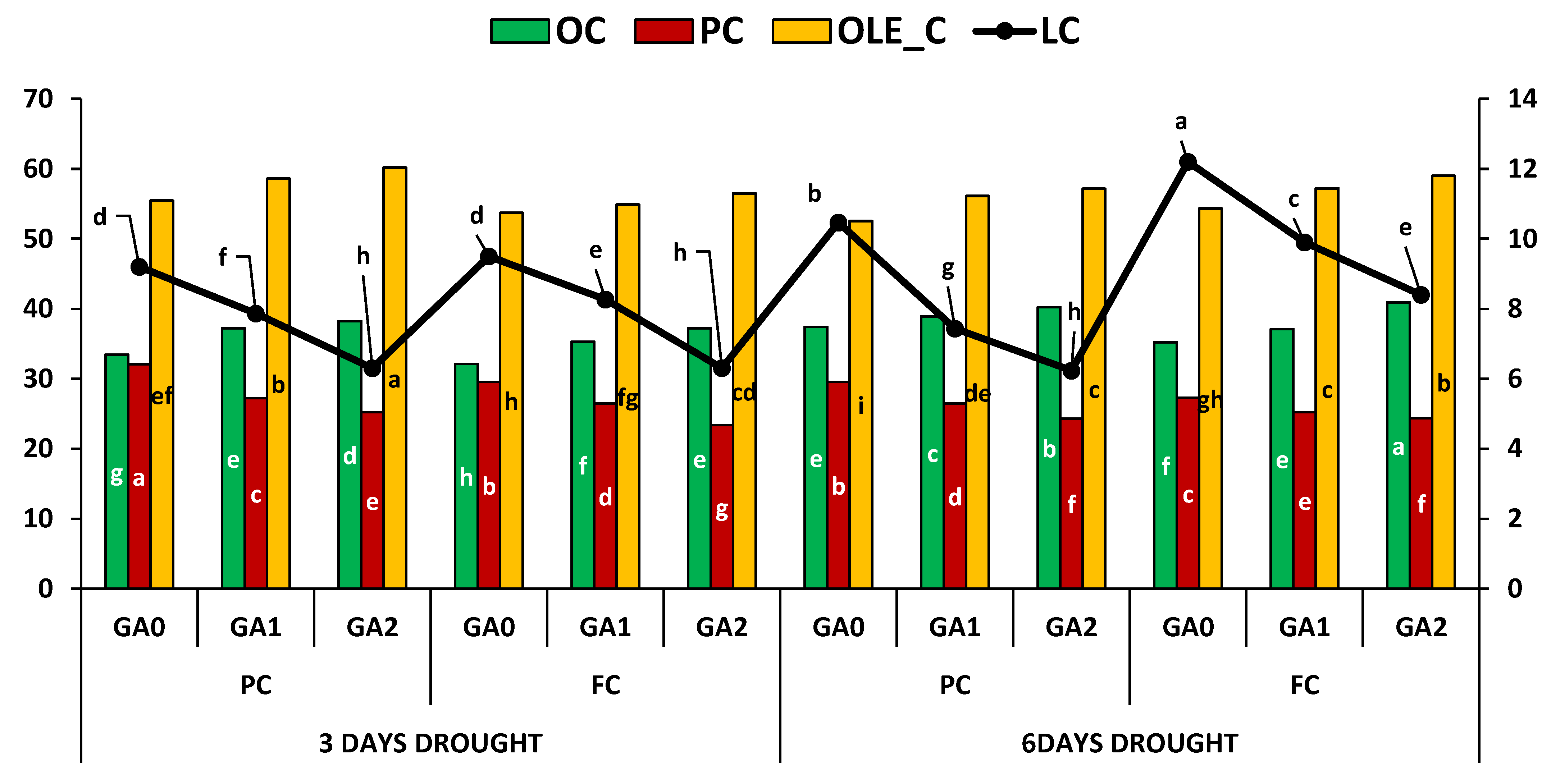
| OC | 0.11 | 13.94 ** | 64.04 ** | 65.61 ** | 2.7 ** | 0.22 ns | 2.35 ** | 1.26 ** |
| PC | 0.02 | 18.78 ** | 85.43 ** | 11.33 ** | 2.06 ** | 0.75 * | 4.29 ** | 1.2 ** |
| OLE_C | 0.19 | 4.91 ** | 54.55 ** | 2.3 ** | 1.47 ** | 47.84 ** | 1.02 * | 0.85 * |
| LC | 0.02 | 12.48 ** | 37.63 ** | 12.96 ** | 0.15 * | 8.03 ** | 1.51 ** | 0.12 * |
| TSW | 0.08 | 0.01 ns | 9.38 ** | 0.04 ns | 0.04 ns | 0 ns | 0.00 ns | 0.11 ns |
| YP | 0.00068 | 0.001 ns | 0.134 ** | 0.008 ** | 0.001 ns | 0.001 ns | 0.001 ns | 0.001 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elahi, N.N.; Raza, S.; Rizwan, M.S.; Albalawi, B.F.A.; Ishaq, M.Z.; Ahmed, H.M.; Mehmood, S.; Imtiaz, M.; Farooq, U.; Rashid, M.; et al. Foliar Application of Gibberellin Alleviates Adverse Impacts of Drought Stress and Improves Growth, Physiological and Biochemical Attributes of Canola (Brassica napus L.). Sustainability 2023, 15, 78. https://doi.org/10.3390/su15010078
Elahi NN, Raza S, Rizwan MS, Albalawi BFA, Ishaq MZ, Ahmed HM, Mehmood S, Imtiaz M, Farooq U, Rashid M, et al. Foliar Application of Gibberellin Alleviates Adverse Impacts of Drought Stress and Improves Growth, Physiological and Biochemical Attributes of Canola (Brassica napus L.). Sustainability. 2023; 15(1):78. https://doi.org/10.3390/su15010078
Chicago/Turabian StyleElahi, Nosheen Noor, Sadia Raza, Muhammad Shahid Rizwan, Bedur Faleh A. Albalawi, Muhammad Zubair Ishaq, Hafiz Munir Ahmed, Sajid Mehmood, Muhammad Imtiaz, Umer Farooq, Muhammad Rashid, and et al. 2023. "Foliar Application of Gibberellin Alleviates Adverse Impacts of Drought Stress and Improves Growth, Physiological and Biochemical Attributes of Canola (Brassica napus L.)" Sustainability 15, no. 1: 78. https://doi.org/10.3390/su15010078