Catch Me If You Can! RNA Silencing-Based Improvement of Antiviral Plant Immunity


Centre for BioSystems, Institute of Phytopathology, Land Use and Nutrition, Justus Liebig University, Heinrich-Buff-Ring 26, D-35392 Giessen, Germany
*
Author to whom correspondence should be addressed.
Viruses 2019, 11(7), 673; https://doi.org/10.3390/v11070673
Submission received: 4 April 2019
/
Revised: 11 July 2019
/
Accepted: 17 July 2019
/
Published: 23 July 2019
(This article belongs to the Special Issue Plant Virus Ecology and Biodiversity)
Abstract
:Viruses are obligate parasites which cause a range of severe plant diseases that affect farm productivity around the world, resulting in immense annual losses of yield. Therefore, control of viral pathogens continues to be an agronomic and scientific challenge requiring innovative and ground-breaking strategies to meet the demands of a growing world population. Over the last decade, RNA silencing has been employed to develop plants with an improved resistance to biotic stresses based on their function to provide protection from invasion by foreign nucleic acids, such as viruses. This natural phenomenon can be exploited to control agronomically relevant plant diseases. Recent evidence argues that this biotechnological method, called host-induced gene silencing, is effective against sucking insects, nematodes, and pathogenic fungi, as well as bacteria and viruses on their plant hosts. Here, we review recent studies which reveal the enormous potential that RNA-silencing strategies hold for providing an environmentally friendly mechanism to protect crop plants from viral diseases.
1. Introduction
2. RNA Silencing-Mediated Antiviral Plant Immunity
RNA silencing (also termed RNA interference, RNAi) is a conserved regulatory mechanism of gene expression in eukaryotic organisms which is triggered by dsRNA-provoking gene silencing by sequence-specific degradation of complementary mRNA transcripts (post-transcriptional gene silencing, PTGS) [34,35] or by inhibition of transcription (transcriptional gene silencing, TGS) [36]. RNA silencing plays a pivotal role in diverse cellular, developmental and physiological processes, regulating gene expression via small noncoding RNAs (sRNAs) [37] and is associated with protection against viral infection, the control of epigenetic modifications, regulation of genome stability, curbing of transposon movement and regulation of heterochromatin formation [36].
The ultimate trigger initiating/eliciting RNA silencing is dsRNA, which is a replication intermediate generated by viral RNA-dependent RNA polymerases (RDRs) of plant-infecting RNA viruses [38]. RNA-silencing mechanisms start with initial processing or cleavage of a precursor dsRNA into short 21–24 nucleotide (nt) small interfering (siRNA) or micro RNA (miRNA) duplexes [39] by an RNaseIII-like enzyme called Dicer (DCL) [40,41]. Double-stranded siRNAs are incorporated into an RNA-induced silencing complex (RISC) containing an Argonaute (AGO) protein that has a sRNA-binding domain and an endonucleolytic activity for cleavage of target RNAs [42]. The activated RISC subsequently unwinds siRNAs, thereby generating a sense (passenger) and an antisense (guide) strand in an ATP-dependent reaction. While the sense strand is degraded, the RISC containing the antisense strand subsequently targets a complementary mRNA transcript via base pairing interaction, degrades the mRNA and thereby inhibits protein biosynthesis [43,44,45].
RNA silencing is the best-studied antiviral defence mechanism in plants [46,47]. Notably, a few studies reported a similar antiviral function in mammals [48,49,50,51]; however, its diversity depends mainly on the existence of multiple copies of the core RNA-silencing pathway components, which, presumably, are the result of gene duplication followed by specialization [52,53]. For example, the model plant Arabidopsis thaliana has four DCLs, ten AGOs and six RDRs, which are involved in different silencing-related pathways [46]. In Arabidopsis, DCL3, DCL4 and DCL2 are important for virus-induced RNA silencing, and DCL3 is also crucial against DNA viruses [54,55,56,57]. AGO1, AGO2, AGO4, AGO5, AGO7 and AGO10 of Arabidopsis are the central players in antiviral RNA silencing [58]. RDR1, RDR2 and RDR6 are shown to display antiviral activity amplifying virus-derived small interfering RNAs or function as a silencing signal [59] (Figure 1). Besides RDR mediation, amplification of small-interfering viral RNA (vsiRNA), short distance (cell-to-cell) as well as long distance (e.g., phloem) spreading of silencing signals is an important aspect of PTGS for establishing systemic antiviral immunity. The reception of long-distance mRNA PTGS and the generation of secondary siRNAs in the recipient cell/tissue requires coordinated activity of a set of proteins. These proteins include AGO1/AGO2 [60], DCL2 [61], RDR6 [52] and SGS3 (SUPPRESSOR OF GENE SILENCING 3) [62], which orchestrate to facilitate the silencing signal amplification process. Notably, proteins associated with TGS pathways also contribute to systemic PTGS, as RNA polymerase IVa, RDR2, AGO4 and DCL3 are involved in the reception of long-distance silencing [63,64].
Interestingly, there is increasing evidence that PTGS and TGS mediated by endogenous sRNAs are key regulators of PTI and ETI [65,66].
Given the complex regulatory mechanisms of antiviral plant defence responses, viruses often cause severe diseases with immense economic losses in agricultural production, suggesting that they have evolved an efficient counter-defence to circumvent plant antiviral immunity [67]. Consistent with this notion, viruses possess a huge repertoire of proteins, which act as RNA-silencing suppressors to dampen host antiviral defences. Viral suppressors of RNA silencing (VSRs) are phylogenetically unrelated, mostly multifunctional proteins that antagonize multiple steps of the RNA-silencing pathway, including (i) impairment of viral siRNA biogenesis by inhibiting DCL proteins and/or the activity of cofactors, (ii) sequestration of dsRNA/siRNA, (iii) promotion of AGO protein destabilization prior to RISC formation or (iv) transportation of the mobile silencing signal into the peroxisomes to disable plant defence [57,68]. Additionally, some VSRs play a pivotal role in replication, assembly or movement of viruses [12]. VSRs have been identified from almost all plant virus genera [57,67]; nevertheless, the multi-functionality of these proteins and the biochemical processes in which they may be involved, such as fine-tuning the plant-virus interaction, hampers the unravelling of their exact mode of action.
To overcome RNA-silencing suppression caused by VSRs, plants have evolved specific defence mechanisms [32], supporting the hypothesis of a molecular arms race between VSRs and RNA-silencing pathway genes [69]; however, studying the diversity of VSRs and the co-evolution between plants and viruses will increase our understanding of plant molecular biology as well as biochemical and cellular activities [70]. Importantly, a lack or inactivation of VSRs leads to the recovery of plants from viral infections [40,71]. For detailed information on the role of VSRs in host-virus interactions, the readers are referred to recent reviews [57,72,73].
3. Improvement of Plant Immunity Using RNA Silencing-Based Plant Protection Strategies
Over the last decade RNA silencing has emerged as a powerful genetic tool for scientific research. In addition to fundamental research for the assessment of gene function, RNA-silencing technology has been employed to develop plants with improved resistance to biotic stresses based on their function to provide protection from invasion by foreign nucleic acids, such as viruses (reviewed by [74,75]). This natural phenomenon can be used to control agronomically relevant plant diseases, based on the demonstration that in vitro feeding of dsRNA can signal PTGS of target genes in various plant pests and pathogens [76,77,78,79]. Indeed, expression of such dsRNAs in the corresponding host plant conferred protection from predation or infection [80]. This biotechnological method, termed host-induced gene silencing (HIGS), has emerged as a promising alternative in plant protection because it combines high selectivity for the target organism with minimal side effects, as compared with chemical treatments. In previous studies, we have demonstrated that transgenic Arabidopsis and barley (Hordeum vulgare) plants, expressing a 791 nucleotide (nt) dsRNA (CYP3RNA) targeting all three CYP51 genes (FgCYP51A, FgCYP51B, FgCYP51C) in Fusarium graminearum (Fg), inhibited fungal infection via HIGS [81,82].
4. RNA Silencing-Based Crop Protection Against Viruses
Many viruses are transmitted via vectors or physical wounds, multiply rapidly and spread across the same or different plant species. Plants naturally resist against viruses using an RNA-silencing mediated defence, which is often not sufficiently effective to stop viral infection completely because siRNA molecules complementary to viral sequences usually appear at later stages of infection [84,85]. Mimicking the RNA-silencing mechanism in planta to generate siRNAs using genetic engineering and biotechnology approaches may help to induce resistance against viruses even before the onset of infection. Since RNA silencing has been shown to synergize with plant innate immunity pathways, integration of different defence layers is expected to ensure a robust defence response against plant viruses [32].
The inoculation of plants with attenuated strains of viruses or viroids that confer cross-protection against more virulent strains to reduce yield losses in cash crops, such as potato or tomato, was already recognized more than half a century ago [86]; however, the first report indicating that plants can be genetically transformed for resistance to virus disease development was published in 1986 [87]. The authors generated a chimeric gene encoding the tobacco mosaic virus (TMV) (species Tobacco mosaic virus) CP that was introduced into tobacco cells through Agrobacterium tumefaciens-mediated transformation [87]. Transgenic plants showed suppression of symptom development after infection with TMV. At that time, the underlying mechanism of transgene-induced viral resistance and cross-protection remained elusive. More than three decades later, the induction of RNA silencing by tentransgenic expression of virus-derived dsRNA in planta has been successfully implemented to control plant viral diseases.
Since the first publications in the early 1990s [88], a vast number of studies on host-derived gene silencing in plant-virus interactions have been conducted (Table 1). In 2011, there were more than 30 new reports on RNA-silencing technology for controlling a wide array of viral plant diseases. Early in 2012, studies revealed the efficiency of RNA silencing to control viral diseases, e.g., in: Solanum tuberosum [89], Cucumis melo [90], grapevine [91], banana [92] and rice [93]. In this review, we present an overview on RNA silencing mediated control of plant viral disease in transgenic crop plants. In the following sections, we will highlight a few studies representative for the significant progress that was achieved in this area, and we will discuss transgenic virus resistance of various crop plants for major plant families, such as Solanaceae, Cucurbitaceae, Fabaceae, Poaceae, Euphorbiaceae and tropical fruits. Due to the vast number of publications reflecting the rapid progress of RNA silencing and its great relevance for agriculture, it is not possible to cover all studies in the following sections. Nevertheless, a complete overview is given in Table 1.
5. RNA Silencing-Mediated Viral Resistance in Solanaceae—Tobacco, Tomato and Potato
The economic importance of plant viruses infecting solanaceous hosts like tobacco is reflected by the vast number of HIGS studies related to the species Nicotiana benthaminana and Nicotiana tabacum (Table 1). Rigid, rod-shaped (+) ssRNA viruses of the genera Tobamoviruses and Cucumovirus cause serious agronomical losses by damaging the leaves, flowers and fruits of their hosts (including tobacco, tomato, cucumber and pepper). Diseases caused by the TMV and cucumber mosaic virus (CMV, species Cucumber mosaic virus) are found worldwide; thus, scientists used genetic engineering to introduce TMV and CMV resistance. The utility of virus-derived transgenes expressing dsRNA that mediate viral RNA silencing was demonstrated by different groups (Table 1). They showed that tobacco plants transformed with inverted repeated sequences of the partial TMV movement protein (MP) gene and the partial sequence of CMV replication protein (RP) gene exhibited PTGS of the corresponding genes [94]. The transgenic plants exhibited complete resistance to TMV or CMV infection. Moreover, the authors proved that the silencing was stably inherited through self-pollination in T4 progeny and that viral resistance was unaffected by low temperature (normally compromises siRNA–mediated gene silencing [166]) [94].
A key step in developing a successful RNA-silencing strategy is the identification of suitable target genes in the infectious agent. Whereas approximately 20% of the publications demonstrated that the replicase gene represents a prominent target, every third study chose partial- to full-length sequences of the CP gene to confer viral disease resistance (Table 1). Transgenic tobacco plants that express dsRNA homologous to the CP gene of TMV and CMV were proven to trigger RNA silencing of the corresponding viral genes [95,96,167]. Moreover, the authors found that the number of siRNAs correlates with the degree of resistance [95]. Interestingly, they showed that only 17% of the transgene-expressing plants generated substantial amounts of siRNAs that confer CMV resistance. Several years later, the same authors addressed the question whether multiple transgene copies coincided with the processing of hpRNA to siRNA and the occurrence of resistant phenotypes [167]. They found no significant correlation between the resistance and the copy number of the transgene and consistent with another study [94]; therefore, structural characteristics of the RNA-silencing construct [148,149], the locus of transgene integration [168,169] and the promotor used for transgene expression [170] as well as undesired transgene silencing provoked by using transgenic technologies [171] may account for RNA-silencing effectiveness.
However, target gene silencing is correlated with the number of siRNAs generated from a dsRNA precursor; thus, it can be used as a molecular marker to predict success in attempts to engineer virus resistance by dsRNA. Previously, it was shown that dsRNA derived from hpRNA constructs of different lengths led to divergent resistance phenotypes [98], which is consistent with our own work related to fungal gene silencing. Using barley as a cereal model, we found that dsRNA constructs targeting two FgCYP51 genes inhibited fungal growth more efficient than single constructs, although both types of dsRNAs decreased fungal infections [unpublished]. Based on these findings and combined with our recent study on the efficacy of dsRNAs with increasing length in RNA silencing of the Fusarium CYP51 genes, we anticipate that constructs of >400 bp in length were more efficient because the number of siRNAs derived from those longer constructs are higher [unpublished].
Whereas most studies used virus-derived dsRNAs or dsRNAs derived from hpRNAs processed by host RNA silencing machinery into siRNA to develop virus-resistant plants, miRNA-based approaches have been explored for engineering plant virus resistance [91,97,116,161]. The first study that investigated the possibility of modifying plant miRNA sequences to target specific viral transcripts was published in 2006 by Niu et al. [161]. They used a 273-bp backbone of the miRNA precursor pre-miRNA159a to generate artificial pre-miRNA159a containing sequences complementary to the plant viruses turnip yellow mosaic virus (TYMV, species Turnip yellow mosaic virus) and turnip mosaic virus (TuMV, species Turnip mosaic virus) [161]. They designed their amiRNAs to target the sequence of two silencing suppressors, P69 of TYMV and HC-Pro of TuMV. Transgenic Arabidopsis plants that were expressing amiR-P69159a and amiR-HC-Pro159a exhibited resistance to both viruses. This pioneering work was subsequently confirmed by two studies using amiRNAs to target the VSR 2b and 2a of CMV in tobacco [97] and tomato [116].
The authors claimed that the use of amiRNAs has several advantages: (i) high specificity, thus diminishing off-target effects, (ii) ease of amiRNA design and (iii) applicability to diverse viral pathogens. Consistent with this, the design of synthetic RNAs and amiRNAs has been reported as an effective tool for functional gene studies in plants as well as for therapeutic approaches in animal and human cells [172,173].
The use of target-specific dsRNA or amiRNA as an anti-viral agent offers unprecedented potential as a new plant protection strategy; therefore, successful field application will require optimization of an RNA-silencing construct design necessary to maximize the efficacy of the RNA-silencing-based pathogen control. Consistent with this, the RNA-silencing construct design can be directed either to combine high selectivity for the target organism with minimal side effects on beneficial microorganisms, or generating chimeric sequences to achieve broad-spectrum resistance to multiple viral pathogens.
Consistent with the idea of parallel or simultaneous silencing of different targets by using a single chimeric transgene construct, a recent study demonstrated that fused viral coat protein coding sequences from potato virus X (PVX), potato virus Y (PVY) and potato virus S (PVS) conferred simultaneous resistance against all three RNA viruses in potato [122]. The authors fused 180 bp (PVX), 240 bp (PVY) and 180 bp (PVS) fragments for a total of 600 bp sequence for developing broad-spectrum virus resistance in transgenic potato plants. Simultaneous resistance to multiple viruses was previously reported by the transgenic expression of fused-tandem, repeat, virus-derived dsRNAs [89,102,174]. However, resistance to mixed infection by two different potato viruses can be achieved without using fused-tandem constructs [175].
Although dsRNA as well as hpRNA which represent the intermediate forms during viral replication, are the key triggers of RNA-silencing machinery [176,177] and are, thus, expected to exhibit stronger virus resistance [178], earlier studies were performed using antisense RNA constructs [88,110]. For example, transgenic tobacco plants expressing an antisense RNA that compromises the complete coding sequence of the AL1 gene encoded in members of the species Tomato golden mosaic virus in the genus Geminivirus reduced symptom development and viral replication [88]. The authors argued that the AL1 gene is required for DNA replication and, thus, together with its conservation in all Geminiviridae, makes it a promising target. Moreover, the transgenic cassette compromised the 5′ sequences of two other open-reading frames, AL2 and AL3, resulting in a 1258-bp fragment. The authors found that the lines expressing the most antisense RNAs exhibited the highest resistance. Importantly, the copy number of the integrated DNA did not correlate with the level of resistance or antisense RNA [88]; however, the percentage of plants that showed a reduction in symptom development after inoculation with TGMV differed between lines transformed with the same construct. Thus, this, together with the results from other studies (reviewed above), points to the variation in gene expression that can be explained, for example, by the transcriptional activity of the region surrounding the integrated transgene.
6. RNA-Silencing-Mediated Viral Resistance in Cucurbitaceae—Cucumber, Melon and Watermelon
Cucurbit crops, such as melon, cucumber, squash/pumpkin and watermelon, are susceptible to at least 35 viruses [179] that can cause massive damage up to total loss [180]. Improvement of viral resistance relies mainly on biotechnological approaches, such as genetic engineering.
Previously, we have demonstrated that transgenic Arabidopsis and barley (Hordeum vulgare) plants, expressing a 791 nt dsRNA (CYP3RNA) that targets all three CYP51 genes (FgCYP51A, FgCYP51B, FgCYP51C) in Fusarium graminearum (Fg), inhibited fungal infection via HIGS [82]. Concurrently, the HIGS technology enables us to generate constructs that are highly specific for the targeted genes, preventing side effects on other (beneficial) microbes and host plants; however, the homology of CYP3RNA to the CYP51 genes raise the possibility that this HIGS strategy can be used to control a wide range of fungal pathogens. Consistent with this idea, the silencing of a single-target gene can confer broad-spectrum and durable resistance to multiple viral pathogens of the cucurbitacous host melon (Cucumis melo L.) [90].
Virus resistance can be achieved through the absence of host factors, known as susceptibility factors, that are required for the virus to complete its biological cycle [181]. Moreover, it was shown that different viruses have common susceptibility factors, thus indicating that RNA-mediated silencing of those host factors could confer broad-spectrum disease resistance. In melon, the eukaryotic translation initiation factor Cm-eIF4E represents such a common susceptibility factor [181]. In 2012, Rodríguez-Hernández et al. [90] generated transgenic melon plants expressing a hpRNA that induced silencing of Cm-eIF4E. The authors hypothesized that Cm-eIF4E may control the susceptibility of a broad range of viruses; therefore, they challenged these transgenic melon plants with eight agronomically important melon-infecting viruses and identified that they were resistant to cucumber vein yellowing virus (CVYV), melon necrotic spot virus (MNSV), Moroccan watermelon mosaic virus (MWMV) and zucchini yellow mosaic virus (ZYMV) [90]. A previous report has shown that silencing of the VSR HC-Pro from ZYMV conferred resistance not only to ZYMV but also to the papaya ringspot virus and watermelon mosaic virus (WMV) in cucumber and melon [126]. This finding is based on co-silencing effects related to sequence homologies between the HC-Pro genes of those three viruses. Recently we found that fungal resistance induced by CYP51-dsRNAs (targeting the fungal ergosterol biosynthesis) in Arabidopsis mirrors the co-silencing of non-target FgCYP51 genes [unpublished]. Overall, these results suggest that resistance conferred by single dsRNA constructs can be mediated by co-silencing effects on non-targeted genes of a wide range of pathogens.
However, multiple-virus resistance in transgenic plants can also be achieved by using chimeric transgene constructs composed of viral gene segments that induce PTGS without the expression of dsRNA trigger molecules [127,182,183,184,185,186]. The underlying mechanism is based on transgene silencing in a length-dependent manner that is independent of the ability to provide homology-dependent trans-inactivation of a homologous, incoming virus [187]. The authors found that the expression of large segments (387–453 bp) of the nucleocapsid (N) encoding gene of the tospovirus were silenced in transgenic tobacco plants mediating tomato spotted wilt virus (TSWV) resistance through PTGS. Importantly, small N gene segments (92–235 bp) were ineffective in conferring resistance unless they were transcriptionally fused to the non-target GFP gene DNA. The authors argued that the inability of a small N transgene alone to induce homology-dependent virus resistance was because they are incapable of inducing gene silencing [187]. Importantly, the same authors provided the hint that any viral sequence with a minimum length of 59–110 bp could confer RNA silencing-mediated resistance when fused to a transcribed DNA, designated as a silencer DNA (in this case a green fluorescent protein (GFP) gene) [182]. However, in a subsequent study they replaced the silencer GFP DNA with the full-length CP gene of turnip mosaic virus (TuMV) linked to 218 or 110 bp N gene segments for the transformation into Nicotiana benthamiana [188]. Transgenic lines with the 218 bp N gene segment linked to the TuMV CP gene exhibited TSWV resistance. More than one decade later, they transferred their strategy for engineering multiple-virus resistance in transgenic watermelon [127] and generated a chimeric transgene consisting of three viral CP gene segments of CMV, cucumber green mottle mosaic virus (CGMMV) and WMV fused together and cloned into a plant transformation vector carrying a silencer DNA comprising the middle half N gene of watermelon silver mottle virus (WSMoV), resulting in a transgene length of 1.7 kb. The authors identified two transgenic lines (R0 Lines 6 and 14) that showed resistance to all three viruses individually and mixed infection [127]. Together, these studies showed that PTGS is the underlying mechanism for the multiple-virus resistance that can be triggered without the expression and formation of transgene-derived dsRNAs.
7. RNA-Silencing-Mediated Viral Resistance in Fabaceae—Soybean
Conventional soybean protection measures based on managing virus vectors were less efficient [189]; therefore, RNA-silencing technology represents a promising alternative. A recent study demonstrated that RNA-mediated silencing of the soybean mosaic virus (SMV) P3 cistron induced resistance to five different SMV strains, the soybean-infecting bean common mosaic virus (BCMV) and the WMV in transgenic soybean [133]. Soybean mosaic virus belongs to the virus family Potyviridae of which the genus Potyvirus contains the most numerous and most important plant viruses. SMV is the most devastating viral pathogen in soybean and can cause severe yield losses [190]. The P3 protein is involved in virus replication, movement, pathogenesis and SMV virulence [191,192,193] and, thus, represents a suitable target for RNA-silencing-mediated approaches [133]. The P3 cistron-silencing mediated by a 302-bp inverted repeat (IR) provoked significantly enhanced SMV resistance under field conditions over three years. However, SMV resistance can also be achieved by expression of viral genes and sequences in transgenic soybeans [194,195,196,197].
Further studies reported successful engineering of viral resistance in soybean using RNA-silencing strategies [128,129,130,131,132,198]; thus, SMV resistance was achieved by silencing the VSR HC-Pro [130,132] and the CP gene [199]. Multiple-virus resistance was conferred by expressing a transgene designed to express several shorter IRs [129]. The authors chose specific, highly conserved sequences of less than 150 bp derived from three viruses, alfalfa mosaic virus (AMV), bean pod mottle virus (BPMV) and SMV to assemble the short IRs. Transgenic lines exhibited simultaneous resistance to these viruses, blocking systemic spread and eliminating seed mottling. The authors argued that their strategy of construct assembly makes it easy to incorporate additional IR sequences of viral origin, thus, conferring resistance to a wide range of soybean-infecting viruses [129]. The same authors conducted another study to compare virus-resistant transgenic soybeans with their non-transgenic counterparts to verify whether biologically important changes occurred due to IR insertion [200]. The authors followed the concept of “substantial equivalence” developed by the Organisation for Economic Co-operation and Development (OECD) in 1993 [201] and further elaborated by FAO/WHO [199,200]. Their substantial equivalent analysis comprised: (i) proximate analyses, including moisture, ash content, crude protein, crude fat and carbohydrates; (ii) the content of 18 amino acids; (iii) fatty acid composition; (iv) the analyses of two important antinutritional factors (lectin and trypsin inhibitors); (v) the contents of isoflavones, including daidzin, glycitin, genistin, daidzein, glycitein and genistein, of transgenic soybeans and non-transgenic counterpart seeds. Their results showed that the RNA-silencing-mediated virus-resistant transgenic soybeans are as safe and nutritious as their traditional counterparts [200]. The authors claimed their findings can serve as baseline information for future generations of RNA-silencing-mediated virus-resistant transgenic crops.
To use the sense CP gene to direct RNA silencing of viral target genes can induce PTGS, although the complementarity to RNA transcripts is not given. Moreover, most studies were using sense gene sequences as transgenic controls (Table 1). Interestingly, transgenic soybeans expressing the sense CP gene of the soybean dwarf virus (SbDV) exhibited SbDV-CP-specific siRNAs, and those plants remained symptomless after SbDV inoculation [198]; however, the underlying mechanisms remain elusive.
8. RNA-Silencing-Mediated Viral Resistance in Poaceae—Rice
RNA-silencing-mediated biotechnological approaches to generate genetically engineered cereals, such as maize, rice, wheat and barley, represent a promising tool to improve plant immunity towards viral infections. RNA-silencing-based plant protection strategies provide a promising alternative to genetic resistance approaches, especially if there are no naturally occurring genes that confer virus resistance to e.g., rice dwarf virus (RDV), a dsRNA virus of the family Reoviridae which causes severe disease in rice crops. Symptoms of RDV include stunting of plant growth and white chlorotic spots on leaves, leading to decreased grain yields of their hosts. RDV is transmitted exclusively by leafhoppers (Nephotettix spp.). The genome of RDV consist of 12 segments of dsRNA, designated S1-S12 that encode seven structural and five non-structural proteins [202]. Shimizu and co-authors showed in 2009 [138] that RNA silencing of the non-structural RDV protein Pns12 confers resistance to RDV by insertion of a IR construct that led to the formation of a 500-bp Pns12-complementary dsRNA in transgenic rice plants [138]. Moreover, they generated transgenic rice plants carrying an IR construct specific to the Pns4 non-structural protein to compare resistance phenotypes of both RNA-silencing targets. Whereas Pns12-dsRNA expressing plants exhibited complete RDV resistance, Pns4-dsRNA plants displayed incomplete resistance. The authors argued that targeting Pns12, which plays a crucial role in viral replication at an early stage of infection [203], was more effective for controlling RDV than targeting the phosphoprotein Pns4, which is localized around the viroplasm matrix, forming minitubules, and is expressed at a relatively late stage of viral infection [204]. Importantly, the same authors showed that targeting of viroplasm matrix proteins represents an applicable strategy to control viruses of the family Reoviridae [93,141]. In the case of the genus Fijivirus, RNA silencing of viroplasm protein P9-1 of rice black streaked dwarf virus (RBSDV) induced virus resistance in rice [141]. In another study, they showed that IR-derived dsRNA that corresponds to the non-structural viroplasm protein Pns9 of rice gall dwarf virus (RGDV) confers strong resistance to RGDV infection in transgenic rice plants [93]. Consistent with their study published in 2009, Shimizu and co-authors further demonstrated that the identification of suitable potent viral target genes is essential to guarantee strong resistance effects mediated by RNA silencing-based plant protection approaches, as not all RNA-silencing constructs exhibit equal effectiveness in preventing virus infection [140]. They attempted to develop rice stripe virus (RSV) resistance by targeting seven different RSV proteins (pC1-4 and p2-4). The targets pC3 and pC4, which encode a nucleocapsid protein and a movement protein of RSV, respectively, were more resistant to RSV infection compared with RSV-resistant rice cultivars cv. Musashikogane and Sainokagayaki. Similar results were obtained using RNA silencing of NP pC5 gene and MP pC6 gene of rice grassy stunt virus (RGSV) that displayed strong RGSV resistance indicating that NP and MP represent promising targets to control viral disease caused by members of the genus Tenuivirus [142]. Targeting the pC1 gene, which encodes a putative RNA-dependent RNA polymerase, also conferred high level resistance to RSV compared with susceptible wild-type rice [143]. Transgenic rice plants that harbour p2-(protein of unknown function) and p3 (VSR)-specific RNA-silencing constructs exhibited moderate resistance to RSV infection. By contrast, the IR constructs that correspond to the pC2 gene (encodes a glycoprotein of unknown function) as well as the p4 gene (encodes a non-structural protein of unknown function) failed to induce enhanced disease resistance, underlining that not all RNA-silencing constructs establish equal resistance phenotypes [140].
9. RNA-Silencing-Mediated Viral Resistance in Poaceae—Wheat and Barley
Wheat streak mosaic virus (WSMV) (genus Tritimoviruses; family Potyviridae) causes yellowish-orange streaks on leaves of their hosts, which are restricted to the Gramineae family, and is transmitted by eriophyid mites. WSMV is one of the most important destructive viral diseases of wheat causing severe yield losses [205,206]. There are two reports demonstrating that RNA-silencing-mediated virus control represents a promising alternative to natural resistance strategies [143,144]. The authors selected the nuclear inclusion protein NIa gene of WSMV as an RNA-silencing target. Notably, transgenic wheat plants harbouring a single insertion of the Nia transgene were immune to WSMV infection [143,144].
Another study in barley used the RNA-silencing strategy to control barley yellow dwarf virus (BYDV) [146], which occurs throughout the world and affects a wide range of gramineous hosts. Yield losses caused by the impairment of tillering, flowering and kernel production range from 5%–30% in wheat and barley in years of BYDV outbreaks [207]. Barley plants can be protected from BYDV infection by transformation with hpRNA constructs expressing dsRNA complementary to the polymerase gene of BYDV [146]. Notably, BYDV-PAV immunity was not affected by cereal yellow dwarf virus-RPV co-infection based on the finding that some viruses have the capacity to enhance replication and spread of the co-infecting viruses and to inactivate PTGS [208]. The authors presumed that using hpRNA technology to generate transgenic BYDV-immune barley plants might have great potential for field trials [146].
10. RNA-Silencing-Mediated VIRAL Resistance in Euphorbiacea—Cassava
Cassava production in sub-Saharan Africa is limited by two devastating viral diseases: Cassava mosaic disease (CMD) and cassava brown streak disease (CBSD). CMD is widespread over the East African countries, where 150 million people depend on the tropical staple cassava as their largest source for carbohydrates [209]. CMD is caused by African cassava mosaic virus (ACMV), which can result in severe losses of approximately 20%–30% of the annual production [209]. RNA-silencing technology has been extensively used to bring effective solutions to viral diseases that diminish cassava yields and reduce farmer incomes in East Africa. Towards this, a collaborative program named the Virus Resistant Cassava for Africa (VIRCA) project was founded in 2005 [209]. The VIRCA strategies rely on RNA-silencing-mediated viral disease control through transgenic expression of RNA-silencing triggers. Consistent with this, there are several studies demonstrating CMD and CBSD resistance using RNA-silencing technology (Table 1).
For example, CBSD resistance was achieved by targeting the full-length CP of cassava brown streak Uganda virus (CBSUV) through transgenic expression of RNA silencing constructs in cassava [153]. Previously, the authors tested three RNA-silencing constructs consisting of full-length (894 bp), N-terminal (397 bp) and C-terminal (491 bp) portions of the CP gene of CBSUV in transgenic Nicotiana benthamiana plants and identified FL-CP as the most efficient construct [105]. However, the same authors underlined the significance of their findings by testing seven FL-CP-siRNA-producing lines under conditions of naturally vectored disease pressure under confined field trials [154]. They demonstrated that field-grown transgenic cassava plants remained symptom-free over the trial period of 11 months, underlining the great potential RNA-silencing technology holds for reaching goals of the VIRCA project. Furthermore, they confirmed RNA-silencing-mediated CBSD resistance through a typical vegetative propagation cycle [155]. Their studies provided significant progress towards the development and delivery of CBSD-resistant planting materials for farmers’ use in East Africa. Additionally, Vanderschuren et al. [152] strengthened the potential of RNA-silencing technology to control CBSV in the West African cultivar TME7. Recently, Beyene et al. (2017) [156] genetically modified the Ugandan farmer-preferred cassava cultivar, TME 204, with an improved RNA-silencing construct, consisting of the CP sequences of Ugandan cassava brown streak virus (UCBSV) and CBSV fused in tandem. The resulting transgenic lines exhibited high levels of resistance to both viruses [156] under subsequent field trials in Uganda and Kenya [151].
ACMV, as one of the causal agents of CMD, was also a focus of the VIRCA project [209]. Pioneering work towards RNA-silencing-mediated generation of CMD-resistant cassava was performed in collaboration between the labs of Wilhelm Gruissem (Switzerland) and Peng Zhang (China) [147,148,149,152]. First, they used antisense RNAs against viral mRNAs, encoding essential non-structural proteins required for viral replication and transcriptional regulation of ACMV, such as Rep (AC1), TrAP (AC2) and Ren (AC3) [147]. Towards this, they inserted full-length sequences of those target genes in antisense orientation, engineering ACMV-resistant transgenic cassava, with antisense-AC3 lines exhibiting the highest level of resistance. However, the use of sense and antisense RNA strategies produced lines showing large variations in resistance levels under high viral infection pressure [147,210]. The authors argued that using hp-derived dsRNA approaches are more efficient based on siRNA quantities. Consistent with this idea, they observed a dose-dependent sRNA-mediated ACMV resistance in transgenic cassava expressing a short (155 bp) hp construct [149]. Whereas other RNA-silencing-based strategies rely on the use of long introns and hp arms longer than 400 bp to produce intron hp-derived sRNA (Table 1), they provided a proof-of-concept that short introns and short hp arms combined with a strong promotor (p35s) might represent the most efficient strategy to reach the highest levels of target-specific sRNAs [149]. Notably, they found a substantial quantity of sRNA derived from the base-paired region of the hp that were longer compared to the known 21–24 nt RNA-silencing-related sRNAs. Whether these long sRNA contribute to target gene silencing and virus resistance remain unknown.
Interestingly, in 2007, they expanded their former RNA-silencing strategies in controlling DNA viruses of the family Geminiviridae through expression of dsRNA homologous to viral non-coding sequences [148]. They designed an RNA-silencing construct that consisted of the bidirectional promotor and the common region sequence of ACMV. Although they identified ACMV-resistant lines containing siRNAs derived from non-coding sequences, they concluded that transformation efficiencies of transgenic cassava plants harbouring siRNAs originating from ACMV-dsRNA targeting viral coding sequences were much higher [148]. Whether the low plant regeneration frequency was due to the transgene they transformed remained speculative; however, their results principally suggest that RNA-silencing technology can be expanded to target the entire genome of a DNA virus to confer disease resistance.
11. RNA-Silencing-Mediated Viral Resistance in Fruits—Citrus
The viral disease of citrus represents a serious threat to farmers in all citrus-growing areas of the world. Citrus tristeza virus (CTV) can cause severe losses of fruit quantity and quality of its hosts, primarily orange, grapefruit and lime [211], along with the death of the tree. The group of Leandro Peña in Spain provided huge progress in engineering CTV-resistant Mexican lime [158,159,212]. Their RNA-silencing strategies rely on the silencing of CTV-VSR based on their former work where they identified p23 as an important CTV pathogenicity determinant through its ectopic expression that induced viral-like symptoms [213,214]. Surprisingly, five of the p23-transgenic lines displayed characteristics of PTGS and TGS, such as the accumulation of p23-specific siRNAs, low levels of the corresponding mRNA as well as transgene silencing resulting in a CTV-immune phenotype [212]. Based on this finding that Mexican lime plants transformed with a silenced p23 transgene showed resistance to CTV infection, they overexpressed the 3′-terminal 549 nt region of the p23 gene together with the 3′UTR in sense, antisense and hp formats to generate RNA-silencing-mediated CTV resistance [158]. Unexpectedly, they found that transgene-derived siRNA levels varied in different transformants irrespective of their response to CTV, indicating that high abundancies of transgene-derived siRNA are not necessarily correlated with CTV resistance [158]. The authors suggest that this lack of correlation is presumably specific for RNA viruses, further arguing that there is a siRNA threshold below which the virus can overcome resistance, which is consistent with previous reports [215,216]. However, in a subsequent study, they extended their RNA-silencing approach and transformed Mexican lime with an intron hp vector carrying full-length sequences of two more CTV-VSR, p20 and p25, providing proof-of-concept that simultaneous targeting of three VSRs confer transgenic resistance to CTV [159].
Citrus psorosis virus (CPsV), an (−) ssRNA virus, is another causal agent of a serious disease affecting citrus. The group of María Laura García in Argentina developed CPsV-resistant transgenic orange plants via RNA silencing of the CP gene [160]. Notably, they obtained similar results as the group of Leandro Peña working on CTV-resistance in which the level of accumulated siRNAs did not correlate with the degree of virus resistance among different lines/transformants. Those results further support the notion that virus resistance cannot be predicted only by the presence of high amounts of transgene-derived siRNAs, whether this is a general aspect of using RNA-silencing technology or something specific to RNA silencing pathways in citrus trees remains to be elucidated.
12. Outlook: Non-GMO RNA Spray Confer Virus Resistance
While all the reviewed studies provided proof-of-concept that RNA-silencing-based plant protection is an effective strategy for controlling diseases caused by viral pathogens, the broad applicability of HIGS remains questionable due to fact that generation of genetically modified (GM) crops is time-consuming and still poorly accepted in many European countries. Moreover, it relies on the transformability of a plant, thus, limiting or making it an exclusive approach for transformable crop plants only. Whereas a great number of studies have been published on HIGS-mediated silencing of target genes in pathogenic viruses (Table 1), silencing of such targets through exogenously applied dsRNA has been described in only a few studies [217,218,219,220,221]. The induction of virus resistance by exogenous application of dsRNA has been reviewed recently [for details, see 188]. Therefore, a breakthrough was achieved by showing that inhibitory dsRNA is effective upon spray application. Recently, we have shown that spray applications of CYP3RNA also protect barley from fungal infection via a process termed ‘spray-induced gene silencing’ (SIGS) [222]. In the same year, Konakalla et al. [223] showed that exogenous application of dsRNA targeting the VSR p126 gene and CP gene of TMV conferred virus resistance in tobacco [223]. Importantly, authors confirmed the spreading of dsRNA from local to systemic tissue by one hour after rub-inoculation of dsRNA using semi-quantitative RT-PCR. Moreover, they showed that p126 dsRNA levels continuously decreased in the local (treated) tissue from 3 dpi to 9 dpi where dsRNA was no longer detectable. This is consistent with our own work, which showed movement of sprayed dsRNA from barley leaves over stems to the root tissue within three days after spray treatment (unpublished data). In 2018, Kaldis et al. [224] showed that exogenously applied dsRNA derived from the HC-Pro and CP genes of ZYMV protect watermelon and cucumber against ZYMV and spread systemically over long distances in cucurbits.
Given the significance of these recent findings, subsequent studies were conducted to improve the stability of sprayed dsRNA and to find a system that allows large-scale production of long dsRNA molecules suitable for application in crop protection [225,226]. To provide long-term protection against the targeted viral pathogen the uptake of sprayed dsRNA into plant cells is a critical step together with improving its stability to overcome environmental degradation by UV radiation or simply surface wash-off by rain. Towards this, Mitter et al. [159] used layered double hydroxide (LDH) clay nanosheets as a dsRNA carrier, a technology originally developed for the delivery of siRNA therapeutics to mammalian cells [227,228]. The authors demonstrated that loading dsRNA on LDH prolonged their durability on the leaf surface for 30 days and increased their stability through protection from nuclease degradation [229]. Moreover, they showed uptake of dsRNA into plant cells and induction of endogenous RNA silencing that mediated systemic protection against the targeted VSR 2b gene of CMV inoculated on cowpea and tobacco [225]. Notably, they proved that the LDH nanocarrier can be completely degraded over time, thus, resulting in slow and sustained release of dsRNA under environmental conditions. However, whereas this study provides a significant step forward in making RNA spray an applicable and sustainable approach towards pathogen and pest control in agriculture, the question is how to produce efficient amounts of dsRNA for spray applications in field trials. Towards this, Manfred Heinlein and his group in France established a dsRNA production system that enables the broad application of dsRNA molecules as a non-GMO crop protection measure [226]. They used the RNA-dependent RNA polymerase of the dsRNA phage phi6 for in vivo synthesis of dsRNA molecules. Towards this, they engineered a phi6-based dsRNA-synthesizing machine within the bacterium Pseudomonas syringae. The authors argued exploiting phi6 that naturally converts ssRNA templates into dsRNA would produce sufficient amounts of dsRNA with higher quality and purity compared to dsRNA generation e.g., by enzymatically synthesized ssRNA strands in vitro and their subsequent physical annealing to dsRNA. Indeed, they showed that application of purified in vivo-produced dsRNA to target either the GFP gene or the viral replicase gene of a GFP-tagged TMV strain provided efficient protection against local and systemic spread of TMV after virus inoculation [226]. Given the significance of their results, this opened the door for large-scale vaccination applications of long dsRNAs to protect crop plants against several pathogens and pests simultaneously. However, cost-effective production methods of dsRNA for commercialisation of exogenous dsRNA application in plant virus control was published previously (for more information, see [229]).
The reviewed studies are of a ground-breaking nature, as they represent significant progress towards establishing an RNA-silencing-based non-GMO crop protection approach using direct-spray applications of dsRNA to target viral pathogens. However, further research is needed to (i) elucidate the mechanistic basis of entry, transport and procession, (ii) solve problems of durability and instability and (iii) implement the large-scale production and purification of exogenously applied dsRNA to make RNA-silencing-based spray approaches for plant protection scientifically and economically achievable.
Acknowledgments
This work was supported by the Deutsche Forschungsgemeinschaft, Research Training Group (RTG) 2355 (project number 325443116) to AK and Deutscher Akademischer Austauschdienst to FYG.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Hull, R. Matthews’ Plant Virology, 4th ed.; Academic Press: San Diego, CA, USA, 2002. [Google Scholar]
- Dall’Ara, M.; Ratti, C.; Bouzoubaa, S.E.; Gilmer, D. Ins and Outs of Multipartite Positive-Strand RNA Plant Viruses: Packaging versus Systemic Spread. Viruses 2016, 8, 228. [Google Scholar] [CrossRef] [PubMed]
- Hull, R. Plant Virology, 5th ed.; Academic Press: Amsterdam, The Netherlands, 2013; p. 1118. [Google Scholar]
- Andika, I.B.; Kondo, H.; Sun, L. Interplays between Soil-Borne Plant Viruses and RNA Silencing-Mediated Antiviral Defense in Roots. Front. Microbiol. 2016, 7, 1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, H.E.; Holmes, E.C.; Gildow, F.E.; Bothe-Goralczyk, M.A.; Stephenson, A.G. Experimental Verification of Seed Transmission of zucchini yellow mosaic virus. Plant Dis. 2011, 95, 751–754. [Google Scholar] [CrossRef] [PubMed]
- Simmons, H.E.; Dunham, J.P.; Zinn, K.E.; Munkvold, G.P.; Holmes, E.C.; Stephenson, A.G. Zucchini yellow mosaic virus (ZYMV, Potyvirus): Vertical transmission, seed infection and cryptic infections. Virus Res. 2013, 176, 259–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitzalis, N.; Heinlein, M. The roles of membranes and associated cytoskeleton in plant virus replication and cell-to-cell movement. J. Exp. Bot. 2017, 18, 117–132. [Google Scholar] [CrossRef]
- Zhou, J.S.; Drucker, M.; Ng, J.C. Direct and indirect influences of virus-insect vector-plant interactions on non-circulative, semi-persistent virus transmission. Curr. Opin. Virol. 2018, 33, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Gallet, R.; Michalakis, Y.; Blanc, S. Vector-transmission of plant viruses and constraints imposed by virus-vector interactions. Curr. Opin. Virol. 2018, 33, 144–150. [Google Scholar] [CrossRef]
- Gouveia, B.C.; Calil, I.P.; Machado, J.P.; Santos, A.A.; Fontes, E.P. Immune Receptors and Co-receptors in Antiviral Innate Immunity in Plants. Front. Microbiol. 2017, 5, 2139. [Google Scholar] [CrossRef]
- Carr, J.P.; Murphy, A.M.; Tungadi, T.; Yoon, J.-Y. Plant defense signals: Players and pawns in plant-virus-vector interactions. Plant Sci. 2019, 279, 87–95. [Google Scholar] [CrossRef]
- Moon, J.Y.; Park, J.M. Cross-Talk in Viral Defense Signaling in Plants. Front. Microbiol. 2016, 7, 904. [Google Scholar] [CrossRef]
- Wu, X.; Valli, A.; García, J.A.; Zhou, X.; Cheng, X. The Tug-of-War between Plants and Viruses: Great Progress and Many Remaining Questions. Viruses 2019, 11, 203. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Garcia, S.; Voinnet, O. Nonsense-Mediated Decay Serves as a General Viral Restriction Mechanism in Plants. Cell Host Microbe 2014, 16, 391–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szádeczky-Kardoss, I.; Gál, L.; Auber, A.; Taller, J.; Silhavy, D. The No-go decay system degrades plant mRNAs that contain a long A-stretch in the coding region. Plant Sci. 2018, 275, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Szádeczky-Kardoss, I.; Csorba, T.; Auber, A.; Schamberger, A.; Nyikó, T.; Taller, J.; Orbán, T.I.; Burgyán, J.; Silhavy, D. The nonstop decay and the RNA silencing systems operate cooperatively in plants. Nucleic Acids Res. 2018, 46, 4632–4648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 16, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Early molecular events in PAMP-triggered immunity. Curr. Opin. Plant Boil. 2009, 12, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef]
- Tang, D.; Wang, G.; Zhou, J.-M. Receptor Kinases in Plant-Pathogen Interactions: More Than Pattern Recognition. Plant Cell 2017, 29, 618–637. [Google Scholar] [CrossRef] [Green Version]
- Weiberg, A.; Wang, M.; Lin, F.M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.D.; Jin, H. Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 2013, 4, 118–123. [Google Scholar] [CrossRef]
- Weiberg, A.; Bellinger, M.; Jin, H. Conversations between kingdoms: Small RNAs. Curr. Opin. Biotechnol. 2015, 32, 207–215. [Google Scholar] [CrossRef]
- Weiberg, A.; Jin, H. Small RNAs--the secret agents in the plant-pathogen interactions. Curr. Opin. Plant Biol. 2015, 26, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-triggered immunity: From pathogen perception to robust defense. Annu. Rev. Plant Biol. 2015, 66, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Kapos, P.; Zhang, Y. NLRs in plants. Curr. Opin. Immunol. 2015, 32, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Kapos, P.; Devendrakumar, K.T.; Li, X. Plant NLRs: From discovery to application. Plant Sci. 2019, 279, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Allan, A.C.; Lapidot, M.; Culver, J.N.; Fluhr, R. An early tobacco mosaic virus-induced oxidative burst in tobacco indicates extracellular perception of the virus coat protein. Plant Physiol. 2001, 126, 97–108. [Google Scholar] [CrossRef]
- Yang, H.; Gou, X.P.; He, K.; Xi, D.H.; Du, J.B.; Lin, H.H.; Li, J. BAK1 and BKK1 in Arabidopsis thaliana confer reduced susceptibility to turnip crinkle virus. Eur. J. Plant Pathol. 2010, 127, 149–156. [Google Scholar] [CrossRef]
- Kørner, C.J.; Klauser, D.; Niehl, A.; Domínguez-Ferreras, A.; Chinchilla, D.; Boller, T.; Heinlein, M.; Hann, D.R. The immunity regulator BAK1 contributes to resistance against diverse RNA viruses. Mol. Plant Microbe Interact. 2013, 26, 1271–1280. [Google Scholar] [CrossRef]
- Niehl, A.; Wyrsch, I.; Boller, T.; Heinlein, M. Double-stranded RNAs induce a pattern-triggered immune signaling pathway in plants. N. Phytol. 2016, 211, 1008–1019. [Google Scholar] [CrossRef]
- Perraki, A.; Gronnier, J.; Gouguet, P.; Boudsocq, M.; Deroubaix, A.F.; Simon, V.; German-Retana, S.; Legrand, A.; Habenstein, B.; Zipfel, C.; et al. REM1.3’s phospho-status defines its plasma membrane nanodomain organization and activity in restricting PVX cell-to-cell movement. PLoS Pathog. 2018, 12, e1007378. [Google Scholar] [CrossRef]
- Calil, I.P.; Fontes, E.P.B. Plant immunity against viruses: Antiviral immune receptors in focus. Ann. Bot. 2017, 119, 711–723. [Google Scholar] [CrossRef]
- Ramesh, S.V.; Sahu, P.P.; Prasad, M.; Praveen, S.; Pappu, H.R. Geminiviruses and Plant Hosts: A Closer Examination of the Molecular Arms Race. Viruses 2017, 9, 256. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H.; Béclin, C.; Fagard, M. Post-transcriptional gene silencing in plants. J. Cell Sci. 2001, 114 Pt 17, 3083–3091. [Google Scholar] [PubMed]
- Vaucheret, H.; Fagard, M. Transcriptional gene silencing in plants: Targets, inducers and regulators. Trends Genet. 2001, 17, 29–35. [Google Scholar] [CrossRef]
- Castel, S.E.; Martienssen, R.A. RNA interference in the nucleus: Roles for small RNAs in transcription, epigenetics and beyond. Nat. Rev. Genet. 2013, 14, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Chekanova, J.A. Small RNAs: Essential regulators of gene expression and defenses against environmental stresses in plants. Wiley Interdiscip. Rev. RNA 2016, 7, 356–381. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ferrer, V.; Voinnet, O. Roles of plant small RNAs in biotic stress responses. Annu. Rev. Plant Biol. 2009, 60, 485–510. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.J.; Baulcombe, D.C. A species of small antisense RNA in posttranscriptional gene silencing in plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 16, 356–363. [Google Scholar] [CrossRef]
- Ketting, R.F. The many faces of RNAi. Dev. Cell 2011, 20, 148–161. [Google Scholar] [CrossRef]
- Vaucheret, H.; Vazquez, F.; Crété, P.; Bartel, D.P. The action of ARGONAUTE1 in the miRNA pathway and its regulation by the miRNA pathway are crucial for plant development. Genes Dev. 2004, 18, 1187–1197. [Google Scholar] [CrossRef]
- Brodersen, P.; Voinnet, O. The diversity of RNA silencing pathways in plants. Trends Genet. 2006, 22, 268–280. [Google Scholar] [CrossRef]
- Ghildiyal, M.; Zamore, P.D. Small silencing RNAs: An expanding universe. Nat. Rev. Genet. 2009, 10, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Paroo, Z. Biochemical principles of small RNA pathways. Annu. Rev. Biochem. 2010, 79, 295–319. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H. Post-transcriptional small RNA pathways in plants: Mechanisms and regulations. Genes Dev. 2006, 20, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.W.; Voinnet, O. Antiviral immunity directed by small RNAs. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Burgess, D.J. Small RNAs: Antiviral RNAi in mammals. Nat. Rev. Genet. 2013, 14, 821. [Google Scholar] [CrossRef] [PubMed]
- Cullen, B.R.; Cherry, S.; Tenoever, B.R. Is RNA interference a physiologically relevant innate antiviral immune response in mammals? Cell Host Microbe 2013, 14, 374–378. [Google Scholar] [CrossRef]
- Li, Y.; Lu, J.; Han, Y.; Fan, X.; Ding, S.W. RNA interference functions as an antiviral immunity mechanism in mammals. Science 2013, 342, 231–234. [Google Scholar] [CrossRef]
- Maillard, P.V.; Ciaudo, C.; Marchais, A.; Li, Y.; Jay, F.; Ding, S.W.; Voinnet, O. Antiviral RNA interference in mammalian cells. Science 2013, 342, 235–238. [Google Scholar] [CrossRef]
- Parent, J.S.; Bouteiller, N.; Elmayan, T.; Vaucheret, H. Respective contributions of Arabidopsis DCL2 and DCL4 to RNA silencing. Plant J. 2015, 81, 223–232. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, Z.; Li, Y.; Wu, J. Biogenesis, Function, and Applications of Virus-Derived Small RNAs in Plants. Front. Microbiol. 2015, 9, 1237. [Google Scholar] [CrossRef] [PubMed]
- Deleris, A.; Gallego-Bartolome, J.; Bao, J.; Kasschau, K.D.; Carrington, J.C.; Voinnet, O. Hierarchical action and inhibition of plant Dicer-like proteins in antiviral defense. Science 2006, 313, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Ye, X.; Morris, T.J. Arabidopsis DRB4, AGO1, AGO7, and RDR6 participate in a DCL4-initiated antiviral RNA silencing pathway negatively regulated by DCL1. Proc. Natl. Acad. Sci. USA 2008, 105, 14732–14737. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H.; Takeda, A.; Chapman, E.J.; Sullivan, C.M.; Fahlgren, N.; Brempelis, K.J.; Carrington, J.C. Arabidopsis RNA-dependent RNA polymerases and dicer-like proteins in antiviral defense and small interfering RNA biogenesis during Turnip Mosaic Virus infection. Plant Cell 2010, 22, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Csorba, T.; Kontra, L.; Burgyán, J. viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479–480, 85–103. [Google Scholar] [CrossRef]
- Carbonell, A.; Carrington, J.C. Antiviral roles of plant Argonautes. Curr. Opin. Plant Biol. 2015, 27, 111–117. [Google Scholar] [CrossRef]
- Wang, X.B.; Wu, Q.; Ito, T.; Cillo, F.; Li, W.X.; Chen, X.; Yu, J.L.; Ding, S.W. RNAi-mediated viral immunity requires amplification of virus-derived siRNAs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 484–489. [Google Scholar] [CrossRef]
- Garcia, D.; Garcia, S.; Pontier, D.; Marchais, A.; Renou, J.P.; Lagrange, T.; Voinnet, O. Ago hook and RNA helicase motifs underpin dual roles for SDE3 in antiviral defense and silencing of nonconserved intergenic regions. Mol. Cell 2012, 48, 109–120. [Google Scholar] [CrossRef]
- Taochy, C.; Gursanscky, N.R.; Cao, J.; Fletcher, S.J.; Dressel, U.; Mitter, N.; Tucker, M.R.; Koltunow, A.M.G.; Bowman, J.L.; Vaucheret, H.; et al. A Genetic Screen for Impaired Systemic RNAi Highlights the Crucial Role of DICER-LIKE 2. Plant Physiol. 2017, 175, 1424–1437. [Google Scholar] [CrossRef]
- Mourrain, P.; Beclin, C.; Elmayan, T.; Feuerbach, F.; Godon, C.; Morel, J.B.; Jouette, D.; Lacombe, A.M.; Nikic, S.; Picault, N.; et al. Arabidopsis SGS2 and SGS3 genes are required for posttranscriptional gene silencing and natural virus resistance. Cell 2000, 101, 533–542. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Molnar, A.; Baulcombe, D.C. Intercellular and systemic movement of RNA silencing signals. EMBO J. 2011, 30, 3553–3563. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, C.A.; Mitter, N.; Christie, M.; Smith, N.A.; Waterhouse, P.M.; Carroll, B.J. Nuclear gene silencing directs reception of long-distance mRNA silencing in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 14741–14746. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Lepère, G.; Jay, F.; Wang, J.; Bapaume, L.; Wang, Y.; Abraham, A.L.; Penterman, J.; Fischer, R.L.; Voinnet, O.; et al. Dynamics and biological relevance of DNA demethylation in Arabidopsis antibacterial defense. Proc. Natl. Acad. Sci. USA 2013, 5, 2389–2394. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.; Stassen, J.H.M.; Furci, L.; Smith, L.M.; Ton, J. The role of DNA (de)methylation in immune responsiveness of Arabidopsis. Plant J. 2016, 88, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Burgyán, J.; Havelda, Z. Viral suppressors of RNA silencing. Trends Plant Sci. 2011, 16, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Incarbone, M.; Zimmermann, A.; Hammann, P.; Erhardt, M.; Michel, F.; Dunoyer, P. Neutralization of mobile antiviral small RNA through peroxisomal import. Nat. Plants 2017, 3, 17094. [Google Scholar] [CrossRef] [PubMed]
- Murray, G.G.; Kosakovsky, P.S.L.; Obbard, D.J. Suppressors of RNAi from plant viruses are subject to episodic positive selection. Proc. Biol. Sci. 2013, 280, 20130965. [Google Scholar] [CrossRef] [Green Version]
- Daròs, J.A. Viral suppressors: Combatting RNA silencing. Nat. Plants 2017, 3, 17098. [Google Scholar] [CrossRef]
- Ghoshal, B.; Sanfaçon, H. Symptom recovery in virus-infected plants: Revisiting the role of RNA silencing mechanisms. Virology 2015, 479–480, 167–179. [Google Scholar] [CrossRef]
- Zhao, J.H.; Hua, C.L.; Fang, Y.Y.; Guo, H.S. The dual edge of RNA silencing suppressors in the virus-host interactions. Curr. Opin. Virol. 2016, 17, 39–44. [Google Scholar] [CrossRef]
- Yang, Z.; Li, Y. Dissection of RNAi-based antiviral immunity in plants. Curr. Opin. Virol. 2018, 32, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Sidahmed, A.M.; Wilkie, B. Endogenous antiviral mechanisms of RNA interference: A comparative biology perspective. Methods Mol. Biol. 2010, 623, 3–19. [Google Scholar] [CrossRef]
- Voinnet, O. Induction and suppression of RNA silencing: Insights from viral infections. Nat. Rev. Genet. 2005, 6, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Price, D.R.; Gatehouse, J.A. RNAi-mediated crop protection against insects. Trends Biotechnol. 2008, 26, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, H.C.; Miao, X.X. Feasibility, limitation and possible solutions of RNAi-based technology for insect pest control. Insect Sci. 2013, 20, 15–30. [Google Scholar] [CrossRef]
- Lilley, C.J.; Davies, L.J.; Urwin, P.E. RNA interference in plant parasitic nematodes: A summary of the current status. Parasitology 2012, 139, 630–640. [Google Scholar] [CrossRef]
- Nunes, C.C.; Dean, R.A. Host-induced gene silencing: A tool for understanding fungal host interaction and for developing novel disease control strategies. Mol. Plant Pathol. 2012, 13, 519–529. [Google Scholar] [CrossRef]
- Koch, A.; Kogel, K.H. New wind in the sails: Improving the agronomic value of crop plants through RNAi-mediated gene silencing. Plant Biotechnol. J. 2014, 12, 821–831. [Google Scholar] [CrossRef]
- Nowara, D.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; Douchkov, D.; Hensel, G.; Kumlehn, J.; Schweizer, P. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 2010, 22, 3130–3141. [Google Scholar] [CrossRef]
- Koch, A.; Kumar, N.; Weber, L.; Keller, H.; Imani, J.; Kogel, K.H. Host-induced gene silencing of cytochrome P450 lanosterol C14α-demethylase-encoding genes confers strong resistance to Fusarium species. Proc. Natl. Acad. Sci. USA 2013, 110, 19324–19329. [Google Scholar] [CrossRef]
- Qi, T.; Guo, J.; Peng, H.; Liu, P.; Kang, Z.; Guo, J. Host-Induced Gene Silencing: A Powerful Strategy to Control Diseases of Wheat and Barley. Int. J. Mol. Sci. 2019, 20, 206. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Negrete, E.A.; Carrillo-Tripp, J.; Rivera-Bustamante, R.F. RNA silencing against geminivirus: Complementary action of posttranscriptional gene silencing and transcriptional gene silencing in host recovery. J. Virol. 2009, 83, 1332–1340. [Google Scholar] [CrossRef]
- Aregger, M.; Borah, B.K.; Seguin, J.; Rajeswaran, R.; Gubaeva, E.G.; Zvereva, A.S.; Windels, D.; Vazquez, F.; Blevins, T.; Farinelli, L.; et al. Primary and secondary siRNAs in geminivirus-induced gene silencing. PLoS Pathog. 2012, 8, e1002941. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, L. The epidemiology of tomato mosaic VII. The effect of TMV on tomato fruit yield and quality under glass. Ann. Appl. Biol. 1964, 54, 209–224. [Google Scholar] [CrossRef]
- Abel, P.P.; Nelson, R.S.; De, B.; Hoffmann, N.; Rogers, S.G.; Fraley, R.T.; Beachy, R.N. Delay of disease development in transgenic plants that express the tobacco mosaic virus coat protein gene. Science 1986, 232, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Day, A.G.; Bejarano, E.R.; Buck, K.W.; Burrell, M.; Lichtenstein, C.P. Expression of an antisense viral gene in transgenic tobacco confers resistance to the DNA virus tomato golden mosaic virus. Proc. Natl. Acad. Sci. USA 1991, 88, 6721–6725. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.; Azhar, U.; Arshad, M.; Zafar, Y.; Mansoor, S.; Asad, S. Engineering broad-spectrum resistance against RNA viruses in potato. Transgenic Res. 2012, 21, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Hernández, A.M.; Gosalvez, B.; Sempere, R.N.; Burgos, L.; Aranda, M.A.; Truniger, V. Melon RNA interference (RNAi) lines silenced for Cm-eIF4E show broad virus resistance. Mol. Plant Pathol. 2012, 13, 755–763. [Google Scholar] [CrossRef]
- Jelly, N.S.; Schellenbaum, P.; Walter, B.; Maillot, P. Transient expression of artificial microRNAs targeting Grapevine fanleaf virus and evidence for RNA silencing in grapevine somatic embryos. Transgenic Res. 2012, 21, 1319–1327. [Google Scholar] [CrossRef]
- Shekhawat, U.K.; Ganapathi, T.R.; Hadapad, A.B. Transgenic banana plants expressing small interfering RNAs targeted against viral replication initiation gene display high-level resistance to banana bunchy top virus infection. J. Gen. Virol. 2012, 93 Pt 8, 1804–1813. [Google Scholar] [CrossRef]
- Shimizu, T.; Nakazono-Nagaoka, E.; Akita, F.; Wei, T.; Sasaya, T.; Omura, T.; Uehara-Ichiki, T. Hairpin RNA derived from the gene for Pns9, a viroplasm matrix protein of Rice gall dwarf virus, confers strong resistance to virus infection in transgenic rice plants. J. Biotechnol. 2012, 157, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Niu, Y.; Zhang, K.; Liu, Y.; Zhou, X. Virus-derived transgenes expressing hairpin RNA give immunity to Tobacco mosaic virus and Cucumber mosaic virus. Virol. J. 2011, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Kalantidis, K.; Psaradakis, S.; Tabler, M.; Tsagris, M. The occurrence of CMV-specific short Rnas in transgenic tobacco expressing virus-derived double-stranded RNA is indicative of resistance to the virus. Mol. Plant Microbe Interact. 2002, 15, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.Q.; Bai, X.Q.; Wan, X.Q.; Guo, Z.K.; Li, L.J.; Gong, H.Y.; Chu, C.C. Expression of TMV coat protein gene RNAi in transgenic tobacco plants confer immunity to tobacco mosaic virus infection. Yi Chuan 2007, 29, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Ye, J.; Fang, R. Artificial microRNA-mediated virus resistance in plants. J. Virol. 2007, 81, 6690–6699. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.K.; Lohuis, D.; Goldbach, R.; Prins, M. High frequency induction of RNA-mediated resistance against Cucumber mosaic virus using inverted repeat constructs. Mol. Breed. 2004, 14, 215–226. [Google Scholar] [CrossRef]
- Di Nicola-Negri, E.; Brunetti, A.; Tavazza, M.; Ilardi, V. Hairpin RNA-mediated silencing of Plum pox virus P1 and HC-Pro genes for efficient and predictable resistance to the virus. Transgenic Res. 2005, 14, 989–994. [Google Scholar] [CrossRef]
- Kamachi, S.; Mochizuki, A.; Nishiguchi, M.; Tabei, Y. Transgenic Nicotiana benthamiana plants resistant to cucumber green mottle mosaic virus based on RNA silencing. Plant Cell Rep. 2007, 26, 1283–1288. [Google Scholar] [CrossRef]
- Winterhagen, P.; Dubois, C.; Sinn, M.; Wetzel, T.; Reustle, G.M. Gene silencing and virus resistance based on defective interfering constructs in transgenic Nicotiana benthamiana is not linked to accumulation of siRNA. Plant Physiol. Biochem. 2009, 47, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.N.; Palukaitis, P. Resistance to multiple viruses in transgenic tobacco expressing fused, tandem repeat, virus-derived double-stranded RNAs. Virus Genes 2011, 43, 454–464. [Google Scholar] [CrossRef]
- Ai, T.; Zhang, L.; Gao, Z.; Zhu, C.X.; Guo, X. Highly efficient virus resistance mediated by artificial microRNAs that target the suppressor of PVX and PVY in plants. Plant Biol. 2011, 13, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Wagaba, H.; Patil, B.L.; Mukasa, S.; Alicai, T.; Fauquet, C.M. Taylor NJ6. Artificial microRNA-derived resistance to Cassava brown streak disease. J. Virol. Methods 2016, 231, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Patil, B.L.; Ogwok, E.; Wagaba, H.; Mohammed, I.U.; Yadav, J.S.; Bagewadi, B.; Taylor, N.J.; Kreuze, J.F.; Maruthi, M.N.; Alicai, T.; et al. RNAi-mediated resistance to diverse isolates belonging to two virus species involved in Cassava brown streak disease. Mol. Plant Pathol. 2011, 12, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Patil, B.L.; Bagewadi, B.; Yadav, J.S.; Fauquet, C.M. Mapping and identification of cassava mosaic geminivirus DNA-A and DNA-B genome sequences for efficient siRNA expression and RNAi based virus resistance by transient agro-infiltration studies. Virus Res. 2016, 213, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Thu, L.T.M.; Thuy, V.T.X.; Duc, L.H.; Son, L.V.; Ha, C.H.; Mau, C.H. RNAi-mediated resistance to SMV and BYMV in transgenic tobacco. Crop Breed. Appl. Biot. 2016, 16, 213–218. [Google Scholar] [CrossRef]
- Reyes, C.A.; De Francesco, A.; Peña, E.J.; Costa, N.; Plata, M.I.; Sendin, L.; Castagnaro, A.P.; García, M.L. Resistance to Citrus psorosis virus in transgenic sweet orange plants is triggered by coat protein-RNA silencing. J. Biotechnol. 2011, 151, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Batuman, O.; Mawassi, M.; Bar-Joseph, M. Transgenes consisting of a dsRNA of an RNAi suppressor plus the 3′ UTR provide resistance to Citrus tristeza virus sequences in Nicotiana benthamiana but not in citrus. Virus Genes 2006, 33, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Bendahmane, M.; Gronenborn, B. Engineering resistance against tomato yellow leaf curl virus (TYLCV) using antisense RNA. Plant Mol. Biol. 1997, 33, 351–357. [Google Scholar] [CrossRef]
- Mubin, M.; Mansoor, S.; Hussain, M.; Zafar, Y. Silencing of the AV2 gene by antisense RNA protects transgenic plants against a bipartite begomovirus. Virol. J. 2007, 19, 10. [Google Scholar] [CrossRef]
- Asad, S.; Haris, W.A.; Bashir, A.; Zafar, Y.; Malik, K.A.; Malik, N.N.; Lichtenstein, C.P. Transgenic tobacco expressing geminiviral RNAs are resistant to the serious viral pathogen causing cotton leaf curl disease. Arch. Virol. 2003, 148, 2341–2352. [Google Scholar] [CrossRef]
- Nahid, N.; Amin, I.; Briddon, R.W.; Mansoor, S. RNA interference-based resistance against a legume mastrevirus. Virol. J. 2011, 8, 499. [Google Scholar] [CrossRef] [PubMed]
- Medina-Hernández, D.; Rivera-Bustamante, R.F.; Tenllado, F.; Holguín-Peña, R.J. Effects and effectiveness of two RNAi constructs for resistance to Pepper golden mosaic virus in Nicotiana benthamiana plants. Viruses 2013, 5, 2931–2945. [Google Scholar] [CrossRef] [PubMed]
- Schwind, N.; Zwiebel, M.; Itaya, A.; Ding, B.; Wang, M.B.; Krczal, G.; Wassenegger, M. RNAi-mediated resistance to Potato spindle tuber viroid in transgenic tomato expressing a viroid hairpin RNA construct. Mol. Plant Pathol. 2009, 10, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, H.; Zhang, J.; Zhang, C.; Gong, P.; Ziaf, K.; Xiao, F.; Ye, Z. Expression of artificial microRNAs in tomato confers efficient and stable virus resistance in a cell-autonomous manner. Transgenic Res. 2011, 20, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Ntui, V.O.; Kong, K.; Azadi, P.; Khan, R.S.; Chin, D.P.; Igawa, T.; Mii, M.; Nakamura, I. RNAi-Mediated Resistance to Cucumber Mosaic Virus (CMV) in Genetically Engineered Tomato. AJPS 2014, 5, 554–572. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, A.; Ramos, P.L.; Fiallo, E.; Callard, D.; Sánchez, Y.; Peral, R.; Rodríguez, R.; Pujol, M. Intron-hairpin RNA derived from replication associated protein C1 gene confers immunity to tomato yellow leaf curl virus infection in transgenic tomato plants. Transgenic Res. 2006, 15, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Zrachya, A.; Kumar, P.P.; Ramakrishnan, U.; Levy, Y.; Loyter, A.; Arazi, T.; Lapidot, M.; Gafni, Y. Production of siRNA targeted against TYLCV coat protein transcripts leads to silencing of its expression and resistance to the virus. Transgenic Res. 2007, 16, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Nagayama, A.; Ogawa, T.; Ishida, I.; Okada, Y. Transgenic potato expressing a double-stranded RNA-specific ribonuclease is resistant to potato spindle tuber viroid. Nat. Biotechnol. 1997, 15, 1290–1294. [Google Scholar] [CrossRef] [PubMed]
- Kreuze, J.F.; Klein, I.S.; Lazaro, M.U.; Chuquiyuri, W.J.; Morgan, G.L.; Mejía, P.G.; Ghislain, M.; Valkonen, J.P. RNA silencing-mediated resistance to a crinivirus (Closteroviridae) in cultivated sweet potato (Ipomoea batatas L.) and development of sweet potato virus disease following co-infection with a potyvirus. Mol. Plant Pathol. 2008, 9, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Tahir, M.N.; Asad, S.; Bilal, R.; Van Eck, J.; Jander, G.; Mansoor, S. RNAi-Mediated Simultaneous Resistance Against Three RNA Viruses in Potato. Mol. Biotechnol. 2017, 59, 73–83. [Google Scholar] [CrossRef]
- Missiou, A.; Kalantidis, K.; Boutla, A.; Tzortzakaki, S.; Tabler, M.; Tsagris, M. Generation of transgenic potato plants highly resistant to potato virus Y (PVY) through RNA silencing. Mol. Breed. 2004, 14, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Fang, G.; Grumet, R. Genetic engineering of potyvirus resistance using constructs derived from the zucchini yellow mosaic virus coat protein gene. Mol. Plant Microbe Interact. 1993, 6, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Krubphachaya, P.; Jurícek, M.; Kertbundit, S. Induction of RNA-mediated resistance to papaya ringspot virus type W. J. Biochem. Mol. Biol. 2007, 31, 404–411. [Google Scholar] [CrossRef]
- Leibman, D.; Wolf, D.; Saharan, V.; Zelcer, A.; Arazi, T.; Yoel, S.; Gaba, V.; Gal-On, A. A high level of transgenic viral small RNA is associated with broad potyvirus resistance in cucurbits. Mol. Plant Microbe Interact. 2011, 24, 1220–1238. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Ku, H.M.; Chiang, Y.H.; Ho, H.Y.; Yu, T.A.; Jan, F.J. Development of transgenic watermelon resistant to Cucumber mosaic virus and Watermelon mosaic virus by using a single chimeric transgene construct. Transgenic Res. 2012, 21, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Tougou, M.; Furutani, N.; Yamagishi, N.; Shizukawa, Y.; Takahata, Y.; Hidaka, S. Development of resistant transgenic soybeans with inverted repeat-coat protein genes of soybean dwarf virus. Plant Cell Rep. 2006, 25, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sato, S.; Ye, X.; Dorrance, A.E.; Morris, T.J.; Clemente, T.E.; Qu, F. Robust RNAi-based resistance to mixed infection of three viruses in soybean plants expressing separate short hairpins from a single transgene. Phytopathology 2011, 101, 1264–1269. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, M.J.; Pak, J.H.; Jung, H.W.; Choi, H.K.; Lee, Y.H.; Baek, I.Y.; Ko, J.M.; Jeong, S.C.; Pack, I.S.; et al. Characterization of SMV resistance of soybean produced by genetic transformation of SMV-CP gene in RNAi. Plant Biotechnol. Rep. 2013, 7, 425–433. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, M.J.; Pak, J.H.; Im, H.H.; Lee, D.H.; Kim, K.H.; Lee, J.H.; Kim, D.H.; Choi, H.K.; Jung, H.W.; et al. RNAi-mediated Soybean mosaic virus (SMV) resistance of a Korean Soybean cultivar. Plant Biotechnol. Rep. 2016, 10, 257. [Google Scholar] [CrossRef]
- Gao, L.; Ding, X.; Li, K.; Liao, W.; Zhong, Y.; Ren, R.; Liu, Z.; Adhimoolam, K.; Zhi, H. Characterization of Soybean mosaic virus resistance derived from inverted repeat-SMV-HC-Pro genes in multiple soybean cultivars. Theor. Appl. Genet. 2015, 128, 1489–1505. [Google Scholar] [CrossRef]
- Yang, X.; Niu, L.; Zhang, W.; Yang, J.; Xing, G.; He, H.; Guo, D.; Du, Q.; Qian, X.; Yao, Y.; et al. RNAi-mediated SMV P3 cistron silencing confers significantly enhanced resistance to multiple Potyvirus strains and isolates in transgenic soybean. Plant Cell Rep. 2018, 37, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Bonfim, K.; Faria, J.C.; Nogueira, E.O.; Mendes, E.A.; Aragão, F.J. RNAi-mediated resistance to Bean golden mosaic virus in genetically engineered common bean (Phaseolus vulgaris). Mol. Plant Microbe Interact. 2007, 20, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Cruz, A.R.R.; Aragão, F.J.L. RNAi-based enhanced resistance to Cowpea severe mosaic virus and Cowpea aphid-borne mosaic virus in transgenic cowpea. Plant Pathol. 2014, 63, 831–837. [Google Scholar] [CrossRef]
- Ludlow, E.J.; Mouradov, A.; Spangenberg, G.C. Post-transcriptional gene silencing as an efficient tool for engineering resistance to white clover mosaic virus in white clover (Trifolium repens). J. Plant Physiol. 2009, 166, 1557–1567. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, H.; Rajasubramaniam, S.; Rajam, M.V.; Dasgupta, I. RNA-interference in rice against Rice tungro bacilliform virus results in its decreased accumulation in inoculated rice plants. Transgenic Res. 2008, 17, 897–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, T.; Yoshii, M.; Wei, T.; Hirochika, H.; Omura, T. Silencing by RNAi of the gene for Pns12, a viroplasm matrix protein of Rice dwarf virus, results in strong resistance of transgenic rice plants to the virus. Plant Biotechnol. 2009, 7, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Song, Y.; Wu, B.; Jiang, M.; Li, K.; Zhu, C.; Wen, F. Production of transgenic rice new germplasm with strong resistance against two isolations of Rice stripe virus by RNA interference. Transgenic Res. 2011, 20, 1367–1377. [Google Scholar] [CrossRef]
- Shimizu, T.; Nakazono-Nagaoka, E.; Uehara-Ichiki, T.; Sasaya, T.; Omura, T. Targeting specific genes for RNA interference is crucial to the development of strong resistance to Rice stripe virus. Plant Biotechnol. J. 2011, 9, 503–512. [Google Scholar] [CrossRef]
- Shimizu, T.; Nakazono-Nagaoka, E.; Akita, F.; Uehara-Ichiki, T.; Sasaya, T.; Omura, T. Immunity to Rice black streaked dwarf virus, a plant reovirus, can be achieved in rice plants by RNA silencing against the gene for the viroplasm component protein. Virus Res. 2011, 160, 400–403. [Google Scholar] [CrossRef]
- Shimizu, T.; Ogamino, T.; Hiraguri, A.; Nakazono-Nagaoka, E.; Uehara-Ichiki, T.; Nakajima, M.; Akutsu, K.; Omura, T.; Sasaya, T. Strong resistance against Rice grassy stunt virus is induced in transgenic rice plants expressing double-stranded RNA of the viral genes for nucleocapsid or movement proteins as targets for RNA interference. Phytopathology 2013, 103, 513–519. [Google Scholar] [CrossRef]
- Fahim, M.; Ayala-Navarrete, L.; Millar, A.A.; Larkin, P.J. Hairpin RNA derived from viral NIa gene confers immunity to wheat streak mosaic virus infection in transgenic wheat plants. Plant Biotechnol. J. 2010, 8, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Fahim, M.; Millar, A.A.; Wood, C.C.; Larkin, P.J. Resistance to Wheat streak mosaic virus generated by expression of an artificial polycistronic microRNA in wheat. Plant Biotechnol. J. 2012, 10, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Yang, L.; Zhou, S.F.; Wang, H.G.; Li, W.C.; Fu, F.L. Improvement of resistance to maize dwarf mosaic virus mediated by transgenic RNA interference. J. Biotechnol. 2011, 153, 181–187. [Google Scholar] [CrossRef]
- Wang, M.-B.; Abbott, D.C.; Waterhouse, P.M. A single copy of a virus-derived transgene encoding hairpin RNA gives immunity to Barley yellow dwarf virus. Mol. Plant Pathol. 2000, 1, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Vanderschuren, H.; Fütterer, J.; Gruissem, W. Resistance to cassava mosaic disease in transgenic cassava expressing antisense RNAs targeting virus replication genes. Plant Biotechnol. J. 2005, 3, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Vanderschuren, H.; Akbergenov, R.; Pooggin, M.M.; Hohn, T.; Gruissem, W.; Zhang, P. Transgenic cassava resistance to African cassava mosaic virus is enhanced by viral DNA-A bidirectional promoter-derived siRNAs. Plant Mol. Biol. 2007, 64, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Vanderschuren, H.; Alder, A.; Zhang, P.; Gruissem, W. Dose-dependent RNAi-mediated geminivirus resistance in the tropical root crop cassava. Plant Mol. Biol. 2009, 70, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, R.D.; Beyene, G.; Kalyaeva, M.; Fauquet, C.M.; Taylor, N. Improvements in Agrobacterium-mediated transformation of cassava (Manihot esculenta Crantz) for large-scale production of transgenic plants. Plant Cell Tissue Organ Cult. 2015, 121, 591. [Google Scholar] [CrossRef]
- Wagaba, H.; Beyene, G.; Aleu, J.; Odipio, J.; Okao-Okuja, G.; Chauhan, R.D.; Munga, T.; Obiero, H.; Halsey, M.E.; Ilyas, M.; et al. Field Level RNAi-Mediated Resistance to Cassava Brown Streak Disease across Multiple Cropping Cycles and Diverse East African Agro-Ecological Locations. Front. Plant Sci. 2017, 7, 2060. [Google Scholar] [CrossRef] [Green Version]
- Vanderschuren, H.; Moreno, I.; Anjanappa, R.B.; Zainuddin, I.M.; Gruissem, W. Exploiting the combination of natural and genetically engineered resistance to cassava mosaic and cassava brown streak viruses impacting cassava production in Africa. PLoS ONE 2012, 7, e45277. [Google Scholar] [CrossRef]
- Yadav, J.S.; Ogwok, E.; Wagaba, H.; Patil, B.L.; Bagewadi, B.; Alicai, T.; Gaitan-Solis, E.; Taylor, N.J. Fauquet CM. RNAi-mediated resistance to Cassava brown streak Uganda virus in transgenic cassava. Mol. Plant Pathol. 2011, 12, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Ogwok, E.; Odipio, J.; Halsey, M.; Gaitán-Solís, E.; Bua, A.; Taylor, N.J.; Fauquet, C.M.; Alicai, T. Transgenic RNA interference (RNAi)-derived field resistance to cassava brown streak disease. Mol. Plant Pathol. 2012, 13, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Odipio, J.; Ogwok, E.; Taylor, N.J.; Halsey, M.; Bua, A.; Fauquet, C.M.; Alicai, T. RNAi-derived field resistance to Cassava brown streak disease persists across the vegetative cropping cycle. GM Crops Food 2014, 5, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Beyene, G.; Chauhan, R.D.; Ilyas, M.; Wagaba, H.; Fauquet, C.M.; Miano, D.; Alicai, T.; Taylor, N.J. A Virus-Derived Stacked RNAi Construct Confers Robust Resistance to Cassava Brown Streak Disease. Front. Plant Sci. 2017, 18, 2052. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.L.; Spetz, C.; Haugslien, S.; Xing, S.; Dees, M.W.; Moe, R.; Blystad, D.R. Agrobacterium tumefaciens-mediated transformation of poinsettia, Euphorbia pulcherrima, with virus-derived hairpin RNA constructs confers resistance to Poinsettia mosaic virus. Plant Cell Rep. 2008, 27, 1027–1038. [Google Scholar] [CrossRef] [Green Version]
- López, C.; Cervera, M.; Fagoaga, C.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Accumulation of transgene-derived siRNAs is not sufficient for RNAi-mediated protection against Citrus tristeza virus in transgenic Mexican lime. Mol. Plant Pathol. 2010, 11, 33–41. [Google Scholar] [CrossRef]
- Soler, N.; Plomer, M.; Fagoaga, C.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Transformation of Mexican lime with an intron-hairpin construct expressing untranslatable versions of the genes coding for the three silencing suppressors of Citrus tristeza virus confers complete resistance to the virus. Plant Biotechnol. J. 2012, 10, 597–608. [Google Scholar] [CrossRef]
- Reyes, C.A.; Pena, E.J.; Zanek, M.C.; Sanchez, D.V.; Grau, O.; Garcia, M.L. Differential resistance to Citrus psorosis virus in transgenic Nicotiana benthamiana plants expressing hairpin RNA derived from the coat protein and 54K protein genes. Plant Cell Rep. 2009, 28, 1817–1825. [Google Scholar] [CrossRef]
- Niu, Q.W.; Lin, S.S.; Reyes, J.L.; Chen, K.C.; Wu, H.W.; Yeh, S.D.; Chua, N.H. Expression of artificial microRNAs in transgenic Arabidopsis thaliana confers virus resistance. Nat. Biotechnol. 2006, 24, 1420–1428. [Google Scholar] [CrossRef]
- Duan, C.G.; Wang, C.H.; Fang, R.X.; Guo, H.S. Artificial MicroRNAs highly accessible to targets confer efficient virus resistance in plants. J. Virol. 2008, 82, 11084–11095. [Google Scholar] [CrossRef]
- Mitiouchkina, T.Y.; Firsov, A.P.; Titova, S.M.; Pushin, A.S.; Shulga, O.A.; Dolgov, S.V. Different Approaches to Produce Transgenic Virus B Resistant Chrysanthemum. Agronomy 2018, 8, 28. [Google Scholar] [CrossRef]
- Pavli, O.I.; Panopoulos, N.J.; Goldbach, R.; Skaracis, G.N. BNYVV-derived dsRNA confers resistance to rhizomania disease of sugar beet as evidenced by a novel transgenic hairy root approach. Transgenic Res. 2010, 19, 915–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Gao, S.; Lin, Q.; Wang, H.; Que, Y.; Xu, L. Transgenic Sugarcane Resistant to Sorghum mosaic virus Based on Coat Protein Gene Silencing by RNA Interference. BioMed Res. Int. 2015, 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Szittya, G.; Silhavy, D.; Molnár, A.; Havelda, Z.; Lovas, A.; Lakatos, L.; Bánfalvi, Z.; Burgyán, J. Low temperature inhibits RNA silencing-mediated defence by the control of siRNA generation. EMBO J. 2003, 22, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalakouras, A.; Tzanopoulou, M.; Tsagris, M.; Wassenegger, M.; Kalantidis, K. Hairpin transcription does not necessarily lead to efficient triggering of the RNAi pathway. Transgenic Res. 2011, 20, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Weising, K.; Schell, J.; Kahl, G. Foreign genes in plants: Transfer, structure, expression, and applications. Annu. Rev. Genet. 1988, 22, 421–477. [Google Scholar] [CrossRef]
- Han, Y.; Griffiths, A.; Li, H.; Grierson, D. The effect of endogenous mRNA levels on co-suppression in tomato. FEBS Lett. 2004, 563, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, W.; Shimura, H.; Shirakawa, S.; Isoda, R.; Inukai, T.; Matsumura, T.; Masuta, C. Transcriptional silencing of 35S driven-transgene is differentially determined depending on promoter methylation heterogeneity at specific cytosines in both plus-and minus-sense strands. BMC Plant Biol. 2019, 19, 24. [Google Scholar] [CrossRef]
- Rajeevkumar, S.; Anunanthini, P.; Sathishkumar, R. Epigenetic silencing in transgenic plants. Front. Plant Sci. 2015, 6, 693. [Google Scholar] [CrossRef] [Green Version]
- Hosseinahli, N.; Aghapour, M.; Duijf, P.H.G.; Baradaran, B. Treating cancer with microRNA replacement therapy: A literature review. J. Cell Physiol. 2018, 233, 5574–5588. [Google Scholar] [CrossRef]
- Carbonell, A. Design and High-Throughput Generation of Artificial Small RNA Constructs for Plants. Methods Mol. Biol. 2019, 1932, 247–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, B.N.; Yoon, J.Y.; Palukaitis, P. Engineered resistance in potato against potato leafroll virus, potato virus A and potato virus Y. Virus Genes 2013, 47, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.; Kaniewski, W.; Haley, L.; Rozman, R.; Newell, C.; Sanders, P.; Tumer, N.E. Engineering resistance to mixed virus infection in a commercial potato cultivar: Resistance to potato virus X and potato virus Y in transgenic Russet Burbank. Biotechnology 1990, 8, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.J.; Brown, S.; Yuanhai, H.; Ishizuka, M.; Lowe, A.; Solis, A.A.; Grierson, D. A transgene with repeated DNA causes high frequency, post-transcriptional suppression of ACC-oxidase gene expression in tomato. Plant J. 1998, 15, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, P.M.; Graham, M.W.; Wang, M.B. Virus resistance and gene silencing in plants can be induced by simultaneous expression of sense and antisense RNA. Proc. Natl. Acad. Sci. USA 1998, 95, 13959–13964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wesley, S.V.; Helliwell, C.A.; Smith, N.A.; Wang, M.B.; Rouse, D.T.; Liu, Q.; Gooding, P.S.; Singh, S.P.; Abbott, D.; Stoutjesdijk, P.A.; et al. Construct design for efficient, effective and high-throughput gene silencing in plants. Plant J. 2001, 27, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Provvidenti, R. Diseases caused by viruses. In Compendium of Cucurbit Diseases; Zitter, T.A., Hopkins, D.L., Thomas, C.E., Eds.; APS Press: Minnesota, MN, USA, 1996; pp. 37–45. [Google Scholar]
- Gaba, V.; Zelcer, A.; Gal-On, A. cucurbit biotechnology—the importance of virus resistance. In Vitro Cell. Dev. Biol.-Plant 2004, 40, 346–358. [Google Scholar] [CrossRef]
- Truniger, V.; Aranda, M.A. Recessive resistance to plant viruses. Adv. Virus Res. 2009, 75, 119–159. [Google Scholar] [CrossRef]
- Jan, F.J.; Fagoaga, C.; Pang, S.Z.; Gonsalves, D. A minimum length of N gene sequence in transgenic plants is required for RNA-mediated tospovirus resistance. J. Gen. Virol. 2000, 81 Pt 1, 235–242. [Google Scholar] [CrossRef]
- Pang, S.-Z.; Jan, F.-J.; Tricoli, D.M.; Russell, P.F.; Carney, K.J.; Hu, J.S.; Fuchs, M.; Quemada, H.D.; Gonsalves, D. Resistance tosquash mosaic comovirus in transgenic squash plants expressing its coatprotein genes. Mol. Breed. 2000, 6, 87–93. [Google Scholar] [CrossRef]
- Wu, H.W.; Yu, T.A.; Raja, J.A.; Wang, H.C.; Yeh, S.D. Generation of transgenic oriental melon resistant to Zucchini yellow mosaic virus by an improved cotyledon-cutting method. Plant Cell Rep. 2009, 28, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.W.; Yu, T.A.; Raja, J.A.J.; Christopher, S.J.; Wang, S.L.; Yeh, S.D. Double-Virus Resistance of Transgenic Oriental Melon Conferred by Untranslatable Chimeric Construct Carrying Partial Coat Protein Genes of Two Viruses. Plant Dis. 2010, 94, 1341–1347. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.A.; Chiang, C.H.; Wu, H.W.; Li, C.M.; Yang, C.F.; Chen, J.H.; Chen, Y.W.; Yeh, S.D. Generation of transgenic watermelon resistant to Zucchini yellow mosaic virus and Papaya ringspot virus type W. Plant Cell Rep. 2011, 30, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.Z.; Jan, F.J.; Gonsalves, D. Nontarget DNA sequences reduce the transgene length necessary for RNA-mediated tospovirus resistance in transgenic plants. Proc. Natl. Acad. Sci. USA 1997, 94, 8261–8266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, F.J.; Fagoaga, C.; Pang, S.Z.; Gonsalves, D. A single chimeric transgene derived from two distinct viruses confers multi-virus resistance in transgenic plants through homology-dependent gene silencing. J. Gen. Virol. 2000, 81 Pt 8, 2103–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradshaw, J.D.; Rice, M.E.; Hill, J.H. Evaluation of management strategies for bean leaf beetles (Coleoptera: Chrysomelidae) and Bean pod mottle virus (Comoviridae) in soybean. J. Econ. Entomol. 2008, 101, 1211–1227. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lin, J.; Zheng, G.; Zhang, M.; Zhi, H. Recombinant soybean mosaic virus is prevalent in Chinese soybean fields. Arch. Virol. 2014, 159, 1793–1796. [Google Scholar] [CrossRef]
- Jenner, C.E.; Wang, X.; Tomimura, K.; Ohshima, K.; Ponz, F.; Walsh, J.A. The dual role of the potyvirus P3 protein of Turnip mosaic virus as a symptom and avirulence determinant in brassicas. Mol. Plant Microbe Interact. 2003, 16, 777–784. [Google Scholar] [CrossRef]
- Suehiro, N.; Natsuaki, T.; Watanabe, T.; Okuda, S. An important determinant of the ability of Turnip mosaic virus to infect Brassica spp. and/or Raphanus sativus is in its P3 protein. J. Gen. Virol. 2004, 85 Pt 7, 2087–2098. [Google Scholar] [CrossRef]
- Wang, Y.; Khatabi, B.; Hajimorad, M.R. Amino acid substitution in P3 of Soybean mosaic virus to convert avirulence to virulence on Rsv4-genotype soybean is influenced by the genetic composition of P3. Mol. Plant Pathol. 2015, 16, 301–307. [Google Scholar] [CrossRef]
- Wang, X.; Eggenberger, A.L.; Nutter, F.W.; John, H. Pathogen derived transgenic resistance to soybean mosaic virus in soybean. Mol. Breed. 2001, 8, 119. [Google Scholar] [CrossRef]
- Furutani, N.; Hidaka, H.; Kosaka, Y.; Shizukawa, Y.; Kanematsu, S. Coat protein gene-mediated resistance to soybean mosaic virus in transgenic soybean. Breed. Sci. 2006, 56, 119–124. [Google Scholar] [CrossRef]
- Furutani, N.; Yamagishi, N.; Hidaka, S.; Shizukawa, Y.; Kanematsu, S.; Kosaka, Y. Soybean mosaic virus resistance in transgenic soybean caused by post-transcriptional gene silencing. Breed. Sci. 2007, 57, 123–128. [Google Scholar] [CrossRef]
- Lim, H.S.; Ko, T.S.; Hobbs, H.A.; Lambert, K.N.; Yu, J.M.; Mccoppin, N.K.; Korban, S.S.; Hartman, G.L.; Domier, L.L. Soybean mosaic virus helper component-protease alters leaf morphology and reduces seed production in transgenic soybean plants. Phytopathology 2007, 97, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Tougou, M.; Yamagishi, N.; Furutani, N.; Shizukawa, Y.; Takahata, Y.; Hidaka, S. Soybean dwarf virus-resistant transgenic soybeans with the sense coat protein gene. Plant Cell Rep. 2007, 26, 1967–1975. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO. Evaluation of Allergenicity of Genetically Modified Foods; Report of a joint FAO/WHO expert consultation on allergenicity of foods derived from biotechnology; Food and Agriculture Organization of the United Nations: Rome, Italy, 2001. [Google Scholar]
- Zhang, X.; Zhao, P.; Wu, K.; Zhang, Y.; Peng, M.; Liu, Z. Compositional equivalency of RNAi-mediated virus-resistant transgenic soybean and its nontransgenic counterpart. J. Agric. Food Chem. 2014, 14, 4475–4479. [Google Scholar] [CrossRef]
- OECD (Organisation for Economic Cooperation and Development). Concepts and Principles; International Energy Agency Publications: Paris, France, 1993. [Google Scholar]
- Omura, T.; Yan, J. Role of outer capsid proteins in transmission of Phytoreovirus by insect vectors. Adv. Virus Res. 1999, 54, 15–43. [Google Scholar]
- Wei, T.; Kikuchi, A.; Moriyasu, Y.; Suzuki, N.; Shimizu, T.; Hagiwara, K.; Chen, H.; Takahashi, M.; Ichiki-Uehara, T.; Omura, T. The spread of Rice dwarf virus among cells of its insect vector exploits virus-induced tubular structures. J. Virol. 2006, 80, 8593–8602. [Google Scholar] [CrossRef]
- Wei, T.; Kikuchi, A.; Suzuki, N.; Shimizu, T.; Hagiwara, K.; Chen, H.; Omura, T. Pns4 of rice dwarf virus is a phosphoprotein, is localized around the viroplasm matrix, and forms minitubules. Arch. Virol. 2006, 151, 1701–1712. [Google Scholar] [CrossRef]
- Ellis, M.H.; Rebetzke, G.J.; Chu, P. First report of Wheat streak mosaic virus in Australia. Plant Pathol. 2003, 52, 808. [Google Scholar] [CrossRef]
- Ellis, M.H.; Rebetzke, G.J.; Kelman, W.M.; Moore, C.S.; Hyles, J.E. Detection of Wheat streak mosaic virus in four pasture grass species in Australia. Plant Pathol. 2004, 53, 239. [Google Scholar] [CrossRef]
- Miller, W.A.; Rasochová, L. Barley yellow dwarf viruses. Annu. Rev. Phytopathol. 1997, 35, 167–190. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O.; Pinto, Y.M.; Baulcombe, D.C. Suppression of gene silencing: A general strategy used by diverse DNA and RNA viruses of plants. Proc. Natl. Acad. Sci. USA 1999, 23, 14147–14152. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.J.; Halsey, M.; Gaitán-Solís, E.; Anderson, P.; Gichuki, S.; Miano, D.; Bua, A.; Alicai, T.; Fauquet, C.M. The VIRCA Project: Virus resistant cassava for Africa. GM Crops Food 2012, 3, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Chellappan, P.; Masona, M.V.; Vanitharani, R.; Taylor, N.J.; Fauquet, C.M. Broad spectrum resistance to ssDNA viruses associated with transgene-induced gene silencing in cassava. Plant Mol. Biol. 2004, 56, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.F.; Rocha-Pena, M.A. Citrus tristeza virus, Plant Disease of International Importance. In Disease of Fruit Crops; Kumar, J., Chaube, H.S., Singh, U.S., Mukhopdhyuy, A.N., Eds.; Prentice Hall: Englewood Cliffs, NJ, USA, 1992; Volume III, pp. 226–249. [Google Scholar]
- Fagoaga, C.; López, C.; de Mendoza, A.H.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Post-transcriptional gene silencing of the p23 silencing suppressor of Citrus tristeza virus confers resistance to the virus in transgenic Mexican lime. Plant Mol. Biol. 2006, 60, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Ghorbel, R.; López, C.; Fagoaga, C.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Transgenic citrus plants expressing the citrus tristeza virus p23 protein exhibit viral-like symptoms. Mol. Plant Pathol. 2001, 2, 27–36. [Google Scholar] [CrossRef]
- Fagoaga, C.; López, C.; Moreno, P.; Navarro, L.; Flores, R.; Peña, L. Viral-like symptoms induced by the ectopic expression of the p23 gene of Citrus tristeza virus are citrus specific and do not correlate with the pathogenicity of the virus strain. Mol. Plant Microbe Interact. 2005, 18, 435–445. [Google Scholar] [CrossRef]
- Noris, E.; Lucioli, A.; Tavazza, R.; Caciagli, P.; Accotto, G.P.; Tavazza, M. Tomato yellow leaf curl Sardinia virus can overcome transgene-mediated RNA silencing of two essential viral genes. J. Gen. Virol. 2004, 85 Pt 6, 1745–1749. [Google Scholar] [CrossRef]
- Ribeiro, S.G.; Lohuis, H.; Goldbach, R.; Prins, M. Tomato chlorotic mottle virus is a target of RNA silencing but the presence of specific short interfering RNAs does not guarantee resistance in transgenic plants. J. Virol. 2007, 81, 1563–1573. [Google Scholar] [CrossRef]
- Tenllado, F.; Díaz-Ruíz, J.R. Double-stranded RNA-mediated interference with plant virus infection. J. Virol. 2001, 75, 12288–12297. [Google Scholar] [CrossRef] [PubMed]
- Holeva, M.; Sclavounos, A.P.; Kyriakopoulou, P.E.; Voloudakis, A.E. In Vitro produced dsRNA induces resistance against a sever Hellenic CMV isolate in tobacco and tomato. In Proceedings of the 8th International Congress of Plant Molecular Biology, Adelaide, Australia, 20–25 August 2006. [Google Scholar]
- Holeva, M.C.; Sclavounos, A.P.; Milla, S.P.; Kyriakopoulou, P.E.; Voloudakis, A.E. External application of dsRNA of the capsid protein (CP) or 2b gene of CMV reduces the severity of CMV-infection in tobacco. In Proceedings of the VIII International Congress on Molecular Plant–Microbe Interactions, Sorrento, Italy, 21–27 July 2007. [Google Scholar]
- Yin, G.; Sun, Z.; Liu, N.; Zhang, L.; Song, Y.; Zhu, C.; Wen, F. Production of double-stranded RNA for interference with TMV infection utilizing a bacterial prokaryotic expression system. Appl. Microbiol. Biotechnol. 2009, 84, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Gan, D.; Zhang, J.; Jiang, H.; Jiang, T.; Zhu, S.; Cheng, B. Bacterially expressed dsRNA protects maize against SCMV infection. Plant Cell Rep. 2010, 29, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Biedenkopf, B.; Furch, A.C.U.; Abdellatef, E.; Weber, L.; Linicus, L.; Johannsmeier, J.; Jelonek, L.; Goesmann, A.; Cardoza, V.; et al. An RNAi-based control of Fusarium graminearum infections through spraying of long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016, 12, e1005901. [Google Scholar] [CrossRef]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Xu, Z.P.; Carroll, B.J. Induction of virus resistance by exogenous application of double-stranded RNA. Curr. Opin. Virol. 2017, 26, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Konakalla, N.C.; Kaldis, A.; Berbati, M.; Masarapu, H.; Voloudakis, A.E. Exogenous application of double-stranded RNA molecules from TMV p126 and CP genes confers resistance against TMV in tobacco. Planta 2016, 244, 961–969. [Google Scholar] [CrossRef]
- Kaldis, A.; Berbati, M.; Melita, O.; Reppa, C.; Holeva, M.; Otten, P.; Voloudakis, A. Exogenously applied dsRNA molecules deriving from the Zucchini yellow mosaic virus (ZYMV) genome move systemically and protect cucurbits against ZYMV. Mol. Plant Pathol. 2018, 19, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Niehl, A.; Soininen, M.; Poranen, M.M.; Heinlein, M. Synthetic biology approach for plant protection using dsRNA. Plant Biotechnol. J. 2018, 16, 1679–1687. [Google Scholar] [CrossRef] [Green Version]
- Ladewig, K.; Niebert, M.; Xu, Z.P.; Gray, P.P.; Lu, G.Q. Efficient siRNA delivery to mammalian cells using layered double hydroxide nanoparticles. Biomaterials 2010, 31, 1821–1829. [Google Scholar] [CrossRef]
- Ladewig, K.; Xu, Z.P.; Lu, G.Q. Layered double hydroxide nanoparticles in gene and drug delivery. Expert Opin. Drug Deliv. 2009, 6, 907–922. [Google Scholar] [CrossRef]
- Voloudakis, A.E.; Holeva, M.C.; Sarin, L.P.; Bamford, D.H.; Vargas, M.; Poranen, M.M.; Tenllado, F. Efficient double-stranded RNA production methods for utilization in plant virus control. Methods Mol. Biol. 2015, 1236, 255–274. [Google Scholar] [PubMed]
Figure 1.
Antiviral RNA silencing starts with the initial processing or cleavage of a precursor viral dsRNA into short 21–24 nucleotide vsiRNA duplexes by RNaseIII-like enzymes called DCLs. Double-stranded siRNAs are incorporated into an RNA-induced silencing complex RISC, and the activated RISC subsequently unwinds the vsiRNA, thereby generating an antisense (or guide) strand that targets complementary mRNA transcripts via base-pairing interactions. Subsequent degradation of the targeted viral mRNA or inhibition of translation can interfere with protein biosynthesis.
Figure 1.
Antiviral RNA silencing starts with the initial processing or cleavage of a precursor viral dsRNA into short 21–24 nucleotide vsiRNA duplexes by RNaseIII-like enzymes called DCLs. Double-stranded siRNAs are incorporated into an RNA-induced silencing complex RISC, and the activated RISC subsequently unwinds the vsiRNA, thereby generating an antisense (or guide) strand that targets complementary mRNA transcripts via base-pairing interactions. Subsequent degradation of the targeted viral mRNA or inhibition of translation can interfere with protein biosynthesis.


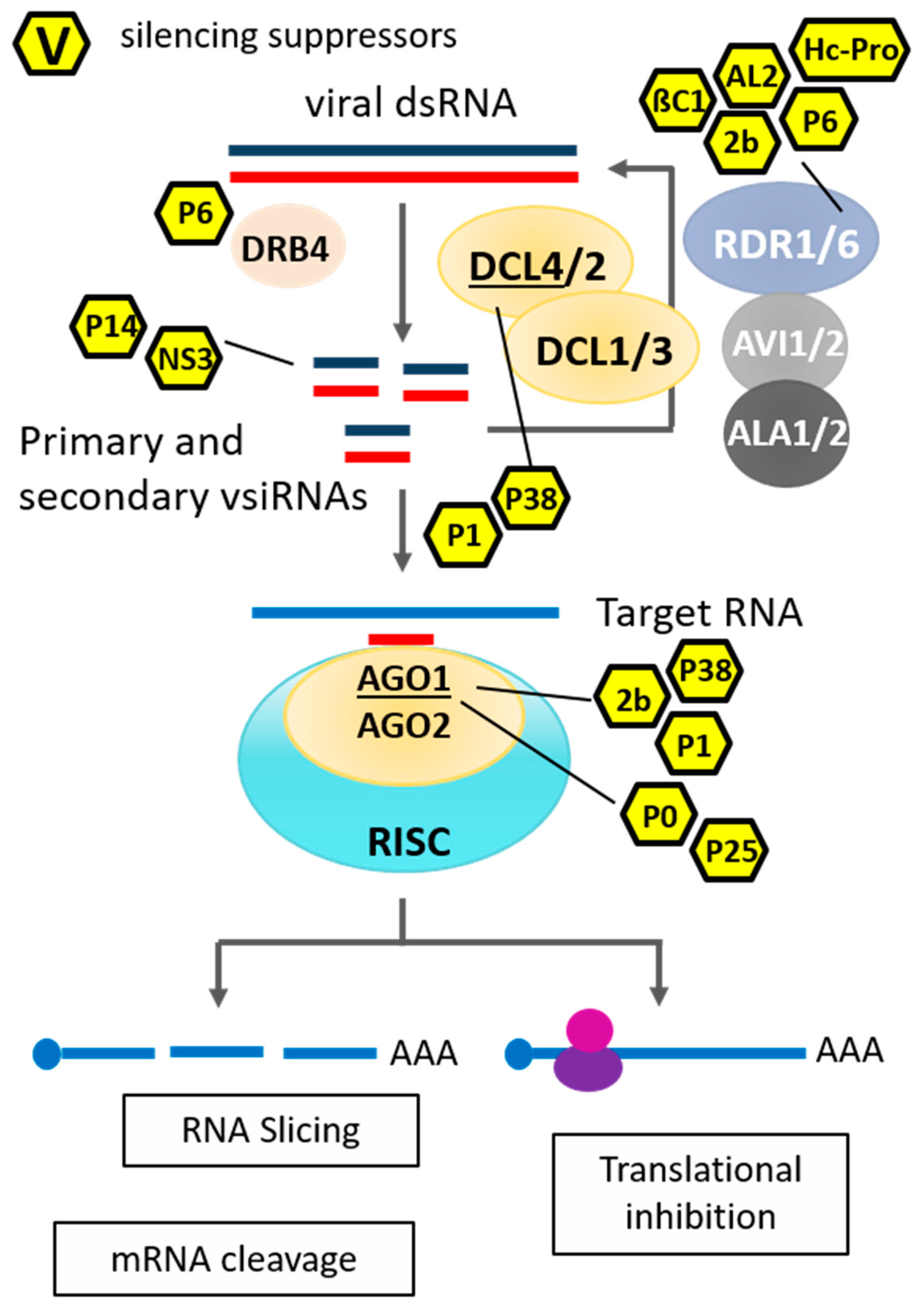{kind=link}
Table 1.
Summary of HIGS applied to control viral pathogens. Host plant family, virus species and genome, target genes, type of RNA silencing trigger and construct length are summarized Hairpin RNA (hpRNA); artificial miRNA (amiRNA).
Table 1.
Summary of HIGS applied to control viral pathogens. Host plant family, virus species and genome, target genes, type of RNA silencing trigger and construct length are summarized Hairpin RNA (hpRNA); artificial miRNA (amiRNA).
| Solanaceae | Virus | Abbr. | Type | Target Gene | RNA Type | Construct Length | Reference |
| Tobacco | |||||||
| tobacco mosaic virus | (+) ssRNA | Movement protein | IR (hpRNA) | not mentioned | [94] | ||
| cucumber mosaic virus | CMV | (+) ssRNA | Replicase | IR (hpRNA) | not mentioned | [94] | |
| cucumber mosaic virus | CMV | (+) ssRNA | Coat protein | IR (hpRNA) | 718 bp | [95] | |
| tobacco mosaic virus | TMV | (+) ssRNA | Coat protein | dsRNA | not mentioned | [96] | |
| cucumber mosaic virus | CMV | (+) ssRNA | VSR 2b | amiRNAs | Precursor miR171a | [97] | |
| cucumber mosaic virus | CMV | (+) ssRNA | Coat protein | hpRNA | [98] | ||
| plum pox virus | PPV | (+) ssRNA | VSR P1 and HC-Pro | IR (hpRNA) | 733 bp (UTR/P1) | [99] | |
| 649 bp (P1/HC-Pro) | |||||||
| 706 bp (HC-Pro) | |||||||
| 678 bp (HC-Pro/P3) | |||||||
| cucumber green mottle mosaic virus | CGMMV | (+) ssRNA | Coat protein | IR (hpRNA) | not mentioned | [100] | |
| tomato bushy stunt virus grapevine leaf virus | TBSV | (+) ssRNA | Defective interfering (DI) Movement protein | Virus-derived (DI) RNA | 720 bp | [101] | |
| GFLV | (+) ssRNA | ||||||
| potato virus Y | PVY | (+) ssRNA | Coat protein | fused, tandem, IR; 200 bp (from each virus) | 600 bp fragment (PVY, PVA, PLRV) | [102] | |
| potato virus A | PVA | (+) ssRNA | cylindrical inclusion (CI) | 1000 bp fragment (PVY, PVA, PLRV, TRV, PMTV) | |||
| potato leafroll virus | PLRV | (+) ssRNA | Coat protein | ||||
| tobacco rattle virus | TRV | (+) ssRNA | Replicase | ||||
| potato mop-top virus | PMTV | (+) ssRNA | Replicase | ||||
| potato virus Y | PVY | (+) ssRNA | VSR HC-Pro | amiRNAs | Precursor miR159a, miR167b and miR171a | [103] | |
| potato virus X | PVX | (+) ssRNA | VSR TGBp1, p25 (p25) | ||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | conserved sequences of P1(CBSV and UCBSV), P3(CBSV and UCBSV), CI(UCBSV), NIb(CBSV and UCBSV), | amiRNA | Precursor miR159a | [104] | |
| cassava brown streak virus | (+) ssRNA | CP(UCBSV) and the 3′UTR | |||||
| cassava brown streak Uganda virus | CBSUV | (+) ssRNA | Coat protein (CBSUV) | hpRNA (pHELLSGATE) | Full-length; N-term. 397 bp; 491 C-term. | [105] | |
| cassava brown streak virus | CBSV | (+) ssRNA | |||||
| African cassava mosaic virus | ACMV | ssDNA | DNA-A and DNA-B genome | dsRNA | 12 constructs of different length | [106] | |
| soybean mosaic virus | SMV | (+) ssRNA | Coat protein | hpRNA (pK7GWIWG2) | not mentioned | [107] | |
| bean yellow mosaic virus | BYMV | (+) ssRNA | Coat protein | hpRNA (pK7GWIWG2) | not mentioned | [107] | |
| citrus psorosis virus | CPsV | (−) ssRNA | Coat protein and 54K gene | hpRNA (pHANNIBAL) | 372 bp (CP); 436 bp (54K) | [108] | |
| citrus tristeza virus | CTV | (+) ssRNA | 3′ p23 (VSR) and 3′ UTR | hpRNA | 900 bp | [109] | |
| tomato golden mosaic virus | TGMV | ssDNA | Replicase (AL1; AL2; AL3 gene) | antisense construct | 1258 bp | [88] | |
| tomato yellow leaf curl virus | TYLCV | ssDNA | Replicase (C1 gene) | antisense RNA | [110] | ||
| tomato leaf curl New Delhi virus | ToLCNDV | ssDNA | virion-sense gene (AV2) | antisense construct | [111] | ||
| cotton leaf curl disease | CLCuD | ssDNA | Replicase (AC1) | sense and antisense RNAs | 446 bp (AC1(5′half)+AC4(ORF)) | [112] | |
| Transcription activator (AC2) | 523 bp (AC1 (3′half)+AC2,AC4) | ||||||
| Rep enhancer (AC3) | 510 bp (AC1(97bp) +AC2+AC3) | ||||||
| chickpea chlorotic dwarf Pakistan virus | CpCDPKV | ssDNA | Rep gene, large intergenic region (LIR) and part of the MP gene | hpRNA | 730 bp | [113] | |
| 5′ Rep, LIR and 5′ MP | |||||||
| pepper golden mosaic virus | PepGMV | ssDNA | Replicase (AC1), intergenic region (IR) and coat protein (AV1) | hpRNA | [114] | ||
| Tomato | potato spindle tuber viroid | PSTVd | (+) ssRNA | Viral sequence | hpRNA | [115] | |
| cucumber mosaic virus | CMV | (+) ssRNA | 2a and 2b genes 3′ UTR | amiRNA | Precursor miR159a | [116] | |
| cucumber mosaic virus | CMV | (+) ssRNA | Replicase | hpRNA | 1138 bp | [117] | |
| tomato yellow leaf curl virus | TYLCV | ssDNA | Replicase (C1) | hpRNA | [118] | ||
| tomato yellow leaf curl virus | TYLCV | ssDNA | Coat protein | hpRNA | [119] | ||
| Potato | potato spindle tuber viroid | PSTVd | (+) ssRNA | Ribunuclease pac1 | dsRNA | [120] | |
| potato virus X | PVX | (+) ssRNA | ORF2 of PVX | hpRNA | 300 bo (ORF2-PVX) | [89] | |
| potato virus Y | PVY | (+) ssRNA | Protease gene PVY | 365 bp (HC-Pro-PVY) | |||
| potato leaf roll virus | PLRV | (+) ssRNA | CP gene PLRV | 300 bp (CP-PLRV) | |||
| sweet potato chlorotic stunt virus | SPCSV | (+) ssRNA | Replicase | hpRNA | [121] | ||
| sweet potato feathery mottle virus | SPFMV | (+) ssRNA | |||||
| potato virus X | PVX | (+) ssRNA | Coat protein | hpRNA | 600 bp: 180 bp (PVX), 240 bp (PVY), 180 bp (PVS) | [122] | |
| potato virus Y | PVY | (+) ssRNA | |||||
| potato virus S | PVS | (+) ssRNA | |||||
| potato virus Y | PVY | (+) ssRNA | CP gene PVY | hpRNA | 600 bp: 200 bp for each target | [102] | |
| potato virus A | PVA | (+) ssRNA | Cylindrical inclusion body PVA | ||||
| potato leaf roll virus | PLRV | (+) ssRNA | CP gene PLRV | ||||
| potato virus Y | PVY | (+) ssRNA | Coat protein | hpRNA | 605 bp | [123] | |
| Cucurbitacea | Virus | Type | Target gene | RNA Type | Construct Length | Reference | |
| Melon | zucchini yellow mosaic virus | ZYMV | (+) ssRNA | Coat protein | antisense | [124] | |
| papaya ringspot virus type W | PRSV-W | (+) ssRNA | Coat protein | hpRNA | [125] | ||
| zucchini yellow mosaic virus cucumber vein yellowing virus | ZYMV CVYV | (+) ssRNA | Cm-eIF4E translation initiation factors (eIF) | hpRNA (pHANNIBAL) | 175 bp | [90] | |
| melon necrotic spot virus | NSV | (+) ssRNA | |||||
| Moroccan watermelon mosaic virus | MWMV | (+) ssRNA | |||||
| (+) ssRNA | |||||||
| Cucumber and Melon | zucchini yellow mosaic virus | ZYMV | (+) ssRNA | VSR HC-Pro | hpRNA (pHANNIBAL) | 657 bp | [126] |
| Watermelon | watermelon silver mottle virus | WSMoV | (+) ssRNA | Partial N gene of WSMoV fused to partial CP gene sequences of CMV, CGMMV, WMV | silencer DNA (the middle half N gene of (WSMoV)) | 449 bp CP-CMV | [127] |
| cucumber mosaic virus | CMV | (+) ssRNA | 449 bp CP-CGMMV | ||||
| cucumber green mottle mosaic virus | CGMMV | (+) ssRNA | 449 bp CP-WMV | ||||
| watermelon mosaic virus | WMV | (+) ssRNA | |||||
| Fabaceae | Virus | Type | Target gene | RNA type | Construct length | Reference | |
| Soybean | soybean dwarf virus | SbDV | (+) ssRNA | Coat protein | hpRNA | 602 bp | [128] |
| alfalfa mosaic virus | AMV | (+) ssRNA | highly conserved sequences of one virus | Short dsRNA stems | 109 bp (AMV) | [129] | |
| bean pod mottle virus | BPMV | (+) ssRNA | 147 bp (BPMV) | ||||
| soybean mosaic virus | SMV | (+) ssRNA | 123 bp (SMV) | ||||
| soybean mosaic virus | SMV | (+) ssRNA | Coat protein | hpRNA | [130] | ||
| soybean mosaic virus | SMV | (+) ssRNA | VSR HC-Pro | hpRNA | [131] | ||
| soybean mosaic virus | SMV | (+) ssRNA | VSR HC-Pro | hpRNA | 268 bp | [132] | |
| soybean mosaic virus | SMV | (+) ssRNA | P3 cistron | hpRNA | 302 bp | [133] | |
| Common bean | bean golden mosaic virus | BGMV | (+) ssRNA | Replicase (AC1) | hpRNA | 411 bp | [134] |
| Cowpea | cowpea severe mosaic virus | CPSMV | (+) ssRNA | proteinase cofactor (CPSMV) Coat protein (CABMV) | hpRNA | 899 bp (415 bp RNA1- CPSMV and 462 bp CP-CABMV) | [135] |
| cowpea aphid-borne mosaic virus | CABMV | (+) ssRNA | |||||
| White clover | white clover mosaic virus | WCMV | (+) ssRNA | Replicase | sense, antisense and hpRNA | 790 bp | [136] |
| Poaceae | Virus | Type | Target gene | RNA type | Construct length | Reference | |
| Rice | rice tungro bacilliform virus | RTBV | dsDNA | ORF IV | dsRNA | 1326 bp | [137] |
| rice dwarf virus | RDV | dsRNA | non-structural protein Pns12 and Pns4 | hpRNA | 500 bp: Pns12-12N | [138] | |
| 499 bp: Pns12-12C | |||||||
| 500 bp: Pns4-4N | |||||||
| 504 bp: Pns4-4M | |||||||
| rice stripe virus | RSV | (−) ssRNA | Coat protein and special-disease protein (SP) | hpRNA | 300 bp (SP); 450 bp (CP) | [139] | |
| 700 bp (SP/CP; CP/SP) | |||||||
| rice stripe virus | RSV | (−) ssRNA | Nucleocapsid (pC3), MP (pC4), glycoprotein (pC2), non-structural protein (p4) | hpRNA | 500 bp (for each target) | [140] | |
| rice black streaked dwarf virus | RBSDV | dsRNA | nonstructural Pns9 protein | hpRNA | 500 bp | [141] | |
| rice grassy stunt virus | RGSV | (−) ssRNA | nucelocapsid protein pC5; movement protein pC6 | dsRNA | 500 bp (for each target) | [142] | |
| Wheat | wheat streak mosaic virus | WSMV | (−) ssRNA | nuclear inclusion protein ‘a’ (NIa) gene | hpRNA (pSTARGATE) | 696 bp | [143] |
| wheat streak mosaic virus | WSMV | (−) ssRNA | Conserved region of WSMV genome | amiRNA | Precursor miR395 | [144] | |
| Maize | maize dwarf mosaic virus | MDMV | (+) ssRNA | Coat protein | hpRNA | 404 bp | [145] |
| Barley | barley yellow dwarf virus | BYDV | (+) ssRNA | Polymerase gene | hpRNA | 1600 bp | [146] |
| Euphorbiaceae | Virus | Type | Target gene | RNA type | Construct length | Reference | |
| Cassava | African cassava mosaic virus | ACMV | ssDNA | Rep (AC1); TrAP (AC2), REn (AC3) | antisense construct | Full-length of each target gene | [147] |
| African cassava mosaic virus | ACMV | ssDNA | bidirectional promoter of ACMV DNA-A | intron-containing dsRNA | 256 bp | [148] | |
| African cassava mosaic virus | ACMV | ssDNA | Rep (AC1) | hpRNA | 154 bp | [149] | |
| Sir Lankan cassava mosaic virus | (SLCMV) | ssDNA | AV1 and AV2 | hpRNA | 527 bp | [117] | |
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | p5001 construct | [150] | |
| cassava brown streak virus | (+) ssRNA | ||||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | Field trials (Chauhan et al. 2015) | [151] | |
| cassava brown streak virus | (+) ssRNA | ||||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | 525 bp | [152] | |
| cassava brown streak virus | (+) ssRNA | ||||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | 894 bp (full-length CP) | [153] | |
| cassava brown streak virus | (+) ssRNA | 401 bp (CP-NT) | |||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | Field trials (Yadav et al. 2011) | [154] | |
| cassava brown streak virus | (+) ssRNA | ||||||
| cassava brown streak Uganda virus | CBSUV CBSV | (+) ssRNA | Coat protein | hpRNA | Field trials (Yadav et al. 2011) | [155] | |
| cassava brown streak virus | (+) ssRNA | ||||||
| cassava brown streak virus | CBSV | (+) ssRNA | Coat protein | hpRNA | 895 bp CP-CBSV | [156] | |
| cassava brown streak Uganda virus | CBSUV | 898 bp CP-CBSUV | |||||
| Poinsettia | poinsettia mosaic virus | PnMV | (+) ssRNA | Coat protein and RNA-dependent RNA-polymerase | hpRNA | 500 bp each target | [157] |
| Tropical fruits | Virus | Type | Target gene | RNA type | Construct length | Reference | |
| Citrus macrophylla | citrus tristeza virus | CTV | (+) ssRNA | 3′ p23 (VSR) and 3′ UTR | hpRNA | 900 bp | [109] |
| Mexican lime | citrus tristeza virus | CTV | (+) ssRNA | 3′ p23 (VSR) and 3′ UTR | sense, antisense and hpRNA | 549 bp | [158] |
| citrus tristeza virus | CTV | (+) ssRNA | VSR p20, p23, p25 | hpRNA | 548 bp (p20); 629 bp (p23); 670 bp (p25) | [159] | |
| citrus psorosis virus | CPsV | (−) ssRNA | Coat protein and 54K | hpRNA | 372 bp CP; 436 bp 54K | [160] | |
| Sweet orange | citrus psorosis virus | CPsV | (−) ssRNA | Coat protein, 54K or 24K genes | hpRNA | 372 bp CP; 436 bp 54K; 312 bp 24K | [108] |
| Banana | banana bunchy top virus | BBTV | ssDNA | Replicase | hpRNA | CP-full-length; 651 bp partial CP sequence | [92] |
| Brassicaceae | Virus | Type | Target gene | RNA type | Construct length | Reference | |
| Arabidopsis | turnip yellow mosaic virus | TYMV | (+) ssRNA | VSR P69 (TYMV) | amiRNAs | Precursor miR159a | [161] |
| turnip mosaic virus | TuMV | (+) ssRNA | VSR HC-Pro (TuMV) | ||||
| cucumber mosaic virus | CMV | (+) ssRNA | 3′ UTR | amiRNAs | Precursor miR159a | [162] | |
| Others | Virus species | Type | Target gene | RNA type | Construct length | Reference | |
| Asteraceae | chrysanthemum virus B | CVB | (+) ssRNA | Coat protein | sense, antisense and hpRNA | 273 bp | [163] |
| Amaranthaceae | beet necrotic yellow vein virus | BNYVV | (+) ssRNA | Replicase | hpRNA | 459 bp, 589 bp and 824 bp | [164] |
| Andropogoneae | sugarcane mosaic virus | SCMV | (+) ssRNA | Coat protein | hpRNA | Not mentioned | [165] |
| sorghum mosaic virus | SrMV | (+) ssRNA | |||||
| Vitaceae | grapevine fanleaf virus | GFLV | (+) ssRNA | Coat protein | amiRNA | Precursor miR319a | [91] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gaffar, F.Y.; Koch, A. Catch Me If You Can! RNA Silencing-Based Improvement of Antiviral Plant Immunity. Viruses 2019, 11, 673. https://doi.org/10.3390/v11070673
AMA Style
Gaffar FY, Koch A. Catch Me If You Can! RNA Silencing-Based Improvement of Antiviral Plant Immunity. Viruses. 2019; 11(7):673. https://doi.org/10.3390/v11070673
Chicago/Turabian StyleGaffar, Fatima Yousif, and Aline Koch. 2019. "Catch Me If You Can! RNA Silencing-Based Improvement of Antiviral Plant Immunity" Viruses 11, no. 7: 673. https://doi.org/10.3390/v11070673
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.