Investigation of Peptide Toxin Diversity in Ribbon Worms (Nemertea) Using a Transcriptomic Approach


A.V. Zhirmunsky National Scientific Center of Marine Biology, Far Eastern Branch, Russian Academy of Sciences, 690041 Vladivostok, Russia
*
Author to whom correspondence should be addressed.
Toxins 2022, 14(8), 542; https://doi.org/10.3390/toxins14080542
Submission received: 31 May 2022
/
Revised: 18 July 2022
/
Accepted: 6 August 2022
/
Published: 8 August 2022
(This article belongs to the Special Issue Evolution, Genomics and Proteomics of Venom)
Abstract
:Nemertea is a phylum of nonsegmented worms (supraphylum: Spiralia), also known as ribbon worms. The members of this phylum contain various toxins, including peptide toxins. Here, we provide a transcriptomic analysis of peptide toxins in 14 nemertean species, including Cephalothrix cf. simula, which was sequenced in the current study. The summarized data show that the number of toxin transcripts in the studied nemerteans varied from 12 to 82. The most represented groups of toxins were enzymes and ion channel inhibitors, which, in total, reached a proportion of 72% in some species, and the least represented were pore-forming toxins and neurotoxins, the total proportion of which did not exceed 18%. The study revealed that nemerteans possess a much greater variety of toxins than previously thought and showed that these animals are a promising object for the investigation of venom diversity and evolution, and in the search for new peptide toxins.
Key Contribution: The transcriptomic analysis revealed 12 to 82 toxin transcripts in nemerteans. Nemertean species belonging to the same class were grouped together according to the contained toxins.
1. Introduction
Animal poisons and venoms are potential sources of new toxic proteins and peptides, representatives of which have already found applications in many areas as therapeutic agents and physiological tools [1,2,3,4]. A large number of these proteins and peptides have been found in marine organisms, and the majority of studies to date have been focused on toxins from cone snails, sea anemones, fish, jellyfish, sea stars, hydras, sea urchins, sea hares, etc. [5,6]. In recent decades, due to the increasing popularity of next-generation sequencing (NGS) technologies, more results from the genome/transcriptome sequencing of neglected animals have appeared in free databases, which makes it possible to use them as a source of new valuable biological information. This kind of study elucidates the ecological aspects of animals’ adaptation and evolution. As a rule, toxic peptides are part of multicomponent mixtures, i.e., venoms or poisons, which have been studied in many animals such as snakes [7], scorpions [8], spiders [9], sea anemones [10], and jellyfishes [11]. However, the study of less popular animals is no less important and allows us to not only find new bioactive peptides, but also to study the evolution of poison and to better understand adaptation processes by conducting comparative analyses of the compositions of animal toxins [12].
One of these little-studied but promising sources of toxins is the marine worms of the Nemertea phylum consisting of more than 1300 species and subdivided into three phylogenetic groups: Palaeonemertea, Pilidiophora, and Hoplonemertea [13,14]. Nemerteans are a rich source of various toxic compounds [15]. Despite the absence of specialized glands for venom and poison secretion, they contain a multicomponent cocktail of toxins that provides them with protection and allows efficient predation [15]. To date, there have been only four works devoted to the screening of toxins using these approaches in nemerteans. The first toxin screening work was carried out in 2014 by Whelan and colleagues on the transcriptomes of nine nemertean species; the number of putative toxin genes found varied from three to seven in different species [16]. More recent works have demonstrated a greater number of toxin genes in nemerteans. A study of the genome of the heteronemertean species N. geniculatus revealed 32 putative toxin genes [17]. In 2020, a proteo-transcriptomic analysis of the hoplonemertean Am. lactifloreus was carried out by von Reumont and colleagues, and resulted in the identification of 26 peptides that potentially play a role in prey capture, immobilization, and predigestion [18]. The most recent study (carried out in 2022 by Verdes and colleagues), devoted to the proteo-transcriptomic analysis of A. valida venom, demonstrated the presence of 85 putative toxins, classified as potentially predatory, defensive, or having dual functions [19]. Based on the results of Luo and colleagues [17], von Reumont and colleagues [18], and Verdes and colleagues [19], it can be assumed that nemertean transcriptomes may contain a much greater variety of toxin-like transcripts than what was shown by Whelan and colleagues [16].
To date, most of the available studies provided information on toxins in a limited number of ribbon worms species and did not reveal the general principles of toxin content in different nemertean classes. The present research was devoted to revealing the toxin transcripts in 14 species of nemerteans belonging to three classes (Palaeonemertea, Pilidiophora, and Hoplonemertea) using the transcriptomic approach, including Cephalothrix cf. simula, which was sequenced in the current study. The data obtained allowed the discovery of general trends in the diversity of peptide toxins in nemerteans.
2. Results
2.1. Transcriptomes Assembly
The transcriptomes of 12 nemertean species (A. lactifloreus, Malacobdella grossa, Paranemertes peregrina, Carinoma hamanako, Cephalothrix hongkongiensis, Tubulanus polymorphus, Baseodiscus unicolor, Hubrechtella ijimai, Lineus longissimus, Lineus ruber, Lineus sanguineus, and Riseriellus occultus) were assembled and annotated using reads, which were downloaded from the SRA. The previously published N. geniculatus transcriptome was annotated [17], and the assembly and annotation of the Ce. cf. simula sequences obtained in the current work were carried out. Table 1 presents data processing statistics. The quality of the final assembly was assessed by BUSCO and ranged from 63.9% (P. peregrina) to 93.7% (L. longissimus). The largest number of nonredundant annotated unique transcripts was obtained for the Ce. cf. simula transcriptome, which was assembled using a hybrid approach using reads from two platforms, Illumina and MinION Oxford Nanopore. This resulted in 25,895 open reading frames (ORFs) with unique BLAST hits, with an average number of 11,564 ORFs with unique BLAST hits in the remaining 13 nemertean species. ORFs with unique BLAST hits were obtained by removing redundant contigs with identical annotations according to the method proposed by Ono and colleagues [20]. This approach allowed reduction of transcriptome redundancy by 17.8% (M. grossa) to 67.6% (Ce. cf. simula).
2.2. Putative Toxin Transcripts in Transcriptomes
To identify toxins, transcripts were annotated using the Tox-Prot and SWISS-PROT/UniProt (E-value 10−6) databases. Two of the resulting annotations were compared, and the annotation with the highest E-value was considered significant (Supplementary File S2).
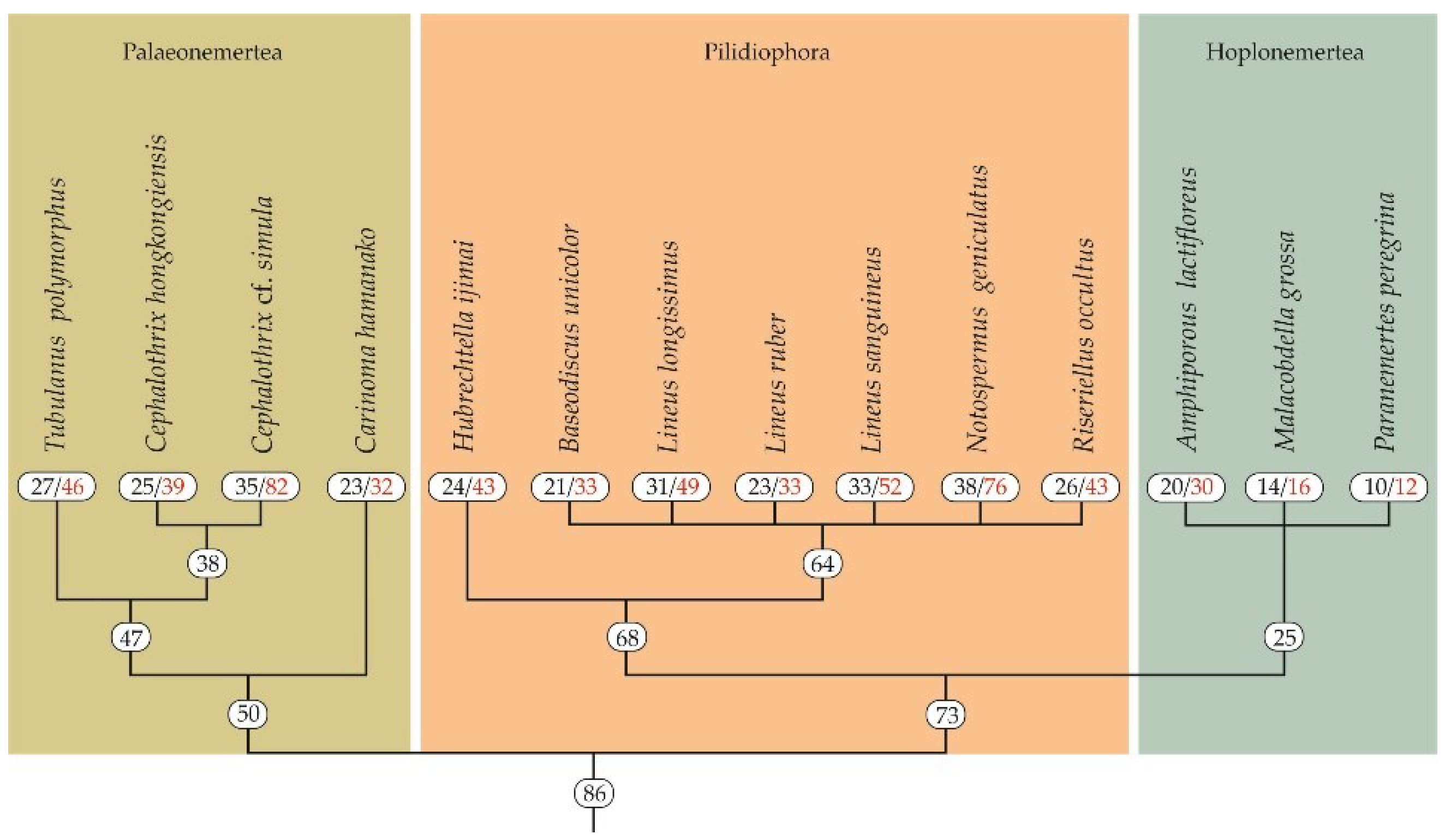
A total of 14 nemerteans belonging to three classes (Palaeonemertea, Pilidiophora, and Hoplonemertea) were shown to possess from 12–16 (in P. peregrina and M. grossa, respectively) to 76–82 (in N. geniculatus and Ce. cf. simula, respectively) toxin transcripts (Figure 1). In palaeonemertean species, on average, 49 transcripts were found, from 32 in Ca. hamanako to 82 in Ce. cf. simula. The average number of toxins in pilidiophorans was 47, ranging from 33 transcripts in B. unicolor and L. ruber to 76 transcripts in N. geniculatus (Figure 1). The number of toxin transcripts in three annotated hoplonemerteans were 12, 16, and 30 in P. peregrina, M. grossa, and A. lactifloreus, respectively, whereas the average number was about 19 transcripts.
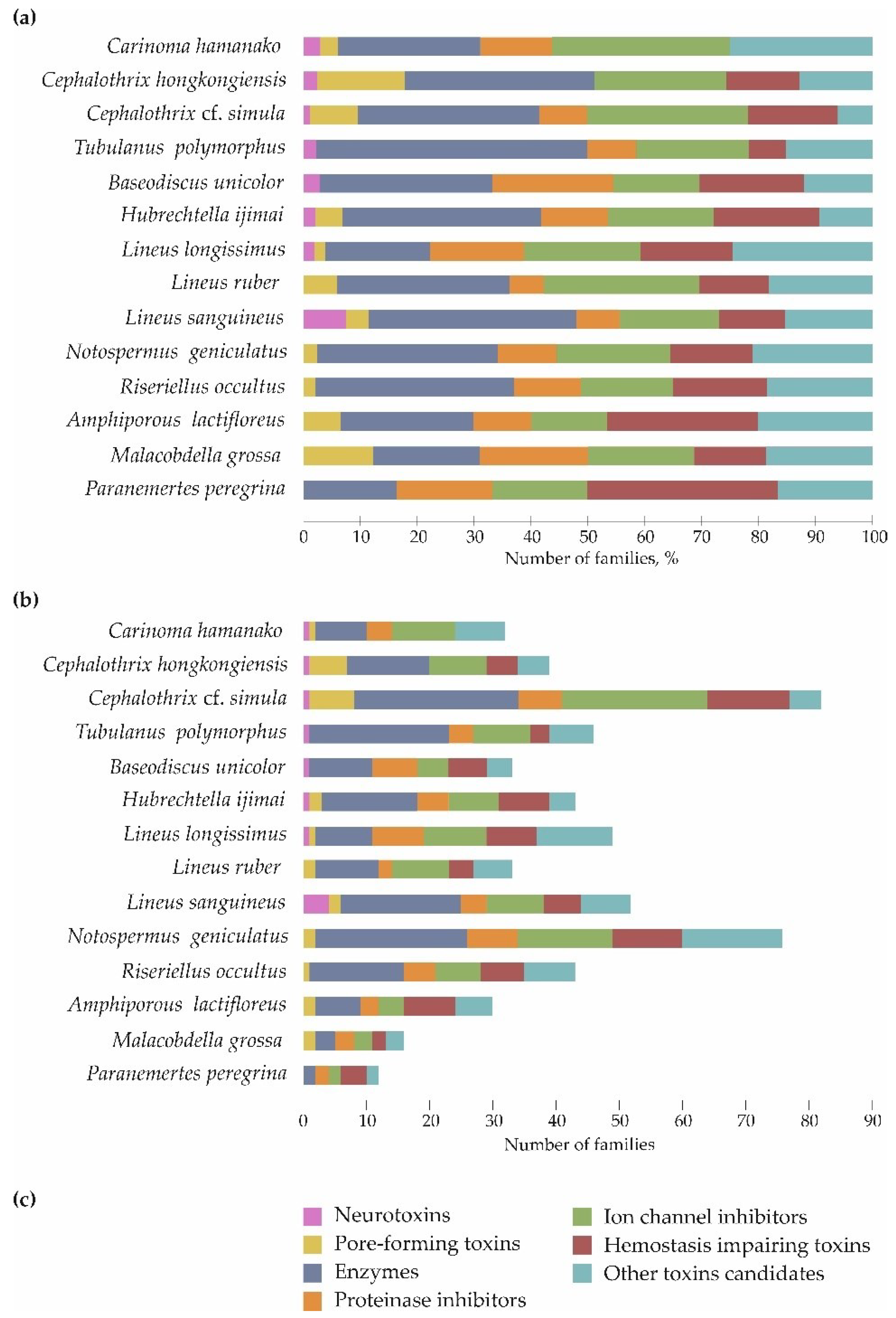
Toxin transcripts were divided into seven groups and assigned to 86 families (Figure 2); up to 14 transcripts belonged to each family. The toxins families were determined using UniProtKB/SWISS-PROT [24] Family & Domains section that provides information about the sequence similarity with other proteins. The most represented group was enzymes, which included 32 families, accounting for 16.7–18.8% (P. peregrina and M. grossa, respectively) to 36.5–47.8% (L. sanguineus and T. polymorphus, respectively) of all toxin transcripts. The enzyme group was followed by a group of ion channel inhibitors (10 families), where the toxin transcripts occupied from 15.2–16.3% (B. unicolor and R. occultus, respectively) to 30.3–39.4% (L. ruber and Ca. hamanako, respectively) of all toxin transcripts. These two groups, as well as the group of other toxin candidates, were present in the transcriptomes of all 14 species of ribbon worms. The least represented group was neurotoxins; their toxin transcripts were found in 6 out of 14 nemerteans and only in the Palaeonemertea and Pilidiphora classes. The largest number of neurotoxin families (three families) was found in L. sanguineus; and five other species bore one family each. Most of the studied species were shown to possess unique toxin families belonging to all toxin groups, except pore-forming toxins. The largest number of unique toxins, four families, was found in L. longissimus, L. sanguineus, and N. geniculatus; no unique toxins were found in Ca. hamanako and M. grossa (Figure 2).
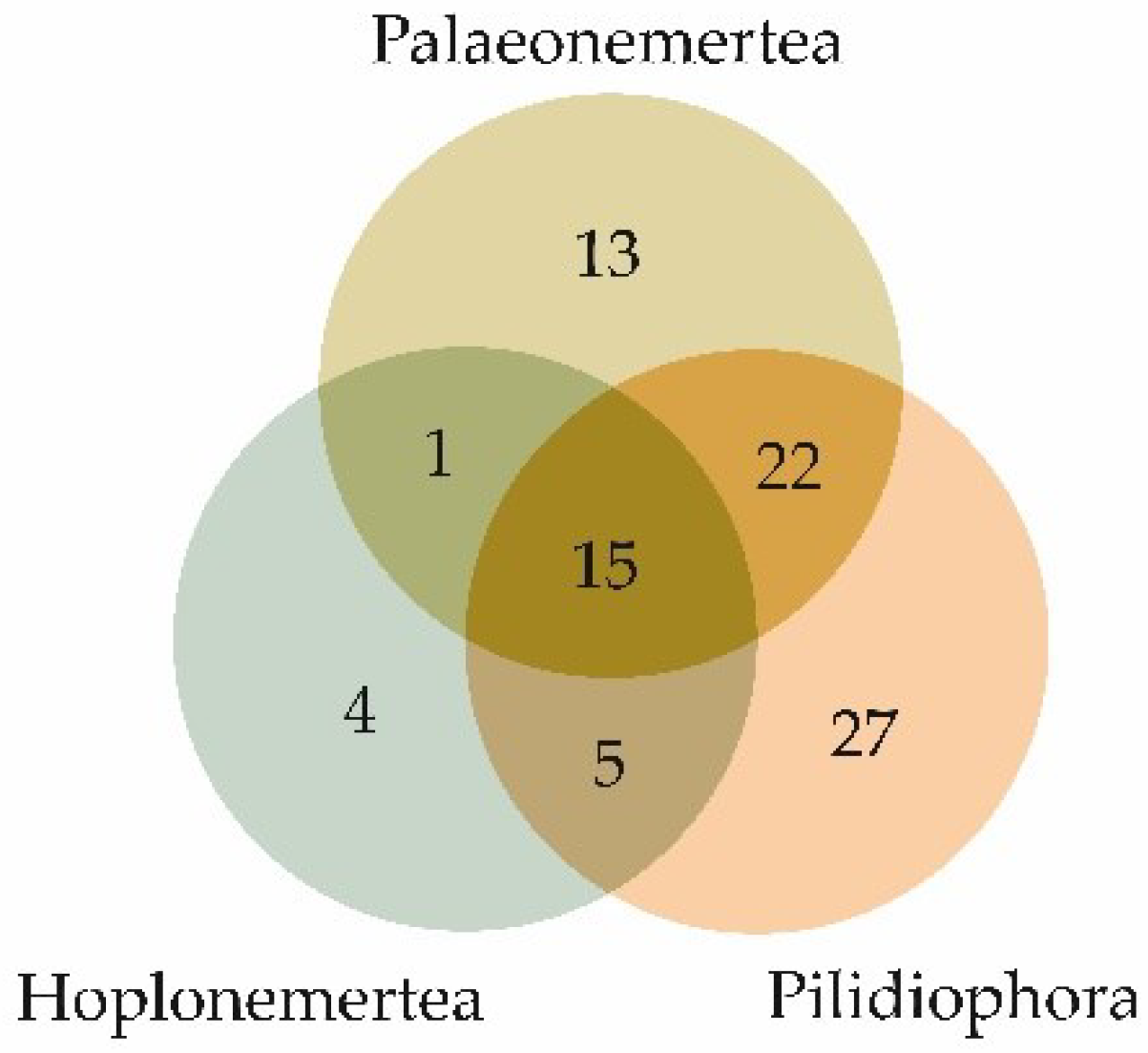
The qualitative compositions of the toxins of the three nemertean classes—Palaeonemertea, Pilidiophora, and Hoplonemertea—had both similarities and differences. Figure 3 demonstrates a Venn diagram of the toxin families’ overlaps between nemertean classes. All the classes had toxin families belonging to all the groups of toxins except neurotoxins, which were only found in palaeonemerteans (one family) and pilidiphorans (four families); toxin families of this group did not intersect between the two classes of nemerteans (Table 2). The largest number of common toxin families between the two classes was found in Palaeonemertea and Pilidiophora species, most of which belonged to the enzyme group. Moreover, most of the unique toxin families of all three nemertean classes were also enzymes (from 37 to 50%).
2.3. Nemertean-Specific Peptide Toxin Transcripts
Previous studies have described nemertean-specific peptide toxins [15], including cytotoxins AI–AIV, neurotoxins BI–BIV (Cerebratulus lacteus) [25], parbolysin (Parbolasia corrugatus) (Berne et al. 2003), and nemertides α-1, α-2, and β (L. longissimus) [26]. It was subsequently found that parbolysin has high homology with C. lacteus cytotoxin A-III [27]. Two nemertean-specific toxins, cytotoxin A-III and nemertide α-1, were identified in the current study. Cytotoxin A-III was detected in L. longissimus, L. ruber, N. geniculatus, H. ijimai, L. sanguineus, and R. occultus (for the first time in the last three). The nemertide α-1 toxin was identified in L. ruber, L. sanguineus, and R. occultus, as in the study by Jacobsson and colleagues [26]. The failure to identify other nemertean-specific toxins might have been caused by different transcriptome assembly techniques.
In a recent study by von Reumont and colleagues [18], non-nemertean-specific toxin transcripts were also identified, including those that were plancitoxin-like, originally isolated from crown-of-thorns starfish, and those that were actitoxin-like, isolated from sea anemones, named U-nemertotoxin-1 and U-nemertotoxin-2, respectively. According to their study, both toxins were typical for nemerteans representing Palaeonemertea, Pilidiophora, and Hoplonemertea. U-nemertotoxin-1 transcripts were found in the Am. lactifloreus and N. geniculatus proboscis transcriptomes and the full-body transcriptomes of seven species from all three nemertean classes: Ce. hongkongiensis, Cephalothrix linearis, Cerebratulus marginatus, T. polymorphus, M. grossa, P. peregrina, L. lacteus, L. longissimus, and L. ruber. The transcripts of U-nemertotoxin-2 were found in the proboscis transcriptomes of Am. lactifloreus and N. geniculatus. In the current study, transcripts corresponding to U-nemertotoxin-1 were found in most nemertean species, except for P. peregrina and H. ijimai. Transcripts presumably related to U-nemertotoxin-2 were identified in 12 out of 14 nemertean species, with the exceptions being L. sanguineus and H. ijimai.
2.4. Assessment of Distance between Species Based on the Presence/Absence of Toxin Families
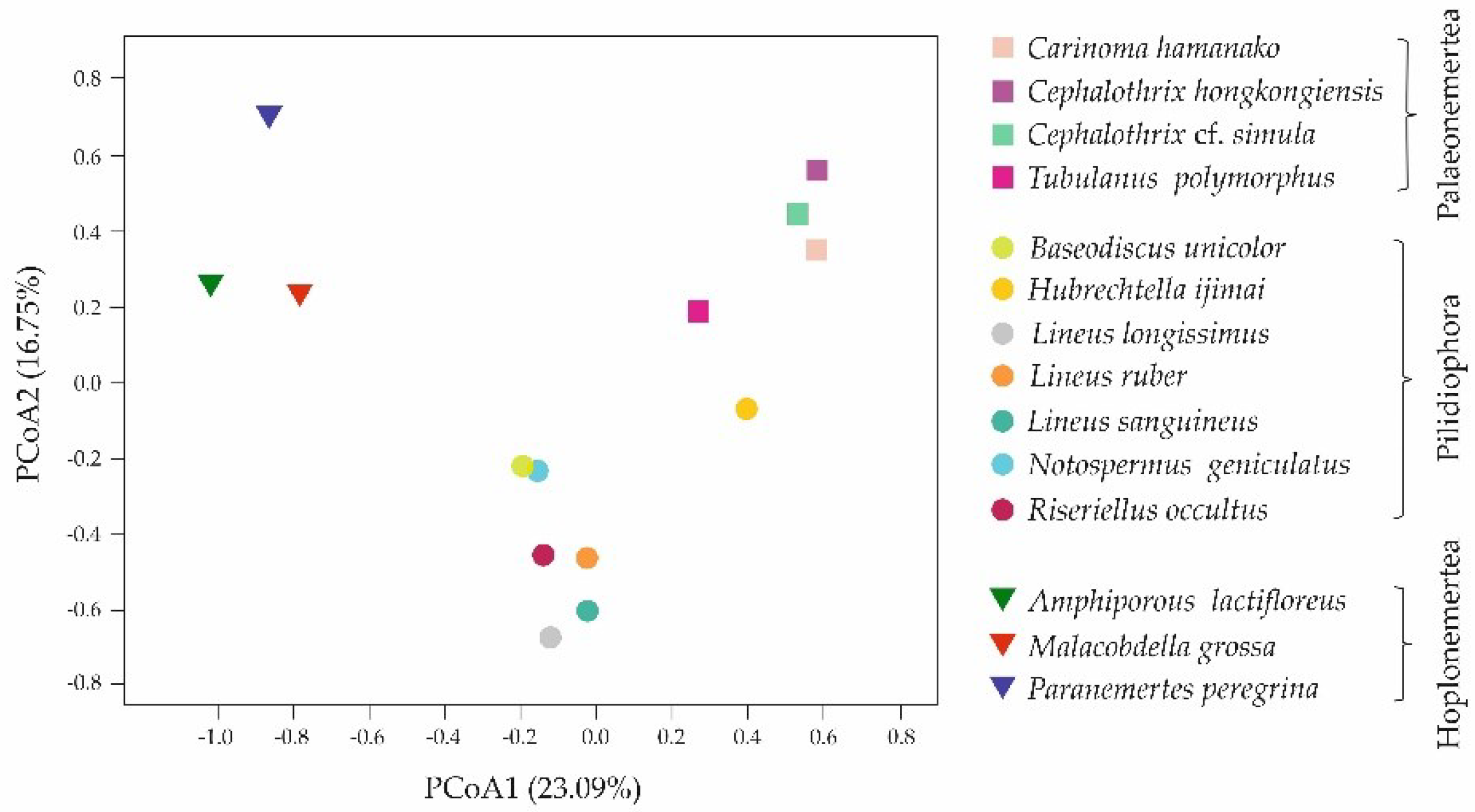
Based on the presence/absence of toxin families in 14 nemertean transcriptomes, principal coordinate analysis (PCoA) was performed using Jaccard distance, which expresses the distance between species (pseudo-F = 3.099, p = 0.0001, PERMANOVA, 10,000 permutations in each test) (Figure 4). The results presented in the figure demonstrate that nemertean species were grouped according to the classes to which they belong.
2.5. Preliminary Assessment of Toxin Transcripts Expression
The toxin transcripts abundance was quantified for preliminary assessment of toxin expression in all nemertean species using the Salmon tool (Tables S1–S14 in Supplementary File S1) and expressed as a percentage; the transcripts with the highest expression, which accounted for 90% of the total expression of the species, were selected (Table 3). For more accurate toxin gene expression levels, the same specimen-preparing conditions should be provided. In all the studied nemerteans, among the transcripts with the highest expression, there were toxins from all seven groups, except for the hoplonemertean species, in which six groups were identified. For the palaeonemerteans, 90% of the total expression was from 10 to 22 toxin transcripts, and the average expression was the highest in representatives of the neurotoxin group (up to 30.3% in Ce. hongkongiensis, from the neurotoxin 20 family) and ion channel inhibitors (up to 33.4% in Ca. hamanako, from the CRISP family) (Table 3). Between 5 and 23 toxin transcripts of pilidiophorans accounted for 90% of the expression, and the highest average expression was shown by representatives of the neurotoxin group (up to 35.3% in L. longissimus, from the neurotoxin 02 (plectoxin) family. 02 (plectoxin) subfamily) and proteinase inhibitors (up to 40.2% in L. sanguineus, from the venom Kunitz-type family). Among the Hoplonemertea members, between five and eight transcripts from 90% of the most expressed toxins were identified. The most expressed were other toxin candidates (up to 47.8% in A. lactifloreus, from the TCTP family) and proteinase inhibitors (up to 56.3% in P. peregrina, from the venom Kunitz-type family sea anemone type 2 potassium channel toxin subfamily) (Table 3).
Despite the grouping of nemertean species according to their classes demonstrated by PCoA (Figure 4), the abundance of the toxin transcripts in closely related species varied (Table 4). This was also true for major transcripts, which made up 50% of the total toxin expression. Three species from the Lineus genus—L. longissimus, L. ruber, and L. sanguineus—with three, two, and two major toxins, respectively, showed more differences than similarities; the major toxins comprised the Kunitz-type family (proteinase inhibitor group and ion channel inhibitors) and did not contain pore-forming toxins, enzymes and hemostasis-impairing toxins. Two Cephalothrix species possessed three major toxins each and were similar only in terms of the presence of MACPF domain-containing toxins and the absence of hemostasis-impairing toxins (Table 4).
3. Discussion
Nemerteans possess various toxins with defensive and offensive functions. These include pyridine toxins (anabaseine, nemertelline, 2,3’-bipyridyl, 3-methyl-2,3’-bipyridyl), tetrodotoxin and its analogues (TTXs), and various peptide toxins. According to current data, pyridine toxins are characteristic of hoplonemerteans [28]. The highest concentration and greatest variety of TTXs are specific to palaeonemerteans [29,30,31,32,33], although trace concentrations have been found in pilidiophorans and hoplonemerteans [32]. Peptide nemertean-specific toxins have been identified in Pilidiophora class representatives [34], and transcripts of non-nemertean-specific toxins have been found in all nemertean classes [16]. Recently, due to the fast development of NGS techniques, the transcriptomic approach has become popular, leading to the complete and efficient identification of peptides and their expression evaluation, which permits the comparison of the mechanisms by which toxins are used in animals. In the current study, we reassembled and annotated the transcriptomes of Whelan and colleagues [16] (M. grossa, P. peregrina, T. polymorphus, Ce. hongkongiensis, L. longissimus, L. ruber, and Am. lactifloreus [12]). The transcriptomes of Ca. hamanako, B. unicolor, H. ijimai, L. sanguineus, and R. occultus were assembled and annotated from the reads deposited in the SRA (NCBI). The previously published transcriptome of N. geniculatus [17] was processed and annotated. In addition, in the present study, the Ce. cf. simula transcriptome was sequenced, assembled, and annotated for the first time. For all of these transcriptomes, the content of toxin transcripts was evaluated; as a result, a total of 588 toxin transcripts were identified, which were divided into 86 families and assigned to seven groups of toxins according to the annotations from the UniProtKB/Swiss-Prot and Tox-Prot databases (Tables S1–S14 in Supplementary File S1). These groups were neurotoxins (5 families), pore-forming toxins (5 families), enzymes (31 families), proteinase inhibitors (10 families), ion channel inhibitors (9 families), hemostasis-impairing toxins (4 families), and other toxin candidates (22 families).
The function of animals’ toxic cocktails is reflected by the composition of toxins and their mechanism of action. The mixtures used to deter predators consist predominantly of compounds that induce an immediate reaction and interfere with fast-acting physiological processes such as nerve transmission. Consequently, many defensive poisons contain toxins that quickly cause paralysis by blocking neuromuscular receptors or acting on pain receptors, causing instant and intense pain [35]. At the same time, venoms used to subdue prey are more diverse in the composition and physiological effects of their toxins [36]. In representatives of all nemertean classes, a mixture of toxins with different activities have been identified—Palaeonemertea and Pilidiophora species contained all seven groups of toxins, and Hoplonemertea species contained six groups; no neurotoxins were found in the latter. Presumably, some of them can play the role of a repellent agent to protect against predators, and others can be used during hunting as immobilizing agents or digestive enzymes. To specify the peptide toxins role for nemerteans, more detailed investigation is necessary to carry out. One of the directions is proteotranscriptomic differential gene expression analyses. To date, there are two articles, devoted to investigation of peptides in mucus, covered the nemertean body and proboscis, a specific weapon organ, demonstrating the characteristics of toxins function based on their expression patterns and proteomic distribution [18,19]. According to Verdes with colleagues, proteins of Antarctonemertes valida with insulin-like growth factor-binding domain (identified in the current research in R. occultus, A. lactifloreus, Ca. hamanako, Ce. hongkongiensis, Ce. cf. simula, T. polymorphus, B. unicolor, H. ijimai, L. longissimus, L. ruber, L. sanguineus, N. geniculatus) and galactose-binding domain (identified in the current research in R. occultus) (Supplementary File S1) were detected in proboscis and were suggested to be a response for predation; protein with Kunitz domain, identified in all nemerteans studied in the current research except for Ce. hongkongiensis (Supplementary File S1), was detected in whole specimen, mucus and proboscis, and presumably had dual functions—predatory and defensive [19]. The research of von Reumont with colleagues provided proteotranscriptomic analysis of Am. lactifloreus mucus and found out the secretion of protein toxins, also identified in the current research: lectin (nattectin-like) (identified in R. occultus), U-actitoxin (identified in Ca. hamanako, Ce. cf. simula, T. polymorphus, L. ruber, N. geniculatus), metalloproteinase M12A (identified in L. sanguineus, N. geniculatus, A. lactifloreus), metalloproteinase M12B (identified in Ce. hongkongiensis, Ce. cf. simula, T. polymorphus, H. ijimai, L. longissimus, L. sanguineus, N. geniculatus, M. grossa), L-amino acid oxidase (identified in Ce. hongkongiensis, Ce. cf. simula, B. unicolor, L. longissimus, L. ruber, L. sanguineus, N. geniculatus, R. occultus), plancitoxin (identified in Ca. hamanako, Ce. hongkongiensis, Ce. cf. simula, T. polymorphus, B. unicolor, L. longissimus, L. ruber, L. sanguineus, N. geniculatus, R. occultus, A. lactifloreus, M. grossa) (Supplementary File S1). Presumably, identified toxins secreted in the mucus could play a defensive role and contribute to predation and the paralysis of prey by facilitating the action of other components of a toxic cocktail [18]. Identified in An. valida and Am. Lactifloreus, toxic peptides with predatory and/or defensive functions were detected in up to 13 nemertean species, studied here, and the common role of peptides for these species may be assumed.
Based on the presence or absence of toxin families, we analyzed the distance between 14 nemertean species for the first time, demonstrating the grouping of species within classes (Figure 4). The largest number of common toxin families between the two classes was found in Palaeonemertea and Pilidiophora species; the toxins families’ profile of Hoplonemertea species was the most unique (Figure 4, Table 2). The variety of toxin composition of nemertean classes may be caused by their different feeding ecology, including diet preferences and hunting strategies. Thus, it was revealed that palaeonemerteans and pilidiophorans within classes and species individually are characterized by a wide range of potential types of prey: palaeonemerteans prefer nematodes, oligochaetes, polychaetes, and other nemerteans, and pilidiophorans prefer all of the above and additionally bivalves and crustaceans [37,38,39,40,41]. On the contrary, for the Hoplonemertea class, small range of victims is typical, most species prefer one systematic group, mainly crustaceans, as prey, and reject others [38,40,42,43]. Therefore, it could be assumed that the diversity of peptide toxins depends on diversity of potential prey, and since diet of hoplonemerteans is limited to one type of victim, they do not require a great variety of toxic agents. In the case of palaeonemerteans and pilidiophorans, a wide range of potential victims may result in a wide range of toxins, while each type of animal can be affected by a specific toxin. An assumption about the relation between toxin composition and diet preferences was put forth by Verdes and colleagues, due to the revealing of toxin specificity for different nemertean classes [19]. The hunting strategy of hoplonemerteans also increases the discrepancy with other nemertean classes: their proboscis is armed with a stylet that pierces the victim and directly injects the venom cocktail, while Palaeonemertea and Pilidiophora representatives wrap the proboscis around the object without piercing [44,45]. This aspect can also be associated with qualitative composition of toxins and may explain the absence of peptide neurotoxins in the representatives of hoplonemerteans—since the prey capture is accompanied by body piercing, its immobilization is not required.
Despite the similar qualitative composition of toxins in representatives of one class according to PCoA (Figure 4), their quantitative compositions variates greatly. The preliminary expression of toxin transcripts was measured, and it was found that the abundance of the same toxin families differed significantly even in closely related species (Table 4). Thus, in three representatives of Lineus genius, two common toxin groups were identified within major toxins between L. longissimus and L. sanguineus (neurotoxins, proteinase inhibitors), and L. longissimus and L. ruber (ion channel inhibitors) (Table 4). This may indicate differences in diet preferences of each species: Lineus species live in the same area; however, the victims are distinguishing [37,38,40,46]. The abundance of major toxins in another closely related species—Ce. hongkongiensis and Ce. cf. simula—was similar for pore-forming toxins and varied significantly for neurotoxins, enzymes (presented in Ce. hongkongiensis) and proteinase inhibitors, ion channel inhibitors (presented in Ce. cf. simula) (Table 4). The victim types, preferred by this nemertean species, have not been studied, therefore, correlation between diet and toxins composition could not be established. However, the variability in expression levels for the same toxin families can also result from different RNA preparation conditions: Ce. cf. simula RNA was extracted from the middle of the nemertean body and did not contain proboscis, and Ce. hongkongiensis RNA sample was obtained from three individuals and the tissue or body parts were not mentioned, there was no information about proboscis presence (Table 1). The same situation was observed within hoplonemerteans, where all transcriptomes were obtained from dissimilar tissues (Table 1), which correlates with the absence of major toxins overlapping between them (Table 4). Nemerteans toxins are thought to be secreted by glandular cells located in the epidermis of the integument for potential use against predators, and by cells located in the proboscis epidermis to contribute to prey capture [15]. Depending on toxins’ functions, their expression levels in these organs can be different [19]. The provided toxin transcripts abundancy estimation was preliminary and needs to be evaluated using the same tissue type and library preparation conditions.
4. Conclusions
Resulting from the transcriptomic analysis, high diversity of toxins and general trends in the distribution of peptide toxins within Nemertea phylum were revealed. The principal coordinate analysis of the distance between 14 nemertean species based on the presence/absence of toxin families in transcriptomes demonstrated that nemertean species were grouped according to the classes to which they belong—Palaeonemertea, Pilidiophora, and Hoplonemertea. The qualitative comparison of the toxin composition of the three nemertean classes showed the toxin families’ overlaps between nemertean classes; the largest number of common toxin families between the two classes was found in Palaeonemertea and Pilidiophora species. The correlation between number of common toxins with evolution distance between the classes is a question for further investigation. Palaeonemertea and Pilidiophora representatives, as the nemerteans with the largest number of toxin transcripts, may be the most promising objects for future studies. The results obtained point to the need for further study of the toxic composition of nemerteans, including proteo-transcriptomic analysis, in order to clarify the spectrum of toxins and study their expression and localization, as well as to search for new, unstudied toxic peptides.
5. Materials and Methods
5.1. Animal Collection
Specimens of the nemertean species Ce. cf. simula were collected in October 2019 and August 2020 from rhizoids of Saccharina sp. in the Spokoynaya Bay, Peter the Great Gulf, and the Sea of Japan (42.7090° N, 133.1809° E) (Figure 5). The species were kindly identified by Dr. Alexey V. Chernyshev from the Far Eastern Branch of the Russian Academy of Sciences, A.V. Zhirmunsky National Scientific Center of Marine Biology (Vladivostok, Russia). Before RNA isolation, animals were kept in aerated aquaria at 17 °C.
5.2. RNA Isolation, Library Preparation, and Sequencing
The total RNA of Ce. cf. simula middle of body was isolated using TRIzol Reagent (Thermo Fisher Scientific, Waltham, MA, USA), according to the manufacturer’s protocol. RNA concentration and quality were assessed using a BioSpec-nano analyzer (Shimadzu, Kyoto, Japan) at 260/280 nm and 260/230 nm. The length of the fragments was estimated by electrophoresis in 1.2% agarose gel in TAE buffer stained with ethidium bromide with an RNA length marker, i.e., the RiboRuler High Range RNA Ladder (Thermo Fisher Scientific). mRNA was isolated from total RNA using the NEBNext Poly(A) mRNA Magnetic Isolation Module kit (New England Biolabs, Ipswich, MA, USA), followed by double-stranded cDNA synthesis using the Mint2 kit (Eurogen, Moscow, Russia). The result was evaluated by electrophoresis in 1.2% agarose gel in TAE buffer stained with ethidium bromide using the 1 kb DNA ladder DNA length marker (Thermo Fisher Scientific). The double-stranded cDNA was isolated from the reaction mixture with the Bioline ISOLATE II PCR and Gel Kit (Meridian Bioscience Inc., Cincinnati, OH, USA), following the manufacturer’s protocol. The concentration of isolated cDNA was assessed using a Qubit fluorometer (Thermo Fisher Scientific). For sample enrichment with low-represented sequences, cDNA was normalized using the Trimmer-2 kit (Evrogen).
Normalized and double-stranded cDNA sequencing was performed on a MinION Mk1B Oxford Nanopore platform (Oxford Nanopore Technologies, Oxford, UK) using the Direct cDNA Sequencing Kit SQK-DCS109 (Oxford Nanopore Technologies), following the manufacturer’s protocol.
To prepare the cDNA library for the Illumina platform (San Diego, CA, USA), normalized and non-normalized cDNA samples were amplified. The library for Illumina was prepared by the NEBNext Ultra II FS DNA Library Prep Kit for Illumina (New England Biolabs), using the protocol for >100 ng cDNA samples. Sequencing was outsourced to JSC “TsGRM “GENETIKO” (Moscow, Russia).
5.3. Transcriptome Assembly and Data Analysis
To assemble the nemertean transcriptomes, we used datasets from the SRA (https://www.ncbi.nlm.nih.gov/sra, accessed on 18 January 2022) that met two requirements: (1) the raw reads contained more than just 3Gbases and (2) after the raw reads were decontaminated, the GC count per read was close to the theoretical distribution, i.e., Am. lactifloreus (SRR11906528), M. grossa (SRR1507002), P. peregrina (SRR1611562), Ca. hamanako (SRR1505094), Ce. hongkongiensis (SRR618505), T. polymorphus (SRR1611583), B. unicolor (SRR1505175), H. ijimai (SRR1505100), L. longissimus (SRR2682192), L. ruber (SRR1324988), L. sanguineus (SRR3581110), and R. occultus (SRR1505179). The read quality was assessed using the FastQC v0.11.9 software package (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/, accessed on 23 January 2022). Adapter removal and quality filtering (Q ≥ 20) were performed using Trimmomatic v0.39 [47]. De novo transcriptomes were assembled using SPAdes v.3.15.3 [48]. The transcriptome assembly was assessed using BUSCO v5.2.1. [49]. The assembled N. geniculatus transcriptome [17] was downloaded from the Marine Genomics Unit (https://marinegenomics.oist.jp/, accessed on 18 January 2022) (Okinawa Institute of Science and Technology).
The Ce. cf. simula transcriptome assembly was carried out using a pipeline, developed in the current study, based on a hybrid method combining data obtained on the MinION Oxford Nanopore and Illumina platforms. The pipeline included the following steps. First, adapters and chimeric sequences were removed from Oxford Nanopore long reads using Porechop (v. 0.2.4) (https://github.com/rrwick/Porechop, accessed on 23 January 2022). Then, Illumina short reads were corrected using prepared Oxford Nanopore reads by minimap2 (v. 2.24-r1122) (ten cycles) [50] and racon (v. 1.4.13) [51]. Unigenes were obtained using the CAP3 program [52]. Illumina short reads were used separately for de novo transcriptome assembly in SPAdes v.3.15.3. Then, assembled from Illumina short reads, the transcriptome was combined with the corrected Oxford Nanopore long reads using minimap2 (v. 2.24-r1122) and racon (v. 1.4.13).
After assembly, all nemertean transcriptomes were decontaminated using SortMeRNA (v.2.1b) [53] and Seal (from the BBMap v.38.95 package) (https://sourceforge.net/projects/bbmap/, accessed on 23 January 2022) from the noncoding RNA and mRNA of viruses, bacteria, fungi, and humans.
For open reading frame determination (>70 amino acids) and the prediction of protein sequences, TransDecoder (v. 5.5.0) (https://github.com/TransDecoder/TransDecoder, accessed on 24 January 2022) was used. Sequence annotation was carried out through the BLASTP search [54] in the SWISS-PROT database (metazoa, 193,521 sequences) (The UniProt Consortium, 2021), as well as protein domain families (HMMER) [55] in the Pfam database (v. 35.0, 19,632 entries), and open reading frames with the highest homology with known sequences (e ≤ 10–6) were selected. The toxin search (BLASTP) was performed against the Tox-Prot database (7343 sequences) [56]. For further analysis, proteins with identical annotation in the SWISS-PROT/Tox-Prot databases were taken. Transcripts expression in transcripts per million (TPM) was measured using Salmon (v.1.7.0) [57]. Mature proteins sequences were determined using SignalP (v. 6.0) [58].
PCoA was performed using the QIIME 2 software package [59]. Initially, a matrix was formed, in which 86 families of toxins were correlated (0: absence; 1: presence) with the studied nemertean species. Based on this matrix, the Jaccard distance was calculated and the graph of beta diversity was plotted.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/toxins14080542/s1, Supplementary File S1. Nemertean putative toxin transcripts: Table S1: Toxin transcripts of Carinoma hamanako, Table S2: Toxin transcripts of Cephalothrix hongkongiensis, Table S3: Toxin transcripts of Cephalothrix cf. simula, Table S4: Toxin transcripts of Tubulanus polymorphus, Table S5: Toxin transcripts of Baseodiscus unicolor, Table S6: Toxin transcripts of Hubrechtella ijimai, Table S7: Toxin transcripts of Lineus longissimus, Table S8: Toxin transcripts of Lineus ruber, Table S9: Toxin transcripts of Lineus sanguineus, Table S10: Toxins transcripts of Notospermus geniculatus, Table S11: Toxin transcripts of Riseriellus occultus, Table S12: Toxin transcripts of Amphiporous lactifloreus, Table S13: Toxin transcripts of Malacobdella grossa, Table S14: Toxin transcripts of Paranemertes peregrina, Supplementary File S2: Sequences of nemertean toxins; Supplementary File S3: Alignments of putative toxins protein sequences.
Author Contributions
Conceptualization, T.Y.M. and A.E.V.; investigation, T.Y.M., A.E.V. and V.G.K.; data curation, V.G.K.; writing—original draft preparation, A.E.V.; writing—review and editing, T.Y.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Sequence files and metadata for all samples used in this study have been deposited in the NCBI Sequence Read Archive repository under BioProject PRJNA832799 (accession numbers SRR18968559, SRR18959724, SRR18957873, SRR19090815), https://www.ncbi.nlm.nih.gov/bioproject/PRJNA832799, accessed on 27 May 2022.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chen, N.; Xu, S.; Zhang, Y.; Wang, F. Animal protein toxins: Origins and therapeutic applications. Biophys. Rep. 2018, 4, 233–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordon, K.d.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T.; Riedl, R. Paracelsus’ legacy in the faunal realm: Drugs deriving from animal toxins. Drug Discov. Today 2022, 27, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Xie, B.; Huang, Y.; Baumann, K.; Fry, B.G.; Shi, Q. From Marine Venoms to Drugs: Efficiently Supported by a Combination of Transcriptomics and Proteomics. Mar. Drugs 2017, 15, 103. [Google Scholar] [CrossRef]
- Nagai, H. Marine Protein Toxins. In Handbook of Marine Natural Products; Fattorusso, E., Gerwick, W.H., Taglialatela-Scafati, O., Eds.; Springer: New York, NY, USA, 2012; pp. 1–32. [Google Scholar]
- Brahma, R.K.; McCleary, R.J.R.; Kini, R.M.; Doley, R. Venom gland transcriptomics for identifying, cataloging, and characterizing venom proteins in snakes. Toxicon 2015, 93, 1–10. [Google Scholar] [CrossRef]
- Petricevich, V.L. Scorpion Venom and the Inflammatory Response. Mediat. Inflamm. 2010, 2010, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Langenegger, N.; Nentwig, W.; Kuhn-Nentwig, L. Spider Venom: Components, Modes of Action, and Novel Strategies in Transcriptomic and Proteomic Analyses. Toxins 2019, 11, 611. [Google Scholar] [CrossRef] [Green Version]
- Madio, B.; King, G.F.; Undheim, E.A.B. Sea Anemone Toxins: A Structural Overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef] [Green Version]
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient Venom Systems: A Review on Cnidaria Toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef] [Green Version]
- Von Reumont, B.M. Studying Smaller and Neglected Organisms in Modern Evolutionary Venomics Implementing RNASeq (Transcriptomics)—A Critical Guide. Toxins 2018, 10, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajihara, H.; Chernyshev, A.V.; Sun, S.-C.; Sundberg, P.; Crandall, F.B. Checklist of Nemertean Genera and Species Published between 1995 and 2007. Species Divers. 2008, 13, 245–274. [Google Scholar] [CrossRef] [Green Version]
- Chernyshev, A.V. An updated classification of the phylum Nemertea. Invertebr. Zool. 2021, 18, 188–196. [Google Scholar] [CrossRef]
- Göransson, U.; Jacobsson, E.; Strand, M.; Andersson, H.S. The Toxins of Nemertean Worms. Toxins 2019, 11, 120. [Google Scholar] [CrossRef] [Green Version]
- Whelan, N.V.; Kocot, K.M.; Santos, S.R.; Halanych, K.M. Nemertean Toxin Genes Revealed through Transcriptome Sequencing. Genome Biol. Evol. 2014, 6, 3314–3325. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.-J.; Kanda, M.; Koyanagi, R.; Hisata, K.; Akiyama, T.; Sakamoto, H.; Sakamoto, T.; Satoh, N. Nemertean and phoronid genomes reveal lophotrochozoan evolution and the origin of bilaterian heads. Nat. Ecol. Evol. 2018, 2, 141–151. [Google Scholar] [CrossRef]
- Von Reumont, B.M.; Lüddecke, T.; Timm, T.; Lochnit, G.; Vilcinskas, A.; Von Döhren, J.; Nilsson, M.A. Proteo-Transcriptomic Analysis Identifies Potential Novel Toxins Secreted by the Predatory, Prey-Piercing Ribbon Worm Amphiporus lactifloreus. Mar. Drugs 2020, 18, 407. [Google Scholar] [CrossRef]
- Verdes, A.; Taboada, S.; Hamilton, B.R.; Undheim, E.A.B. Evolution, Expression Patterns, and Distribution of Novel Ribbon Worm Predatory and Defensive Toxins. Mol. Biol. Evol. 2022, 39, 1–33. [Google Scholar] [CrossRef]
- Ono, H.; Ishii, K.; Kozaki, T.; Ogiwara, I.; Kanekatsu, M.; Yamada, T. Removal of redundant contigs from de novo RNA-Seq assemblies via homology search improves accurate detection of differentially expressed genes. BMC Genom. 2015, 16, 1031. [Google Scholar] [CrossRef] [Green Version]
- Andrade, S.C.S.; Montenegro, H.; Strand, M.; Schwartz, M.L.; Kajihara, H.; Norenburg, J.L.; Turbeville, J.M.; Sundberg, P.; Giribet, G. A Transcriptomic Approach to Ribbon Worm Systematics (Nemertea): Resolving the Pilidiophora Problem. Mol. Biol. Evol. 2014, 31, 3206–3215. [Google Scholar] [CrossRef] [Green Version]
- Kvist, S.; Chernyshev, A.V.; Giribet, G. Phylogeny of Nemertea with special interest in the placement of diversity from Far East Russia and northeast Asia. Hydrobiologia 2015, 760, 105–119. [Google Scholar] [CrossRef]
- Chernyshev, A.V.; Polyakova, N.E. Nemerteans from the deep-sea expedition KuramBio II with descriptions of three new hoplonemerteans from the Kuril-Kamchatka Trench. Prog. Oceanogr. 2019, 178, 102148. [Google Scholar] [CrossRef]
- Jungo, F.; Bairoch, A. Tox-Prot, the toxin protein annotation program of the Swiss-Prot protein knowledgebase. Toxicon 2005, 45, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Kem, W.R.; Blumenthal, K.M. Purification and characterization of the cytotoxic Cerebratulus A toxins. J. Biol. Chem. 1978, 253, 5752–5757. [Google Scholar] [CrossRef]
- Jacobsson, E.; Andersson, H.S.; Strand, M.; Peigneur, S.; Eriksson, C.; Lodén, H.; Shariatgorji, M.; Andrén, P.E.; Lebbe, E.K.M.; Rosengren, K.J.; et al. Peptide ion channel toxins from the bootlace worm, the longest animal on Earth. Sci. Rep. 2018, 8, 4596. [Google Scholar] [CrossRef]
- Butala, M.; Šega, D.; Tomc, B.; Podlesek, Z.; Kem, W.R.; Küpper, F.C.; Turk, T. Recombinant expression and predicted structure of parborlysin, a cytolytic protein from the Antarctic heteronemertine Parborlasia corrugatus. Toxicon 2015, 108, 32–37. [Google Scholar] [CrossRef]
- Kem, W.R. Structure and Action of Nemertine Toxins. Integr. Comp. Biol. 1985, 25, 99–111. [Google Scholar] [CrossRef]
- Ali, A.E.; Arakawa, O.; Noguchi, T.; Miyazawa, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin and related substances in a ribbon worm Cephalothrix linearis (Nemertean). Toxicon 1990, 28, 1083–1093. [Google Scholar] [CrossRef]
- Asakawa, M.; Ito, K.; Kajihara, H. Highly Toxic Ribbon Worm Cephalothrix simula Containing Tetrodotoxin in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxins 2013, 5, 376–395. [Google Scholar] [CrossRef]
- Vlasenko, A.E.; Magarlamov, T.Y. Tetrodotoxin and its analogues in Cephalothrix cf. simula (Nemertea: Palaeonemertea) from the Sea of Japan (Peter the Great Gulf): Intrabody distribution and secretions. Toxins 2020, 12, 745. [Google Scholar]
- Vlasenko, A.E.; Velansky, P.V.; Chernyshev, A.V.; Kuznetsov, V.G.; Magarlamov, T.Y. Tetrodotoxin and its analogues profile in nemertean species from the sea of Japan. Toxicon 2018, 156, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Fenwick, D.; Powell, A.; Dhanji-Rapkova, M.; Ford, C.; Hatfield, R.G.; Santos, A.; Martinez-Urtaza, J.; Bean, T.P.; Baker-Austin, C.; et al. New Invasive Nemertean Species (Cephalothrix Simula) in England with High Levels of Tetrodotoxin and a Microbiome Linked to Toxin Metabolism. Mar. Drugs 2018, 16, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kem, W.R. Structure and membrane actions of a marine worm protein cytolysin, Cerebratulus toxin A-III. Toxicology 1994, 87, 189–203. [Google Scholar] [CrossRef]
- Arbuckle, K. Evolutionary context of venom in animals. In Evolution of Venomous Animals and Their Toxins. Toxinology; Gopalakrishnakone, P., Malhotra, A., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 3–31. [Google Scholar]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Jennings, J.B.; Gibson, R. Observations on the Nutrition of Seven Species of Rhynchocoelan Worms. Biol. Bull. 1969, 136, 405–433. [Google Scholar] [CrossRef]
- McDermott, J.J.; Roe, P. Food, Feeding Behavior and Feeding Ecology of Nemerteans. Am. Zool. 1985, 25, 113–125. [Google Scholar] [CrossRef]
- Stricker, S.A.; Cloney, R.A. The ultarstructure of venom-producing cells in Paranemertes peregrina (Nemertea, Hoplonemertea). J. Morphol. 1983, 177, 89–107. [Google Scholar] [CrossRef]
- Thiel, M.; Cruse, I. Status of the Nemertea as predators in marine ecosystems. Hydrobiologia 2001, 456, 21–32. [Google Scholar] [CrossRef]
- Bourque, D.; Miron, G.; Landry, T. Predation on soft-shell clams (Mya arenaria) by the nemertean Cerebratulus lacteus in Atlantic Canada: Implications for control measures. Hydrobiologia 2001, 456, 33–44. [Google Scholar] [CrossRef]
- Beckers, P.; Bartolomaeus, T.; von Döhren, J. Observations and Experiments on the Biology and Life History of Riseriellus occultus (Heteronemertea: Lineidae). Zool. Sci. 2015, 32, 531–546. [Google Scholar] [CrossRef]
- Chernyshev, A.V. Food and feeding behavior of the nemerteanTortus tokmakovae. Russ. J. Mar. Biol. 2000, 26, 120–123. [Google Scholar] [CrossRef]
- Bürger, O. Die Nemertinen des Golfes von Neapel und der Angrenzenden Meeres–Abschnitte, Fauna und Flora des Golfes von Neapel; Verlag von R. Friedländer & Sohn: Berlin, Germany, 1895; Volume 22. [Google Scholar]
- Bürger, O. Nemertini (Schnurwürmer). In Klassen und Ordnungen des Tier-Reichs; C.F. Winter’sche Verlagshandlung: Leipzig, Germany, 1897; Volume 4, p. 542. [Google Scholar]
- Thiel, M. Nemertines as predators on tidal flats—High Noon at low tide. Hydrobiologia 1997, 365, 241–250. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes De Novo Assembler. Curr. Protoc. Bioinform. 2020, 70, 1–29. [Google Scholar] [CrossRef]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.A.; Zdobnov, E.M. BUSCO Update: Novel and Streamlined Workflows along with Broader and Deeper Phylogenetic Coverage for Scoring of Eukaryotic, Prokaryotic, and Viral Genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed]
- Vaser, R.; Sović, I.; Nagarajan, N.; Šikić, M. Fast and accurate de novo genome assembly from long uncorrected reads. Genome Res. 2017, 27, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Madan, A. CAP3: A DNA Sequence Assembly Program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef] [Green Version]
- Kopylova, E.; Noé, L.; Touzet, H. SortMeRNA: Fast and accurate filtering of ribosomal RNAs in metatranscriptomic data. Bioinformatics 2012, 28, 3211–3217. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.S.; Eddy, S.R.; Portugaly, E. Hidden Markov model speed heuristic and iterative HMM search procedure. BMC Bioinform. 2010, 11, 431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungo, F.; Bougueleret, L.; Xenarios, I.; Poux, S. The UniProtKB/Swiss-Prot Tox-Prot program: A central hub of integrated venom protein data. Toxicon 2012, 60, 551–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Nemertean phylogenetic tree (phylogeny modified from [21,22,23]). The black figures indicate the number of toxin families, and the red figures indicate the number of toxin transcripts.

Figure 2.
Proportional distributions of toxin families’ transcripts in nemertean transcriptomes. (a) Relative distribution, (b) absolute numbers, (c) the color map for both (a,b) charts.
Figure 2.
Proportional distributions of toxin families’ transcripts in nemertean transcriptomes. (a) Relative distribution, (b) absolute numbers, (c) the color map for both (a,b) charts.

Figure 3.
Venn diagram representing the distribution of the numbers of toxin families identified in the three nemertean classes.
Figure 3.
Venn diagram representing the distribution of the numbers of toxin families identified in the three nemertean classes.

Figure 4.
Principal coordinate analysis (PCoA) of the nemertean species based on the presence/absence of the toxin families using Jaccard distance (pseudo−F = 3.099, p = 0.0001, PERMANOVA, 10,000 permutations in each test).
Figure 4.
Principal coordinate analysis (PCoA) of the nemertean species based on the presence/absence of the toxin families using Jaccard distance (pseudo−F = 3.099, p = 0.0001, PERMANOVA, 10,000 permutations in each test).

Figure 5.
(a) A specimen of Cephalothrix cf. simula, (b) location of Ce. cf. simula collection.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Assembly statistics of nemertean transcriptomes.
| Species | NCBI Run Accession Numbers | Sample Description | Seq Platform | Raw Gigabases | Raw Reads | Size (Gb) | Number of Q20 Reads | Number of Contigs | Number of Decontaminated Contigs | Unigenes | Mean Contig Length | Contig N50 | BUSCO, % | Number of ORFs | ORFs with BLAST Hits | ORFs with Unique BLAST Hits |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Palaeonemertea | ||||||||||||||||
| Carinoma hamanako | SRR1505094 | Whole individual | Illumina Genome Analyzer II | 3.8 | 52,121,008 | 2.0 | 30,851,767 | 74,088 | 71,537 | 67,315 | 604.7 | 1206 | 81.2 | 25,169 | 15,209 | 11,453 |
| Cephalothrix hongkongiensis | SRR618505 | Obtained from three individuals | Illumina Genome Analyzer II | 5.2 | 52,224,518 | 2.2 | 23,706,369 | 101,598 | 99,702 | 94,375 | 451.6 | 722 | 78.3 | 28,675 | 15,979 | 11,841 |
| Cephalothrix cf. simula | SRR18959724 SRR18957873 | Middle of body | Illumina NovaSeq6000 | 16.1 | 166,114,952 | 5.2 | 166,114,952 | 183,960 | 125,779 | 498,451 | 1132.8 | 1501 | 93.1 | 208,177 | 79,802 | 25,895 |
| Cephalothrix cf. simula | SRR19090815 SRR18968559 | Middle of body | MinION Oxford Nanopore | 8.6 | 7,649,450 | 7.1 | – | 4,150,437 | ||||||||
| Tubulanus polymorphus | SRR1611583 | Anterior end of body | Illumina HiSeq 2000 | 3.9 | 39,262,732 | 2.6 | 12,802,492 | 83,720 | 81,000 | 77,268 | 609.6 | 1307 | 82.9 | 26,069 | 14,725 | 11,404 |
| Pilidiophora | ||||||||||||||||
| Baseodiscus unicolor | SRR1505175 | Part of one sample | Illumina Genome Analyzer II | 7.1 | 78,906,444 | 3.5 | 78,906,444 | 272,007 | 267,551 | 257,404 | 637.2 | 1575 | 77.6 | 56,878 | 18,067 | 12,335 |
| Hubrechtella ijimai | SRR1505100 | Whole individual | Illumina Genome Analyzer II | 4.8 | 49,505,380 | 2.5 | 22,944,699 | 226,722 | 221,044 | 200,648 | 420.1 | 790 | 64.7 | 36,772 | 16,640 | 11,503 |
| Lineus longissimus | SRR2682192 | Mix of embryonic and juveniles | Illumina HiSeq 2000 | 43.5 | 435,278,178 | 25.4 | 415,964,152 | 184,216 | 172,720 | 155,885 | 894.2 | 3132 | 93.7 | 35,740 | 17,909 | 12,004 |
| Lineus ruber | SRR1324988 | Whole body | Illumina HiSeq 2000 | 5.3 | 52,196,821 | 3.3 | 49,280,205 | 187,137 | 184,858 | 161,352 | 558.7 | 1016 | 80.6 | 40,754 | 19,042 | 12,826 |
| Lineus sanguineus | SRR3581110 | Whole organism | Illumina HiSeq 2000 | 11.7 | 122,617,800 | 5.8 | 115,307,676 | 198,867 | 195,903 | 179,731 | 470.4 | 919 | 79.2 | 46,025 | 22,599 | 14,748 |
| Notospermus geniculatus | Adult tissues and embryonic stages | Assembly | 96,304 | 94,070 | 77,565 | 1286.6 | 1955 | 92.2 | 58,991 | 24,541 | 14,520 | |||||
| Riseriellus occultus | SRR1505179 | Part of one sample | Illumina Genome Analyzer II | 3.9 | 47,787,302 | 2.0 | 33,542,144 | 133,713 | 131,395 | 117,212 | 704.1 | 1531 | 66.4 | 35,025 | 17,445 | 12,462 |
| Hoplonemertea | ||||||||||||||||
| Amphiporous lactifloreus | SRR11906528 | Whole proboscis | Illumina HiSeq 2500 | 7.0 | 46,559,252 | 2.2 | 39,466,994 | 55,758 | 50,950 | 46,014 | 1128.8 | 2464 | 87.0 | 23,027 | 12,214 | 8928 |
| Malacobdella grossa | SRR1611560 | 1 whole animal | Illumina HiSeq 2000 | 3.1 | 30,538,858 | 2.0 | 17,132,560 | 47,700 | 45,334 | 42,351 | 791.0 | 1905 | 84.4 | 15,712 | 9768 | 8030 |
| Paranemertes peregrina | SRR1611562 | Anterior ~1/3 of 1 individual | Illumina HiSeq 2000 | 5.9 | 59,441,992 | 3.6 | 26,918,767 | 75,226 | 69,774 | 66,797 | 533.5 | 1131 | 63.9 | 19,744 | 10,666 | 8282 |
–: not applicable.
Table 2.
Number of toxin families identified in the three nemertean classes.
| Nemertean Classes | Neurotoxins | Pore-Forming Toxins | Enzymes | Proteinase Inhibitors | Ion Channel Inhibitors | Hemostasis-Impairing Toxins | Other Toxin Candidates |
|---|---|---|---|---|---|---|---|
| Common for all | |||||||
| Palaeonemertea/Pilidiophora/Hoplonemertea | 1 | 3 | 2 | 3 | 3 | 3 | |
| Common for pairs | |||||||
| Palaeonemertea/Pilidiphora | 1 | 11 | 1 | 2 | 7 | ||
| Palaeonemertea/Hoplonemertea | 1 | ||||||
| Hoplonemertea/Pilidiphora | 1 | 1 | 3 | ||||
| Unique | |||||||
| Palaeonemertea | 1 | 1 | 5 | 1 | 2 | 1 | 2 |
| Pilidiphora | 4 | 10 | 4 | 2 | 7 | ||
| Hoplonemertea | 2 | 1 | 1 | ||||
Table 3.
Relative abundance of toxin groups’ transcripts identified in three nemertean classes.
| Toxin Groups | Hoplonemertea | Palaeonemertea | Pilidiophora |
|---|---|---|---|
| Neurotoxins | NA | +++ | +++ |
| Pore-forming toxins | +++ | ++ | +++ |
| Enzymes | + | + | + |
| Proteinase inhibitors | +++ | + | +++ |
| Ion channel inhibitors | + | ++ | ++ |
| Hemostasis-impairing toxins | ++ | ++ | + |
| Other toxins candidates | +++ | ++ | + |
NA: not available; +: 0–9.99% of the total sample expression; ++: 10–19.99% of the total sample expression; +++: ≥20% of the total sample expression.
Table 4.
Relative abundance of toxins that together make up 50% of the total toxin expression (major toxins) in nemerteans.
Table 4.
Relative abundance of toxins that together make up 50% of the total toxin expression (major toxins) in nemerteans.
| Protein Family | Tox-Prot Annotation | Expression, % | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Palaeonemertea | Pilidiophora | Hoplonemertea | |||||||||||||
| Carinoma hamanako | Cephalothrix hongkongiensis | Cephalothrix cf. simula | Tubulanus polymorphus | Baseodiscus unicolor | Hubrechtella ijimai | Lineus longissimus | Lineus ruber | Lineus sanguineus | Notosermus geniculatus | Riseriellus occultus | Amphiporous lactifloreus | Malacobdella grossa | Paranemertes peregrina | ||
| Neurotoxins | |||||||||||||||
| Neurotoxin 20 family. | U3-aranetoxin-Ce1a | 30.3 | |||||||||||||
| Neurotoxin 02 (plectoxin) family. 02 (plectoxin) subfamily | Omega-plectoxin-Pt1a | 35.3 | |||||||||||||
| Neurotoxin 10 (Hwtx-1) family. 15 (Hntx-3) subfamily | Hainantoxin-III 8 | 21.9 | |||||||||||||
| Pore-forming toxins | |||||||||||||||
| MACPF domain | Perivitellin-2 67 kDa subunit | 33.2 | |||||||||||||
| DELTA-alicitoxin-Pse2a | 17.2 | 14.7 | |||||||||||||
| Worm cytolysin family | Cytotoxin A-III | 16.0 | 33.8 | ||||||||||||
| Enzymes | |||||||||||||||
| Phospholipase A2 family. Group III subfamily | Phospholipase A2 isozymes PA3A/PA3B/PA5 | 6.0 | 21.9 | ||||||||||||
| Arthropod phospholipase D family. Class II subfamily. Class IIb sub-subfamily | Dermonecrotic toxin LiSicTox-alphaV1 | 16.8 | 23.9 | ||||||||||||
| Dermonecrotic toxin LhSicTox-alphaIA1ii | 17.9 | ||||||||||||||
| Proteinase inhibitors | |||||||||||||||
| Venom Kunitz-type family. Sea Anemone type 2 potassium channel toxin subfamily | PI-actitoxin-Axm2b | 56.3 | |||||||||||||
| Actinia tenebrosa protease inhibitors | 21.6 | 18.9 | |||||||||||||
| Venom Kunitz-type family | Kunitz-type serine protease inhibitor conotoxin Cal9.1a | 14.5 | 40.2 | ||||||||||||
| Kunitz-type serine protease inhibitor conotoxin Cal9.1d | 5.0 | ||||||||||||||
| Kunitz-type serine protease inhibitor microlepidin-1 | 23.6 | ||||||||||||||
| Kunitz-type serine protease inhibitor Bt-KTI | 18.7 | ||||||||||||||
| Natriuretic peptide family | Snake venom metalloprotease inhibitor 02A10 | 28.9 | |||||||||||||
| Ion channel inhibitors | |||||||||||||||
| Sea anemone structural class 9a family | Delta-actitoxin-Amc1a | 27.8 | |||||||||||||
| CRISP family | CRISP/Allergen/PR-1 | 30.4 | |||||||||||||
| Cysteine-rich venom protein Mr30 | 12.6 | ||||||||||||||
| Cysteine-rich venom protein pseudechetoxin-like | 17.9 | ||||||||||||||
| Cysteine-rich venom protein | 18.9 | ||||||||||||||
| CRISP family. Venom allergen 5-like subfamily | Scoloptoxin SSD976 | 9.3 | 8.2 | ||||||||||||
| Venom Kunitz-type family. Sea anemone type 2 potassium channel toxin subfamily | U-actitoxin-Avd3h | 35.6 | |||||||||||||
| U-actitoxin-Avd3l | 6.7 | ||||||||||||||
| Hemostasis-impairing toxins | |||||||||||||||
| Snaclec family | Snaclec 3 | 15.9 | |||||||||||||
| Other toxins candidates | |||||||||||||||
| TCTP family | Translationally controlled tumor protein homolog OS=Branchiostoma belcheri | 47.8 | |||||||||||||
| Translationally controlled tumor protein homolog OS=Brassica oleracea | 21.3 | ||||||||||||||
| Insulin-like growth factor-binding protein-related protein 1 | Insulin-like growth factor-binding protein-related protein 1 | 12.5 | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vlasenko, A.E.; Kuznetsov, V.G.; Magarlamov, T.Y. Investigation of Peptide Toxin Diversity in Ribbon Worms (Nemertea) Using a Transcriptomic Approach. Toxins 2022, 14, 542. https://doi.org/10.3390/toxins14080542
AMA Style
Vlasenko AE, Kuznetsov VG, Magarlamov TY. Investigation of Peptide Toxin Diversity in Ribbon Worms (Nemertea) Using a Transcriptomic Approach. Toxins. 2022; 14(8):542. https://doi.org/10.3390/toxins14080542
Chicago/Turabian StyleVlasenko, Anna E., Vasiliy G. Kuznetsov, and Timur Yu. Magarlamov. 2022. "Investigation of Peptide Toxin Diversity in Ribbon Worms (Nemertea) Using a Transcriptomic Approach" Toxins 14, no. 8: 542. https://doi.org/10.3390/toxins14080542
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.