
Spatially Explicit Environmental Factors Associated with Lymphatic Filariasis Infection in American Samoa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location
2.2. Data Collection
2.2.1. Infection Markers
2.2.2. Survey Design
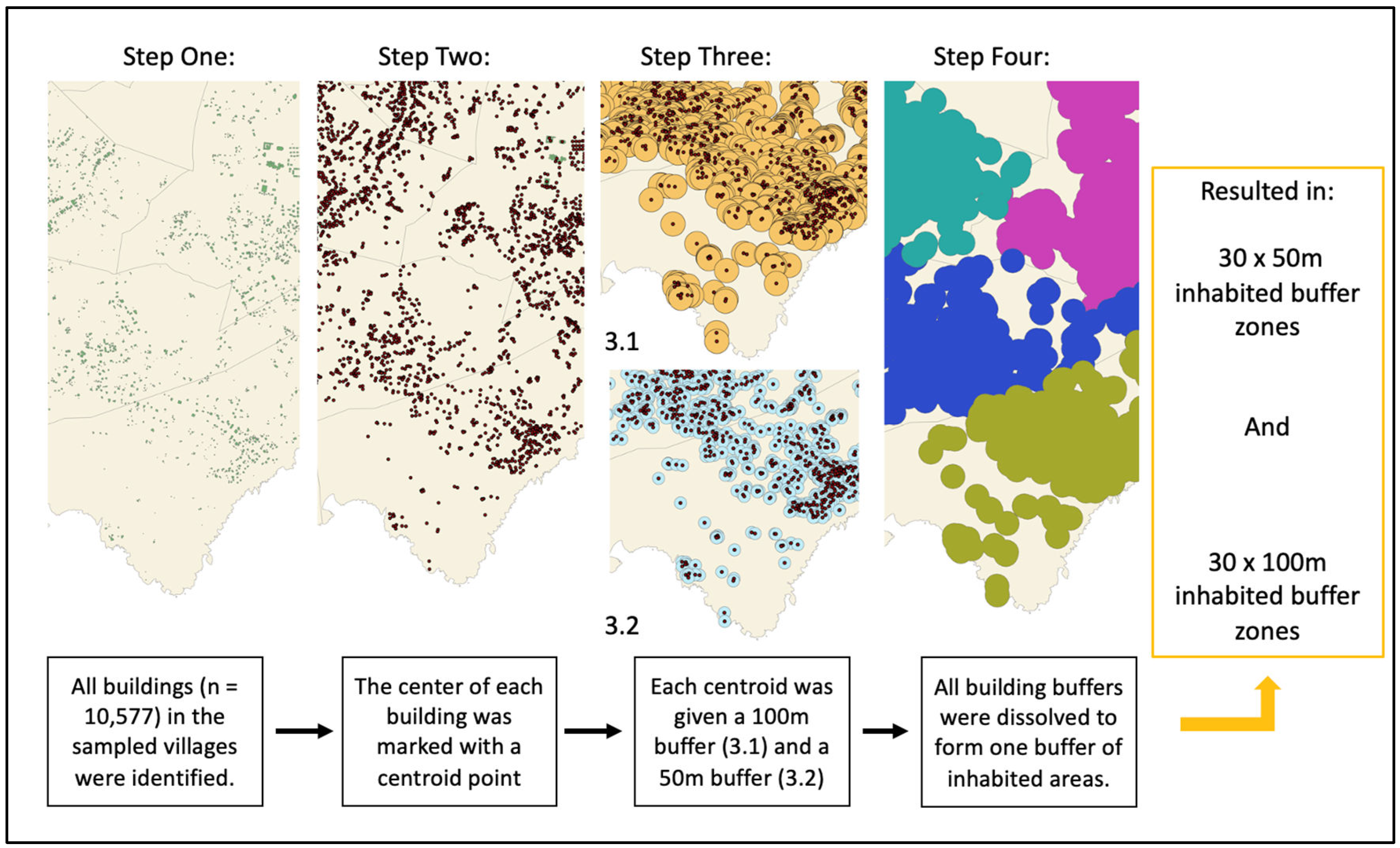
2.2.3. Spatial Layers on Island, Village and Building Boundaries
2.2.4. Environmental Variables and Environmental Data
2.2.5. Environmental Data Collection and Extraction
2.3. Associations between Environmental Variables and LF Infection Markers
3. Results
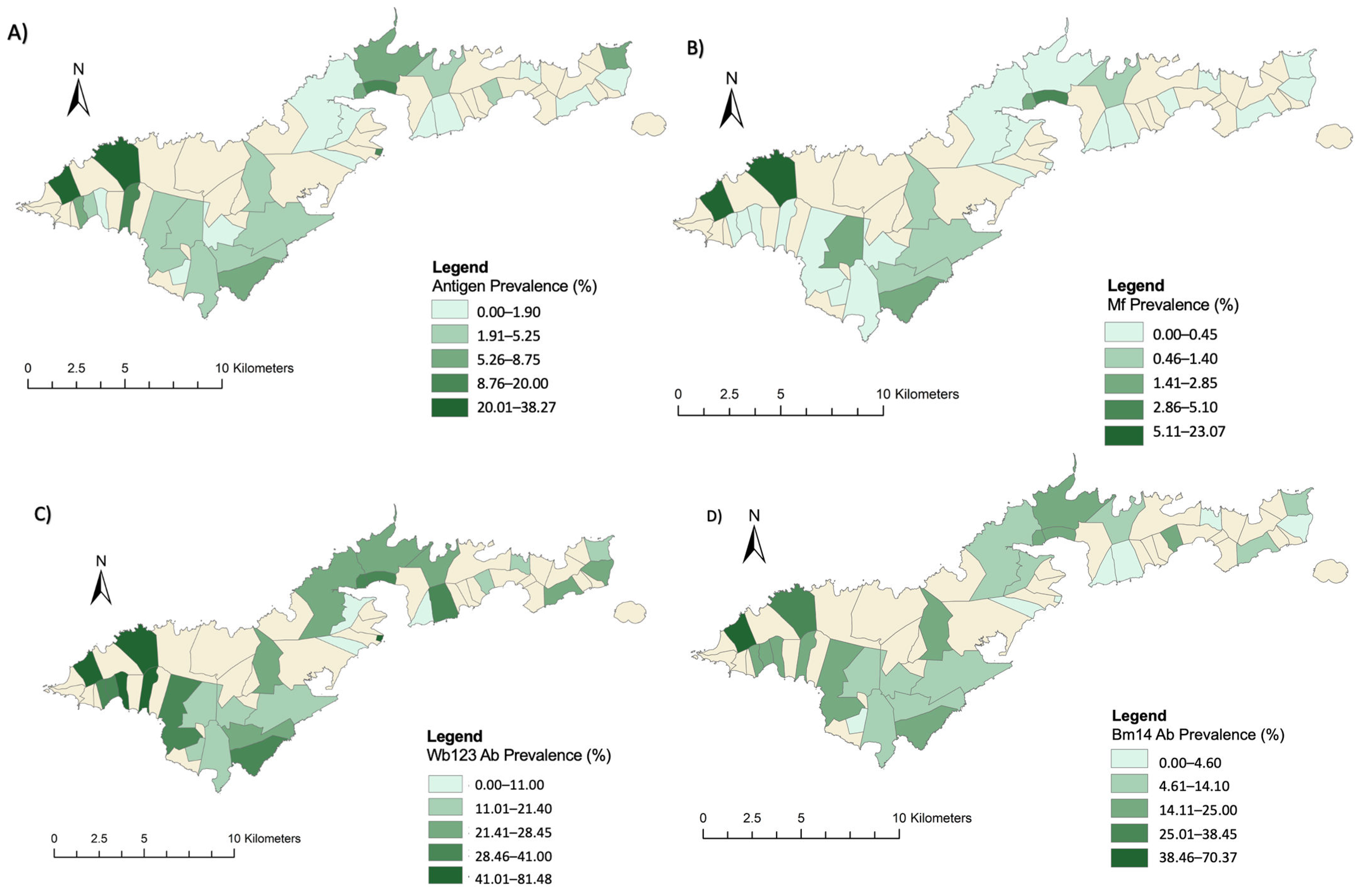
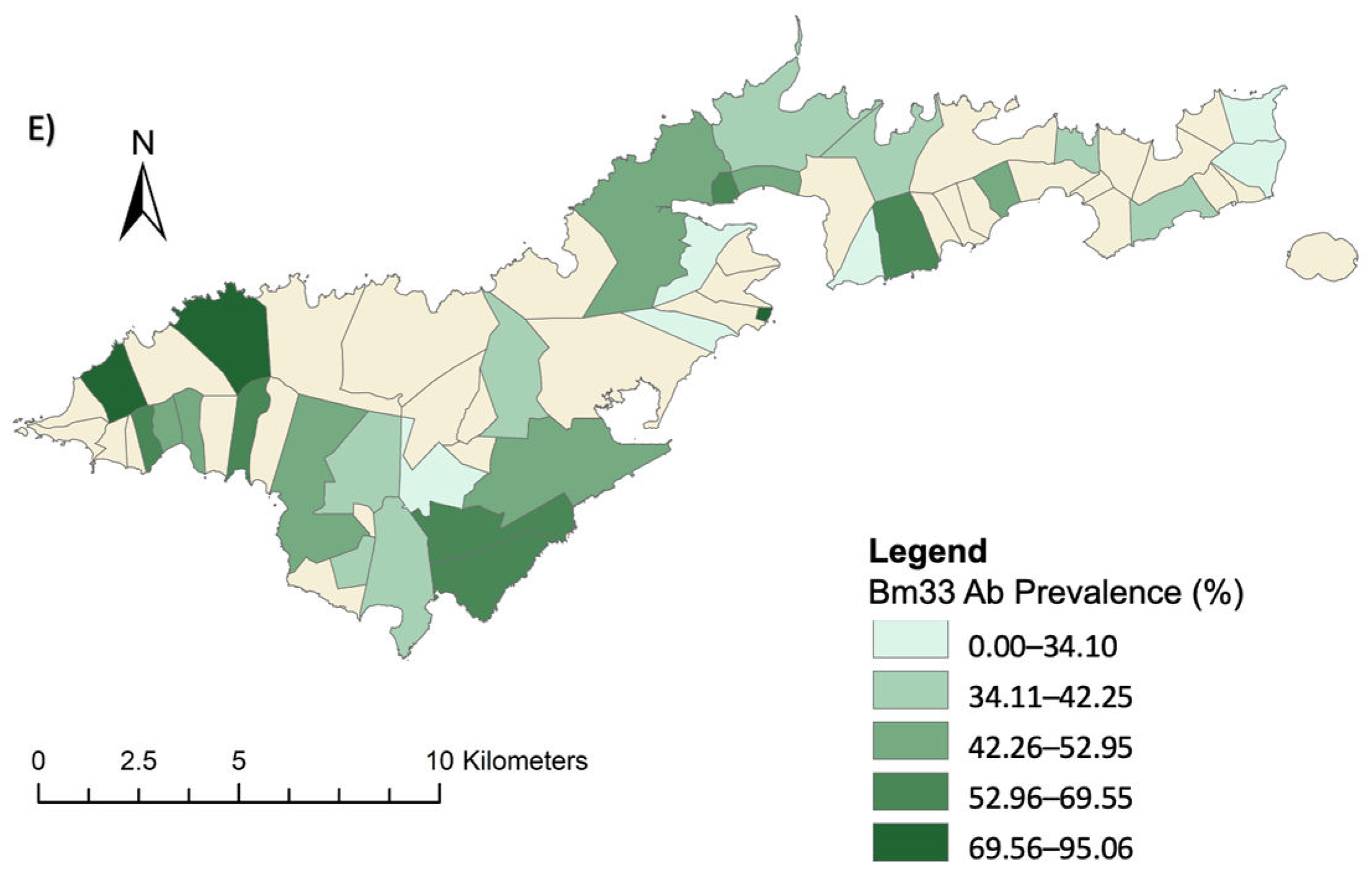
3.1. Village Level Prevalence of Infection Markers
3.2. Village-Level Environmental Data
3.3. Multivariable Poisson Regression Models
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Summary of Studies Investigating Associations between Environmental Variables and Lymphatic Filariasis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | n * | Study Reference | Location | Most Common Relationship Found | |
|---|---|---|---|---|---|
| Temperature | Day Land Surface Temperature | 4 | Mwase et al., 2014 [53] Stensgaard et al., 2011 [2] Eneanya et al., 2018 [32] Kwarteng et al., 2021 [50] | Zambia Uganda Nigeria Ghana | Positive (n = 2) |
| Night Land Surface Temperature | 3 | Mwase et al., 2014 [53] Stensgaard et al., 2011 [2] Kwarteng et al., 2021 [50] | Zambia Uganda Ghana | No relationship was found. | |
| Mean max/min temperature | 2 | Cano et al., 2014 [4] Eneanya et al., 2018 [32] | Global Nigeria | A positive relationship with mean minimum temperature. | |
| Average Annual Temperature | 8 | De Souza et al., 2010 [31] Manhenje et al., 2013 [1] Cano et al., 2014 [4] Stanton et al., 2013 [51] Eneanya et al., 2018 [32] Onapa et al., 2005 [54] Palayandi et al., 2014 [52] Slater & Michael 2013 [55] | Ghana Mozambique Global Burkina Faso Nigeria Uganda India African Continent | Positive, linear (n = 6) | |
| Land | Normalised Difference Vegetation Index (NDVI) | 8 | Mwase et al., 2014 [53] Stensgaard et al., 2011 [2] De Souza et al., 2010 [31] Cano et al., 2014 [4] Stanton et al., 2013 [51] Eneanya et al., 2018 [32] Kwarteng et al., 2021 [50] Slater & Michael 2013 [55] | Zambia Uganda Ghana Global Burkina Faso Nigeria Ghana African Continent | Positive (n = 4) Did not contribute to the model (n = 3) |
| Soil pH | 1 | Eneanya et al., 2018 [32] | Nigeria | No relationship found | |
| Land Cover | 6 | Mwase et al., 2014 [53] Cano et al., 2014 [4] Stanton et al., 2013 [51] Eneanya et al., 2018 [32] Kwarteng et al., 2021 [50] Rwegoshora et al., 2005 [56] | Zambia Global Burkina Faso Nigeria Ghana East Africa | Varied significantly depending on the type of landcover analysed. | |
| Water | Humidity | 2 | De Souza et al., 2010 [31] Palayandi M. 2014 [52] | Ghana India | Positive (n = 1) Negative (n = 1) |
| Potential evapotranspiration | 1 | Eneanya et al., 2018 [32] | Nigeria | Removed due to multicollinearity | |
| Wetness Index | 2 | Eneanya et al., 2018 [32] Grziwotz et al., 2018 (As dew point) [57] | Nigeria French Polynesia | Positive (n = 2) | |
| Distance to waterbody | 8 | Mwase et al., 2014 [53] Stensgaard et al., 2011 [2] Stanton et al., 2013 [51] Eneanya et al., 2018 [32] Nurjazuli & Santjaka 2020 [58] Chesnais et al., 2019 [59] Edirisinghe M. 2017 [3] Kwarteng et al., 2021 [50] | Zambia Uganda Burkina Faso Nigeria Indonesia Democratic Republic of the Congo Sri Lanka Ghana | Positive (n = 4) No relationship (n = 2) | |
| Aridity | 2 | Cano et al., 2014 [4] Eneanya et al., 2018 [32] | Global Nigeria | No relationship (n = 2) | |
| Number of Months with Rainfall | 1 | Stanton et al., 2013 [51] | Burkina Faso | Positive | |
| Mean tidal level | 1 | Grziwotz et al., 2018 [57] | French Polynesia | Positive | |
| Rainfall/Precipitation | 13 | Mwase et al., 2014 [53] Stensgaard et al., 2011 [2] De Souza et al., 2010 [31] Manhenje et al., 2013 [1] Cano et al., 2014 [4] Stanton et al., 2013 [51] Eneanya et al., 2018 [32] Kwarteng et al., 2021 [50] Hussaini et al., 2020 [60] Grziwotz et al., 2018 [57] Onapa et al., 2005 [54] Palayandi M. 2014 [52] Slater & Michael 2013 [55] | Zambia Uganda Ghana Mozambique Global Burkina Faso Nigeria Ghana Nigeria French Polynesia Uganda India African Continent | Positive, nonlinear (n = 7) | |
| Altitude | Elevation | 12 | Mwase et al., 2014 [53] Stensgaard et al., 2011 [2] De Souza et al., 2010 [31] Manhenje et al., 2013 [1] Cano et al., 2014 [4] Stanton et al., 2013 [51] Eneanya et al., 2018 [32] Kwarteng et al., 2021 [50] Onapa et al., 2005 [54] Palayandi. M 2014 [52] Slater & Michael 2013 [55] Dhimal, Ahrens & Kuch 2014 [21] | Zambia Uganda Ghana Mozambique Global Burkina Faso Nigeria Ghana Uganda India African Continent Nepal | Negative, nonlinear (n = 8) |
| Slope | 2 | Eneanya et al., 2018 [32] Kwarteng et al., 2021 [50] | Nigeria Ghana | Negative (n = 2) | |
| Human Factors | House type | 1 | Srividya et al., 2018 [61] | India | The proportion of concrete and tiled, not thatched, houses was higher in hotspots (31.8% and 47.3%) |
| Water in the house | 2 | Nurjazuli & Santjaka 2020 [58] Hussaini et al., 2020 [60] | Indonesia Nigeria | Positive (n = 2) | |
| Distance to stable light | 2 | Eneanya et al., 2018 [32] Kwarteng et al., 2021 [50] | Nigeria Ghana | Negative (n = 2) LF prevalence decreased with increasing distance | |
Appendix B. Extracted Environmental Data for Normalised Difference Vegetation Index (NDVI), Rainfall (mm), Population Density (persons/km2), Elevation (m), Slope Gradient (degrees), and Landcover Class (Percent of Inhabited Buffer Zone covered) in Each Village Inhabited Buffer zone of the 2016 Lymphatic Filariasis Community Survey, American Samoa
| Average Normalised Difference Vegetation Index | Annual Rainfall (mm) | Average Rainfall in Dry Months (mm) | Average Rainfall in Wet Months (mm) | Average Population Density (Persons/km2) | Average Elevation (m) | Average Slope Gradient (Degrees) | Crop Cover in Inhabited Buffer Zone (%) | Forest Cover in Inhabited Buffer Zone (%) | Rangeland Cover in Inhabited Buffer Zone (%) | Urban Cover in Inhabited Buffer Zone (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| VILLAGE | 50 m | 100 m | 50 m | 100 m | 50 m | 100 m | 50 m | 100 m | 50 m | 100 m | 50 m | 100 m | 50 m | 100 m | 50 m | 100 m | 50 m | 100 m | 50 m | 100 m | 50 m | 100 m |
| Afono | 0.4 | 0.4 | 3447.1 | 3422.3 | 245.5 | 243.0 | 352.9 | 350.7 | 22.6 | 19.1 | 18.1 | 20.3 | 10.1 | 10.0 | 0.0 | 0.0 | 51.8 | 61.8 | 1.4 | 3.2 | 16.5 | 9.6 |
| Alao | 0.4 | 0.4 | 2280.8 | 2281.6 | 152.0 | 152.0 | 239.4 | 239.4 | 26.5 | 23.9 | 63.5 | 58.8 | 16.1 | 16.0 | 0.0 | 0.0 | 28.5 | 35.9 | 5.5 | 7.5 | 62.7 | 43.3 |
| Amaua | 0.4 | 0.4 | 3291.9 | 3298.6 | 230.4 | 231.1 | 338.9 | 339.5 | 17.8 | 16.0 | 159.2 | 168.5 | 25.3 | 27.2 | 0.0 | 0.0 | 35.0 | 40.3 | 7.2 | 18.8 | 52.0 | 26.7 |
| Amouli | 0.3 | 0.3 | 2467.6 | 2461.5 | 166.0 | 165.6 | 258.5 | 257.8 | 18.9 | 17.9 | 94.2 | 92.9 | 22.5 | 22.4 | 0.0 | 0.0 | 38.5 | 49.2 | 9.9 | 11.7 | 45.0 | 24.6 |
| Asili | 0.3 | 0.3 | 3877.8 | 3941.9 | 287.6 | 293.2 | 390.5 | 396.5 | 16.0 | 15.3 | 89.5 | 101.7 | 23.4 | 24.4 | 0.0 | 0.0 | 46.3 | 65.0 | 0.0 | 0.0 | 43.6 | 21.1 |
| Aumi | 0.3 | 0.3 | 3755.6 | 3740.4 | 265.9 | 265.0 | 384.2 | 382.6 | 21.5 | 18.9 | 132.4 | 137.0 | 29.0 | 28.5 | 0.0 | 0.0 | 15.1 | 26.4 | 20.3 | 27.0 | 56.2 | 28.4 |
| Fagalii | 0.4 | 0.4 | 2828.0 | 2842.5 | 195.3 | 196.5 | 292.6 | 293.9 | 12.0 | 10.8 | 8.7 | 9.2 | 4.5 | 5.7 | 0.0 | 0.0 | 66.6 | 78.3 | 0.0 | 0.0 | 31.8 | 17.7 |
| Fagamalo | 0.4 | 0.4 | 2985.0 | 2990.1 | 206.0 | 206.5 | 308.4 | 308.9 | 12.7 | 12.7 | 0.1 | 0.9 | 0.5 | 1.9 | 0.0 | 0.0 | 60.6 | 69.1 | 0.0 | 0.0 | 35.6 | 18.8 |
| Faganeanea | 0.3 | 0.3 | 4173.3 | 4155.6 | 290.9 | 289.2 | 427.0 | 425.3 | 21.7 | 17.0 | 112.0 | 115.0 | 31.7 | 31.7 | 0.0 | 0.0 | 42.8 | 42.4 | 10.2 | 16.3 | 32.4 | 15.2 |
| Fagatogo | 0.3 | 0.3 | 4393.5 | 4369.5 | 304.3 | 302.6 | 447.8 | 445.6 | 36.0 | 33.6 | 2.3 | 4.6 | 2.4 | 3.7 | 0.0 | 0.0 | 8.9 | 14.5 | 5.8 | 8.0 | 77.1 | 60.3 |
| Fatumafuti | 0.3 | 0.3 | 3224.8 | 3230.2 | 210.5 | 211.0 | 335.5 | 336.0 | 30.0 | 30.0 | 0.0 | 0.0 | 0.0 | 0.2 | 0.0 | 0.0 | 23.3 | 27.9 | 8.1 | 13.1 | 59.6 | 28.1 |
| Futiga | 0.4 | 0.4 | 3281.0 | 3266.0 | 229.0 | 227.7 | 335.3 | 334.0 | 16.6 | 13.6 | 93.3 | 92.1 | 14.6 | 13.1 | 0.0 | 0.1 | 28.4 | 45.2 | 3.2 | 4.7 | 67.5 | 48.9 |
| Iliili | 0.4 | 0.4 | 3121.1 | 3113.8 | 212.9 | 212.3 | 324.0 | 323.4 | 21.5 | 18.8 | 60.0 | 57.4 | 3.0 | 2.9 | 0.0 | 0.1 | 3.3 | 6.4 | 3.8 | 9.3 | 92.1 | 82.4 |
| Laulii | 0.3 | 0.3 | 3563.8 | 3536.3 | 245.9 | 243.3 | 367.0 | 364.4 | 26.7 | 22.3 | 95.3 | 93.9 | 22.7 | 23.1 | 0.1 | 0.0 | 10.5 | 14.2 | 12.2 | 22.7 | 70.2 | 44.6 |
| Leloaloa | 0.3 | 0.2 | 4390.9 | 4394.9 | 307.4 | 307.8 | 452.3 | 452.5 | 18.4 | 16.0 | 374.8 | 361.3 | 35.0 | 37.2 | 0.5 | 0.3 | 11.6 | 17.1 | 21.1 | 25.0 | 22.9 | 14.6 |
| Leone | 0.3 | 0.3 | 3219.0 | 3315.4 | 226.7 | 235.5 | 329.2 | 338.0 | 17.4 | 16.2 | 42.7 | 54.9 | 8.2 | 9.2 | 0.0 | 0.0 | 10.3 | 19.5 | 1.4 | 2.0 | 85.9 | 73.8 |
| Malaeimi | 0.4 | 0.4 | 4188.3 | 4212.7 | 299.7 | 301.9 | 423.0 | 425.3 | 14.8 | 13.4 | 97.1 | 98.2 | 18.2 | 18.2 | 1.8 | 2.3 | 14.0 | 25.8 | 2.6 | 5.2 | 78.2 | 62.4 |
| Malaeloa Aitulagi | 0.4 | 0.4 | 3849.7 | 3932.0 | 281.5 | 289.0 | 388.0 | 395.6 | 23.2 | 19.1 | 106.7 | 133.6 | 13.9 | 16.4 | 0.0 | 0.0 | 34.9 | 52.7 | 3.5 | 5.6 | 59.2 | 39.3 |
| Masausi | 0.3 | 0.4 | 2425.9 | 2438.0 | 165.3 | 166.1 | 252.3 | 253.5 | 16.7 | 17.5 | 13.1 | 16.1 | 6.1 | 7.2 | 0.0 | 0.0 | 57.1 | 71.8 | 4.7 | 3.4 | 33.9 | 13.6 |
| Nua | 0.3 | 0.3 | 3734.3 | 3733.2 | 275.3 | 275.3 | 377.0 | 376.8 | 12.6 | 11.6 | 116.1 | 115.9 | 27.4 | 26.2 | 0.0 | 0.0 | 17.2 | 32.6 | 3.8 | 2.7 | 59.8 | 35.9 |
| Pago Pago | 0.3 | 0.4 | 4269.6 | 4274.0 | 297.6 | 298.0 | 436.3 | 436.7 | 31.8 | 29.0 | 164.1 | 164.2 | 26.4 | 25.8 | 0.0 | 0.0 | 14.1 | 28.3 | 1.8 | 5.6 | 80.9 | 59.6 |
| Pavaiai | 0.4 | 0.4 | 4042.2 | 4048.7 | 293.2 | 294.1 | 407.7 | 408.3 | 12.4 | 11.5 | 130.4 | 133.8 | 8.1 | 8.5 | 0.1 | 0.2 | 10.3 | 19.1 | 0.5 | 1.6 | 75.1 | 63.3 |
| Satala-Anua-Atuu | 0.2 | 0.2 | 4526.4 | 4518.2 | 318.0 | 317.2 | 461.3 | 460.7 | 30.4 | 27.0 | 402.4 | 378.6 | 38.8 | 39.2 | 0.0 | 0.0 | 1.7 | 6.0 | 16.2 | 26.3 | 65.4 | 44.4 |
| Seetaga | 0.3 | 0.3 | 3818.9 | 3772.2 | 283.3 | 279.5 | 384.4 | 379.9 | 12.5 | 11.6 | 163.2 | 165.8 | 29.0 | 29.4 | 0.0 | 0.0 | 25.8 | 38.9 | 0.1 | 0.1 | 54.0 | 30.0 |
| Tafuna | 0.3 | 0.3 | 3125.5 | 3122.3 | 210.9 | 210.8 | 327.2 | 326.7 | 23.1 | 21.5 | 28.1 | 27.7 | 3.4 | 3.4 | 0.5 | 1.0 | 0.3 | 1.0 | 2.8 | 7.2 | 95.5 | 88.5 |
| Taputimu | 0.4 | 0.4 | 2983.3 | 2995.9 | 203.0 | 204.1 | 307.7 | 308.9 | 14.9 | 14.1 | 35.3 | 35.9 | 1.8 | 1.7 | 0.1 | 0.1 | 12.2 | 22.7 | 0.2 | 1.6 | 87.3 | 75.5 |
| Tula | 0.4 | 0.4 | 2136.7 | 2128.9 | 139.0 | 138.3 | 230.5 | 229.9 | 30.3 | 23.5 | 18.0 | 16.7 | 9.6 | 8.9 | 0.0 | 0.0 | 19.5 | 32.7 | 6.1 | 10.3 | 70.6 | 42.7 |
| Utumea West | 0.4 | 0.3 | 3371.0 | 3376.7 | 246.3 | 246.8 | 341.7 | 342.2 | 12.0 | 10.0 | 160.3 | 155.6 | 24.7 | 24.3 | 0.0 | 0.0 | 57.6 | 58.7 | 0.0 | 0.0 | 23.2 | 10.1 |
| Vaitogi | 0.4 | 0.4 | 2838.4 | 2842.5 | 188.4 | 188.8 | 298.5 | 298.8 | 15.3 | 14.0 | 38.2 | 40.4 | 2.3 | 2.6 | 0.2 | 0.2 | 14.8 | 27.1 | 1.3 | 2.1 | 82.7 | 66.8 |
| Vatia | 0.4 | 0.3 | 3192.1 | 3165.1 | 213.3 | 211.6 | 331.9 | 329.1 | 23.1 | 19.0 | 46.0 | 37.3 | 20.0 | 17.7 | 0.2 | 0.1 | 29.4 | 45.0 | 7.5 | 7.0 | 56.1 | 31.5 |
References
- Manhenje, I.; Galan-Puchades, M.T.; Fuentes, M.V. Socio-environmental variables and transmission risk of lymphatic filariasis in central and northern Mozambique. Geospat. Health 2013, 7, 391–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stensgaard, A.-S.; Vounatsou, P.; Onapa, A.W.; Simonsen, P.E.; Pedersen, E.M.; Rahbek, C.; Kristensen, T.K. Bayesian geostatistical modelling of malaria and lymphatic filariasis infections in Uganda: Predictors of risk and geographical patterns of co-endemicity. Malar. J. 2011, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edirisinghe, G. Socio-economic and Environmental Determinants of Filariasis in Matara District of Sri Lanka. Am. Acad. Sci. Res. J. Eng. Technol. Sci. 2017, 32, 105–118. Available online: https://asrjetsjournal.org/index.php/American_Scientific_Journal/article/view/3015/1156 (accessed on 22 April 2022).
- Cano, J.; Rebollo, M.; Golding, N.; Pullan, R.L.; Crellen, T.; Soler, A.; Kelly-Hope, L.A.; Lindsay, S.W.; Hay, S.I.; Bockarie, M.J.; et al. The global distribution and transmission limits of lymphatic filariasis: Past and present. Parasites Vectors 2014, 7, 466. [Google Scholar] [CrossRef] [PubMed]
- Ton, T.G.N.; Mackenzie, C.; Molyneux, D.H. The burden of mental health in lymphatic filariasis. Infect. Dis. Poverty 2015, 4, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garchitorena, A.; Raza-Fanomezanjanahary, E.M.; Mioramalala, S.A.; Chesnais, C.B.; Ratsimbasoa, C.A.; Ramarosata, H.; Bonds, M.H.; Rabenantoandro, H. Towards elimination of lymphatic filariasis in southeastern Madagascar: Successes and challenges for interrupting transmission. PLoS Negl. Trop. Dis. 2018, 12, e0006780. [Google Scholar] [CrossRef] [Green Version]
- WHO. Global programme to eliminate lymphatic filariasis: Progress report, 2020. WHO Wkly. Epidemiol. Rec. 2021, 96, 497–508. Available online: https://www.who.int/publications/i/item/who-wer9641-497-508 (accessed on 9 January 2022).
- Yajima, A.; Ichimori, K. Progress in the elimination of lymphatic filariasis in the Western Pacific Region: Successes and challenges. Int. Health 2020, 13 (Suppl. S1), S10–S16. [Google Scholar] [CrossRef]
- WHO. Kiribati Eliminates Lymphatic Filariasis Western Pacific; WHO Press: Manila, Philippines, 2019. Available online: https://www.who.int/westernpacific/news/item/07-10-2019-kiribati-eliminates-lymphatic-filariasis (accessed on 7 October 2019).
- WHO. Guideline: Alternative Mass Drug Administration Regimens to Eliminate Lymphatic Filariasis; World Health Organization: Geneva, Switzerland, 2017. Available online: https://apps.who.int/iris/bitstream/handle/10665/259381/9789241550161-eng.pdf (accessed on 17 March 2022).
- Coutts, S.P.; King, J.D.; Pa’Au, M.; Fuimaono, S.; Roth, J.; King, M.R.; Lammie, P.J.; Lau, C.L.; Graves, P.M. Prevalence and risk factors associated with lymphatic filariasis in American Samoa after mass drug administration. Trop. Med. Health 2017, 45, 22. [Google Scholar] [CrossRef]
- Graves, P.M.; Sheridan, S.; Fuimaono, S.; Lau, C.L. Demographic, socioeconomic and disease knowledge factors, but not population mobility, associated with lymphatic filariasis infection in adult workers in American Samoa in 2014. Parasit. Vectors 2020, 13, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Lau, C.L.; Won, K.Y.; Becker, L.; Soares Magalhaes, R.J.; Fuimaono, S.; Melrose, W.; Lammie, P.J.; Graves, P.M. Seroprevalence and Spatial Epidemiology of Lymphatic Filariasis in American Samoa after Successful Mass Drug Administration. PLoS Negl. Trop. Dis. 2014, 8, e3297. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.L.; Sheridan, S.; Ryan, S.; Roineau, M.; Andreosso, A.; Fuimaono, S.; Tufa, J.; Graves, P.M. Detecting and confirming residual hotspots of lymphatic filariasis transmission in American Samoa 8 years after stopping mass drug administration. PLoS Negl. Trop. Dis. 2017, 11, e0005914. [Google Scholar] [CrossRef] [PubMed]
- Sheel, M.; Sheridan, S.; Gass, K.; Won, K.; Fuimaono, S.; Kirk, M.; Gonzales, A.; Hedtke, S.M.; Graves, P.M.; Lau, C.L. Identifying residual transmission of lymphatic filariasis after mass drug administration: Comparing school-based versus community-based surveillance—American Samoa, 2016. PLoS Negl. Trop. Dis. 2018, 12, e0006583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- QIMR Berghofer Medical Research Institute. Draft Assessment Report for Aedes polynesiensis: Department of Agriculture, Water and the Environment. 2016. Available online: https://www.awe.gov.au/sites/default/files/env/consultations/d733c7c1-5d70-429e-a6b5-a03bc2ddfd9f/files/draft-risk-assessment-report-aedes-polynesiensis.pdf (accessed on 9 April 2022).
- Chaves, L.F.; Morrison, A.C.; Kitron, U.D.; Scott, T.W. Nonlinear impacts of climatic variability on the density-dependent regulation of an insect vector of disease. Glob. Chang. Biol. 2012, 18, 457–468. [Google Scholar] [CrossRef]
- Lardeux, F.; Cheffort, J. Ambient temperature effects on the extrinsic incubation period of Wuchereria bancrofti in Aedes polynesiensis: Implications for filariasis transmission dynamics and distribution in French Polynesia. Med. Vet. Entomol. 2001, 15, 167–176. [Google Scholar] [CrossRef]
- Slater, H.; Michael, E. Predicting the Current and Future Potential Distributions of Lymphatic Filariasis in Africa Using Maximum Entropy Ecological Niche Modelling. PLoS ONE 2012, 7, e32202. [Google Scholar] [CrossRef]
- May, R.M.; Deyle, E.; Hsieh, C.-H.; Munch, S. Detecting Causality in Complex Ecosystems. Science 2012, 228, 496–500. [Google Scholar]
- Dhimal, M.; Gautam, I.; Kreß, A.; Müller, R.; Kuch, U. Spatio-Temporal Distribution of Dengue and Lymphatic Filariasis Vectors along an Altitudinal Transect in Central Nepal. PLoS Negl. Trop. Dis. 2014, 8, e3035. [Google Scholar] [CrossRef]
- Lau, C.L.; Meder, K.; Mayfield, H.J.; Kearns, T.; McPherson, B.; Naseri, T.; Thomsen, R.; Hedtke, S.M.; Sheridan, S.; Gass, K.; et al. Lymphatic filariasis epidemiology in Samoa in 2018: Geographic clustering and higher antigen prevalence in older age groups. PLoS Negl. Trop. Dis. 2020, 14, e0008927. [Google Scholar] [CrossRef]
- Lau, C.L.; Won, K.Y.; Lammie, P.J.; Graves, P.M. Lymphatic Filariasis Elimination in American Samoa: Evaluation of Molecular Xenomonitoring as a Surveillance Tool in the Endgame. PLoS Negl. Trop. Dis. 2016, 10, e0005108. [Google Scholar] [CrossRef]
- Schmaedick, M.A.; Koppel, A.L.; Pilotte, N.; Torres, M.; Williams, S.A.; Dobson, S.L.; Lammie, P.J.; Won, K.Y. Molecular Xenomonitoring Using Mosquitoes to Map Lymphatic Filariasis after Mass Drug Administration in American Samoa. PLoS Negl. Trop. Dis. 2014, 8, e3087. [Google Scholar] [CrossRef] [PubMed]
- Cameron, M.M.; Ramesh, A. The use of molecular xenomonitoring for surveillance of mosquito-borne diseases. Philos. Trans. R. Soc. B 2021, 376, 20190816. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, M.A.; Salamah, M.M.I.; Thabet, H.S.; Tawfik, R.A.; Mehrez, M.M.; Hamdy, D.M. Molecular xenomonitoring (MX) and transmission assessment survey (TAS) of lymphatic filariasis elimination in two villages, Menoufyia Governorate, Egypt. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Dorkenoo, M.A.; De Souza, D.K.; Apetogbo, Y.; Oboussoumi, K.; Yehadji, D.; Tchalim, M.; Etassoli, S.; Koudou, B.; Ketoh, G.K.; Sodahlon, Y.; et al. Molecular xenomonitoring for post-validation surveillance of lymphatic filariasis in Togo: No evidence for active transmission. Parasit. Vectors 2018, 11, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irish, S.R.; Al-Amin, H.M.; Paulin, H.N.; Mahmood, A.S.M.S.; Khan, R.K.; Muraduzzaman, A.K.M.; Worrell, C.M.; Flora, M.S.; Karim, M.J.; Shirin, T.; et al. Molecular xenomonitoring for Wuchereria bancrofti in Culex quinquefasciatus in two districts in Bangladesh supports transmission assessment survey findings. PLoS Negl. Trop. Dis. 2018, 12, e0006574. [Google Scholar] [CrossRef] [Green Version]
- Lau, C.L.; Sheel, M.; Gass, K.; Fuimaono, S.; David, M.C.; Won, K.Y.; Sheridan, S.; Graves, P.M. Potential strategies for strengthening surveillance of lymphatic filariasis in American Samoa after mass drug administration: Reducing ‘number needed to test’ by targeting older age groups, hotspots, and household members of infected persons. PLoS Negl. Trop. Dis. 2020, 14, e0008916. [Google Scholar] [CrossRef]
- Cadavid Restrepo, A.M.; Gass, K.; Won, K.Y.; Sheel, M.; Robinson, K.; Graves, P.M.; Fuimaono, S.; Lau, C.L. Potential Use of Antibodies to Provide an Earlier Indication of Lymphatic Filariasis Resurgence in Post-Mass Drug Administration Surveillance, American Samoa. Int. J. Infect. Dis. 2022, 117, 378–386. [Google Scholar] [CrossRef]
- Souza, D.d.; Kelly-Hope, L.; Lawson, B.; Wilson, M.; Boakye, D. Environmental Factors Associated with the Distribution of Anopheles gambiae s.s. in Ghana; an Important Vector of Lymphatic Filariasis and Malaria. PLoS ONE 2010, 5, e9927. [Google Scholar] [CrossRef] [Green Version]
- Eneanya, O.A.; Cano, J.; Dorigatti, I.; Anagbogu, I.; Okoronkwo, C.; Garske, T.; Donnelly, C.A. Environmental suitability for lymphatic filariasis in Nigeria. Parasites Vectors 2018, 11, 513. [Google Scholar] [CrossRef] [Green Version]
- The World Bank. American Samoa. Available online: https://data.worldbank.org/country/AS (accessed on 3 April 2022).
- Keener, V.; Grecni, Z.; Tagarino, K.A.; Shuler, C.; Miles, W. Climate Change in American Samoa: Indicators and Considerations for Key Sectors. Available online: https://eastwestcentre.org/PIRCA-AmericanSamoa (accessed on 17 May 2022).
- NOAA. American Samoa Baseline Observatory Pago Pago: Earth System Research Laboratories. 2021. Available online: https://gml.noaa.gov/obop/smo/ (accessed on 12 April 2022).
- Holst-Rice, S.; Messina, A.; Biggs, T.; Vargas-Angel, B.; Whitall, D. Baseline Assessment of Faga’alu Watershed: A Ridge to Reef Assessment in Support of Sediment Reduction Activities and Future Evaluation of their Success. 2016. Available online: https://www.researchgate.net/publication/298423519_Baseline_Assessment_of_Faga’alu_Watershed_A_Ridge_to_Reef_Assessment_in_Support_of_Sediment_Reduction_Activities_and_Future_Evaluation_of_Their_Success (accessed on 10 April 2022).
- Esri, M.; Earth Star Geographics. Topographic Basemap 2021. Available online: https://www.arcgis.com/home/item.html?id=c61ad8ab017d49e1a82f580ee1298931 (accessed on 28 March 2022).
- Wangdi, K.; Sheel, M.; Fuimaono, S.; Graves, P.M.; Lau, C.L. Lymphatic filariasis in 2016 in American Samoa: Identifying clustering and hotspots using non-spatial and three spatial analytical methods. PLoS Negl. Trop. Dis. 2022, 16, e0010262. [Google Scholar] [CrossRef]
- Villages of Tutuila and Buildings, Tutuila, UTM & Lat/Long. National Marine Sanctuary of American Samoa. 2003. Available online: https://dusk.geo.orst.edu/djl/samoa/ (accessed on 28 March 2022).
- MODIS/Terra Land Surface Temperature/Emissivity 8-Day L3 Global 1km SIN Grid V061. 2021. Available online: https://doi.org/10.5067/MODIS/MOD11A2.061 (accessed on 28 March 2022).
- Local Climatological Data Set. 2016. Available online: https://americansamoa.prism.spc.int/environment-statistics/rainfall.html (accessed on 17 April 2022).
- Tutuila. American Samoa 1/3 arc-second MHW Coastal Digital Elevation Model Data File. 2013. Available online: https://www.ncei.noaa.gov/metadata/geoportal/rest/metadata/item/gov.noaa.ngdc.mgg.dem:4610/html# (accessed on 28 March 2022).
- Global Land Use/Land Cover with Sentinel-2 and Deep Learning. 2022. Available online: https://env1.arcgis.com/arcgis/rest/services/Sentinel2_10m_LandCover/ImageServer (accessed on 7 April 2022).
- Landsat Collection 2 Level-2 Surface Reflectance and Surface Temperature. 2021. Available online: https://www.usgs.gov/landsat-missions/landsat-collection-2-level-2-science-products (accessed on 7 April 2022).
- American Population Raster American Samoa 2020 Data Input: Settlement footprint from Facebook’s High-Resolution Population Density Maps Pacific Data Hub. 2020. Available online: https://stats.pacificdata.org/vis?fs[0]=Topic%2C0%7CPopulation%23POP%23&pg=0&fc=Topic&df[ds]=ds%3ASPC2&df[id]=DF_POP_DENSITY&df[ag]=SPC&df[vs]=1.0&pd=2021%2C2021&dq=A.AS.&ly[cl]=INDICATOR (accessed on 28 March 2022).
- Mercer, D.R.; Marie, J.; Bossin, H.; Faaruia, M.; Tetuanui, A.; Sang, M.C.; Dobson, S.L. Estimation of Population Size and Dispersal of Aedes polynesiensis on Toamaro motu, French Polynesia. J. Med. Entomol. 2012, 49, 971–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esri Inc. ArcGIS Pro, 2.8.6th ed.; Esri Inc.: Redlands, CA, USA, 2020. [Google Scholar]
- NOAA Climate Data Record (CDR) of AVHRR Normalized Difference Vegetation Index (NDVI). 2019. Available online: https://doi.org/10.7289/V5ZG6QH9 (accessed on 9 April 2022).
- RStudio: Integrated Development for R. PBC. 2020. Available online: http://www.rstudio.com/ (accessed on 7 March 2022).
- Kwarteng, E.V.S.; Andam-Akorful, S.A.; Kwarteng, A.; Asare, D.-C.B.; Quaye-Ballard, J.A.; Osei, F.B.; Duker, A.A. Spatial variation in lymphatic filariasis risk factors of hotspot zones in Ghana. BMC Public Health 2021, 21, 230. [Google Scholar] [CrossRef]
- Stanton, M.C.; Molyneux, D.H.; Kyelem, D.; Bougma, R.W.; Koudou, B.G.; Kelly-Hope, L.A. Baseline drivers of lymphatic filariasis in Burkina Faso. Geospat. Health 2013, 8, 159–173. [Google Scholar] [CrossRef]
- Palaniyandi, M. A geo-spatial modeling for mapping of filariasis transmission risk in India, using remote sensing and GIS. Int. J. Mosq. Res. 2014, 1, 20–28. [Google Scholar]
- Mwase, E.; Stensgaard, A.-S.; Nsakashalo-Senkwe, M.; Mubila, L.; Mwansa, J.; Songolo, P.; Shawa, S.; Simonsen, P. Mapping the Geographical Distribution of Lymphatic Filariasis in Zambia. PLoS Negl. Trop. Dis. 2014, 8, e2714. [Google Scholar] [CrossRef]
- Onapa, A.W.; Simonsen, P.E.; Baehr, I.; Pedersen, E.M. Rapid assessment of the geographical distribution of lymphatic filariasis in Uganda, by screening of schoolchildren for circulating filarial antigens. Ann. Trop. Med. Parasitol. 2005, 99, 141–153. [Google Scholar] [CrossRef]
- Slater, H.; Michael, E. Mapping, Bayesian Geostatistical Analysis and Spatial Prediction of Lymphatic Filariasis Prevalence in Africa. PLoS ONE 2013, 8, e71574. [Google Scholar]
- Rwegoshora, R.T.; Pedersen, E.M.; Mukoko, D.A.; Meyrowitsch, D.W.; Masese, N.; Malecela-Lazaro, M.N.; Ouma, J.H.; Michael, E.; Simonsen, P.E. Bancroftian filariasis: Patterns of vector abundance and transmission in two East African communities with different levels of endemicity. Ann. Trop. Med. Parasitol. 2005, 99, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Grziwotz, F.; Strauß, J.F.; Hsieh, C.-H.; Telschow, A. Empirical Dynamic Modelling Identifies different Responses of Aedes Polynesiensis Subpopulations to Natural Environmental Variables. Nature 2018, 8, 16768. [Google Scholar] [CrossRef] [Green Version]
- Nurjazuli, N.; Santjaka, A. The Potential Sources of Transmission and Distribution of Lymphatic Filariasis in Semarang City, Central Java, Indonesia. Unnes J. Public Health 2020, 9, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Chesnais, C.B.; Awaca-Uvon, N.-P.; Vlaminck, J.; Tambwe, J.-P.; Weil, G.J.; Pion, S.D.; Boussinesq, M. Risk factors for lymphatic filariasis in two villages of the Democratic Republic of the Congo. Parasit. Vectors 2019, 12, 162. [Google Scholar] [CrossRef] [PubMed]
- Hussaini, A.; Isaac, C.; Rahimat, H.; Collins, I.; Cedric, O.; Solomon, E. The Burden of Bancroftian Filariasis in Nigeria: A Review. Ethiop. J. Health Sci. 2020, 30, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Srividya, S.; Subramanian, S.; Sadanandane, C.; Vasuki, V.; Jambulingam, P. Determinants of transmission hotspots and filarial infection in households after eight rounds of mass drug administration in India. Trop. Med. Int. Health 2018, 23, 1251–1258. [Google Scholar] [CrossRef] [PubMed]




| Variable Description | Dataset | Source | Spatial Resolution | Temporal Resolution |
|---|---|---|---|---|
| Temperature | MODIS/Terra Land Surface Temperature/Emissivity 8-Day L3 Global 1 km SIN Grid V06 [40] | USGS Earth Explorer | 1 km | Weekly (2016) |
| Rainfall | Department of Commerce–Local Climate Data [41] | Pacific Environment Data Portal | 1 km | Monthly (2016) |
| Elevation | NOAA–American Samoa 1/3 arc-second MHW Coastal Digital Elevation Model [42] | USGS Earth Explorer | 10 m | 1984-2012 |
| Landcover | Sentinel-2 10 m Land Use/Land Cover Timeseries [43] | Esri | 10 m | 2017-2021 |
| Surface reflectance (used to derive normalized difference vegetation index (NDVI)) | Landsat Collection 2 Level-2 Surface Reflectance and Surface Temperature [44] | USGS Earth Explorer | 30 m | Weekly (2016) |
| Population density | High Resolution Settlement Layer (HRSL) [45] | Pacific Data Hub based on Data for Good at Meta dataset using census data from 2010/2011 | 100 m | 2020 (Based on 2011 census with a population growth rate of 0.23% applied) |
| Landcover Class | Description |
|---|---|
| Crop | Areas with human planted vegetation below tree height. |
| Rangeland | Areas of shrubs, natural fields, and grassland, with no evidence of artificial plotting or tall tree coverage. |
| Trees | Areas of tall and dense vegetation, around 4.5 m or higher, including forests, swamps, mangroves, and savannas. |
| Built/Urban | Areas of human development including major paved roads, houses, towns, and large areas of asphalt. |
| Variable | Buffer Size | Mean | Standard Deviation | Minimum | Maximum |
|---|---|---|---|---|---|
| Village population | - | 786.16 | 804.42 | 47.00 | 3195.00 |
| Participants/village | - | 89.03 | 81.10 | 5.00 | 307.00 |
| Antigen (counts/village) | - | 4.50 | 6.45 | 0.00 | 31.00 |
| Mf (counts/village) | - | 1.13 | 2.46 | 0.00 | 12.00 |
| Wb123 Ab (counts/village) | - | 22.80 | 21.32 | 2.00 | 83.00 |
| Bm14 Ab (counts/village) | - | 11.67 | 12.84 | 0.00 | 57.00 |
| Bm33 Ab (counts/village) | - | 40.63 | 40.17 | 4.00 | 175.00 |
| Annual rainfall (mm) | 50 m | 3426.79 | 654.41 | 2136.74 | 4526.41 |
| 100 m | 3430.70 | 654.78 | 2128.86 | 4518.20 | |
| Average dry season rainfall per month (mm) | 50 m | 239.70 | 50.75 | 139.01 | 317.97 |
| 100 m | 240.10 | 50.95 | 138.28 | 317.21 | |
| Average wet season rainfall per month (mm) | 50 m | 351.70 | 63.60 | 230.54 | 461.31 |
| 100 m | 352.03 | 63.57 | 229.91 | 460.71 | |
| Average elevation (m) | 50 m | 95.51 | 95.66 | 0.00 | 402.41 |
| 100 m | 96.28 | 92.07 | 0.05 | 378.61 | |
| Average slope gradient (degrees) | 50 m | 15.96 | 11.54 | 0.00 | 39.17 |
| 100 m | 16.23 | 11.49 | 0.23 | 39.18 | |
| Average NVDI | 50 m | 0.34 | 0.05 | 0.21 | 0.41 |
| 100 m | 0.34 | 0.05 | 0.23 | 0.41 | |
| Crop cover in inhabited buffer zone (%) | 50 m | 0.11 | 0.34 | 0.00 | 1.78 |
| 100 m | 0.14 | 0.44 | 0.00 | 2.28 | |
| Tree cover in inhabited buffer zone (%) | 50 m | 26.35 | 18.89 | 0.29 | 66.59 |
| 100 m | 35.96 | 20.63 | 1.02 | 78.29 | |
| Rangeland cover in inhabited buffer zone (%) | 50 m | 5.36 | 5.79 | 0.00 | 21.09 |
| 100 m | 8.27 | 8.31 | 0.00 | 27.04 | |
| Urban cover in inhabited buffer zone (%) | 50 m | 59.10 | 21.88 | 16.48 | 95.54 |
| 100 m | 40.72 | 22.88 | 9.56 | 88.53 | |
| Average population density (person/km2) | 50 m | 20.30 | 6.74 | 12.00 | 36.00 |
| 100 m | 18.16 | 6.00 | 10.00 | 33.58 |
| Microfilaria | Antigen | Wb123 Ab | Bm14 Ab | Bm33 Ab | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| RR (95% CI) | p-Value | RR (95% CI) | p-Value | RR (95% CI) | p-Value | RR (95% CI) | p-Value | RR (95% CI) | p-Value | |
| Climatic Variables | ||||||||||
| Annual Rainfall | - | - | - | - | - | - | 1.0 (1.0, 1.0) | 0.01 | - | - |
| Dry Season Rainfall | - | - | - | - | - | - | - | - | - | - |
| Wet Season Rainfall | 1.01 (1.0, 1.02) | <0.01 | 1.01 (1.0, 1.01) | <0.01 | 1.0 (0.99, 1.0) | 0.1 | - | - | 1.0 (1, 1) | <0.01 |
| Population Variables | ||||||||||
| Population Density | 0.88 (0.81, 0.96) | <0.01 | 0.89 (0.85, 0.93) | <0.01 | 0.96 (1.03, 1.06) | <0.01 | 0.96 (0.95, 0.98) | <0.01 | 0.98 (0.96, 0.99) | <0.01 |
| Topographical and Landcover Variables | ||||||||||
| Elevation | - | - | - | - | - | - | 1.0 (1.0, 1.0) | 0.04 | - | - |
| Slope gradient | 0.91 (0.84, 0.98) | 0.02 | 0.91 (0.88, 0.94) | <0.01 | 1.01 (1.0, 1.02) | 0.04 | - | - | 1.01 (0.99, 1.02) | 0.09 |
| NDVI | - | - | 0.00 (0, 0.06) | <0.01 | 0.03 (0, 0.31) | <0.01 | 0.001 (0.0, 0.02) | <0.01 | 0.01 (0.01, 0.58) | <0.01 |
| Crop class | - | - | - | - | 0.85(0.71, 1.01) | 0.07 | - | - | 0.78 (0.67, 0.89) | <0.01 |
| Tree class | 1.18 (1.09, 1.29) | <0.01 | 1.07 (1.04, 1.09) | <0.01 | 1.05 (1.03, 1.06) | <0.01 | 1.09 (1.07, 1.11) | <0.01 | 1.06 (1.05, 1.08) | <0.01 |
| Urban class | 1.09 (1.01, 1.18) | 0.02 | 1.02 (0.99, 1.04) | 0.17 | 1.02 (1.01, 1.04) | <0.01 | 1.04 (1.03, 1.06) | <0.01 | 1.04 (1.03, 1.08) | <0.01 |
| Rangeland class | 1.42 (1.17, 1.76) | <0.01 | 1.12 (1.06, 1.19) | 0.03 | - | - | - | - | 1.05 (1.02, 1.08) | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemin, M.E.; Cadavid Restrepo, A.; Mayfield, H.J.; Lau, C.L. Spatially Explicit Environmental Factors Associated with Lymphatic Filariasis Infection in American Samoa. Trop. Med. Infect. Dis. 2022, 7, 295. https://doi.org/10.3390/tropicalmed7100295
Lemin ME, Cadavid Restrepo A, Mayfield HJ, Lau CL. Spatially Explicit Environmental Factors Associated with Lymphatic Filariasis Infection in American Samoa. Tropical Medicine and Infectious Disease. 2022; 7(10):295. https://doi.org/10.3390/tropicalmed7100295
Chicago/Turabian StyleLemin, Morgan E., Angela Cadavid Restrepo, Helen J. Mayfield, and Colleen L. Lau. 2022. "Spatially Explicit Environmental Factors Associated with Lymphatic Filariasis Infection in American Samoa" Tropical Medicine and Infectious Disease 7, no. 10: 295. https://doi.org/10.3390/tropicalmed7100295