
Respiratory Syncytial Virus G Protein Sequence Variability among Isolates from St. Petersburg, Russia, during the 2013–2014 Epidemic Season

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Samples
2.2. Isolation of RSV in Cell Culture
2.3. RT-PCR of the G Protein Gene
2.4. Capillary Sequencing
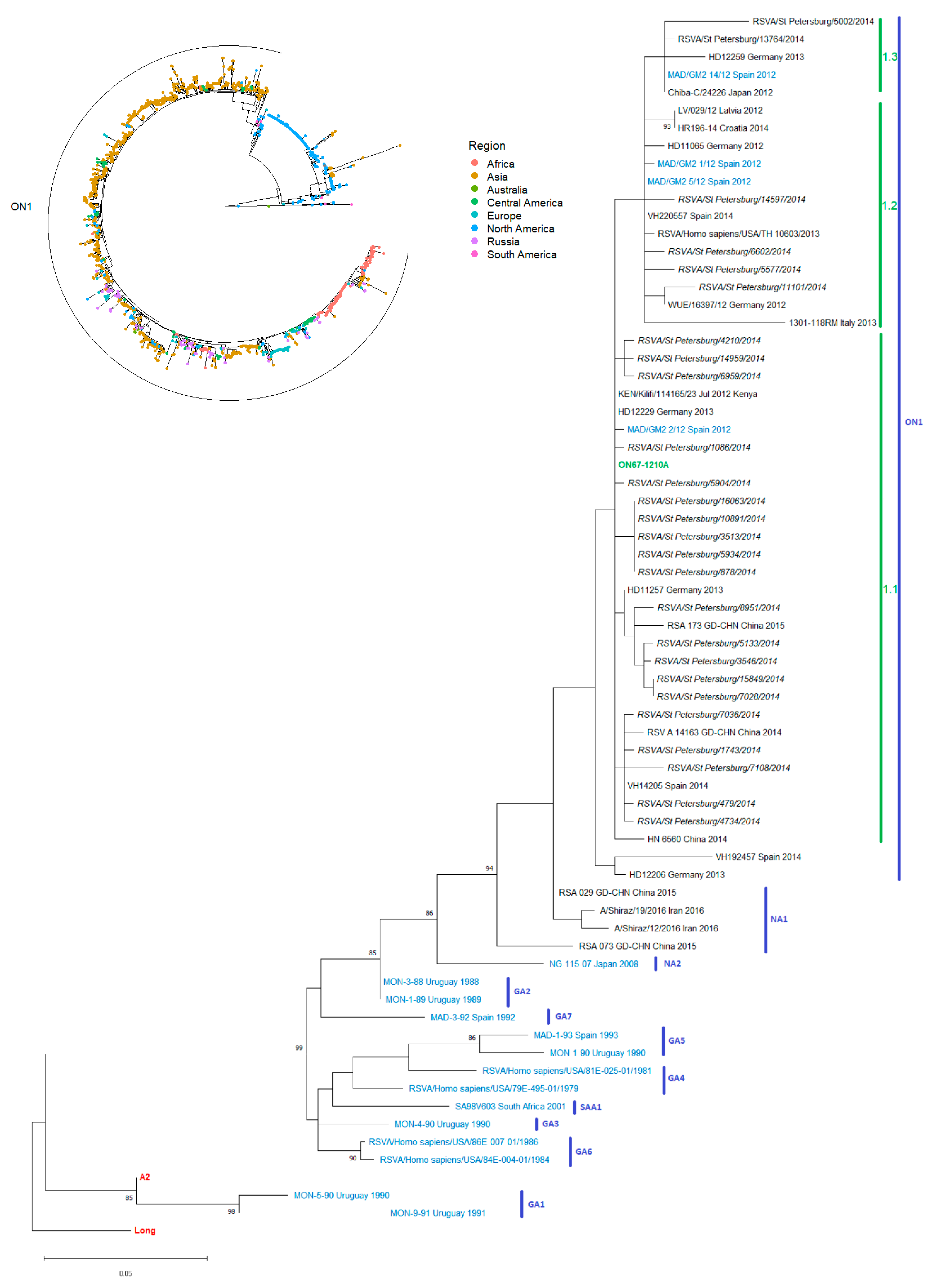
2.5. Phylogenetic Analysis of the G Gene (Nucleic Acid) Sequences
2.6. RSV Glycosylation Site Analysis
2.7. Statistical Analysis
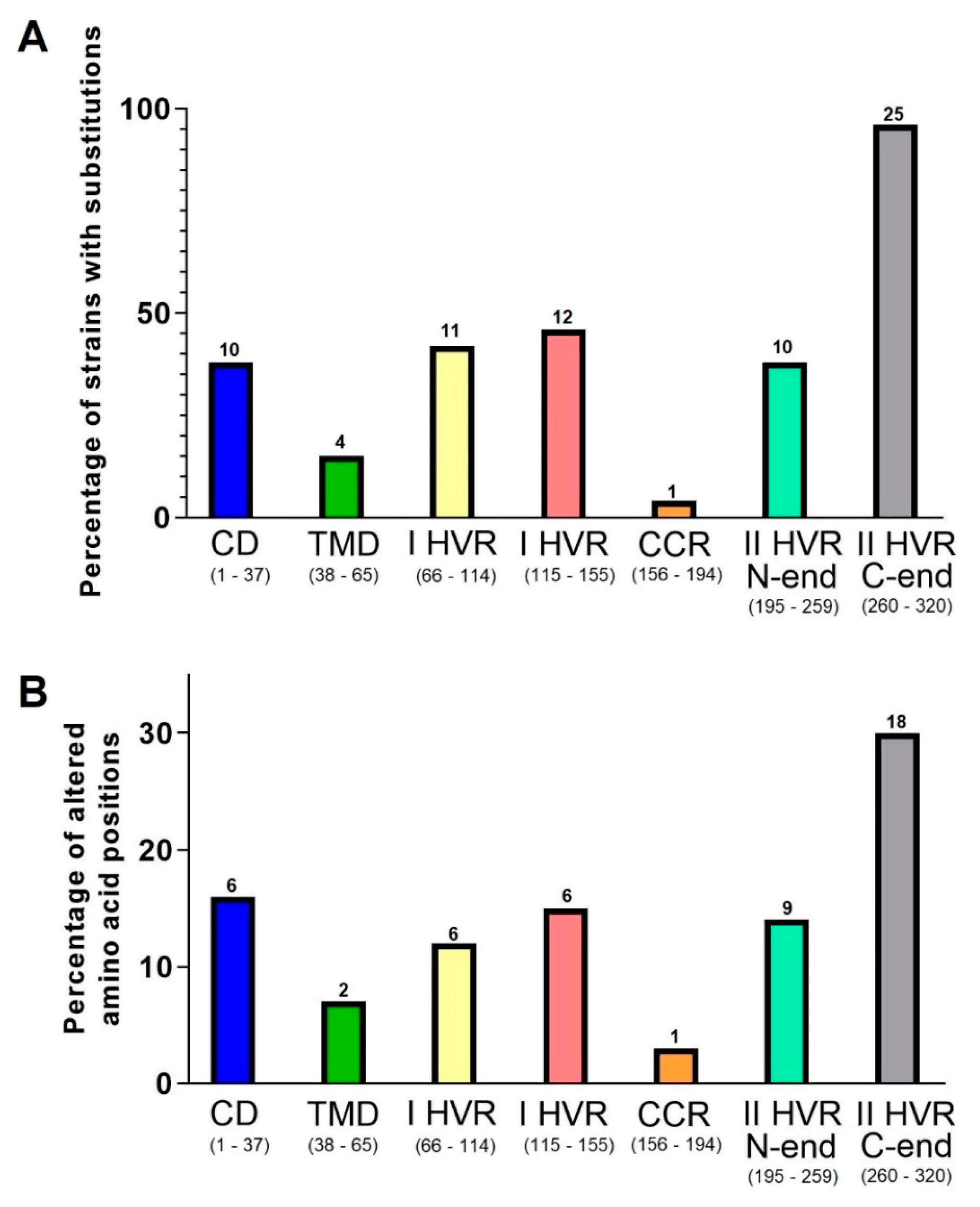
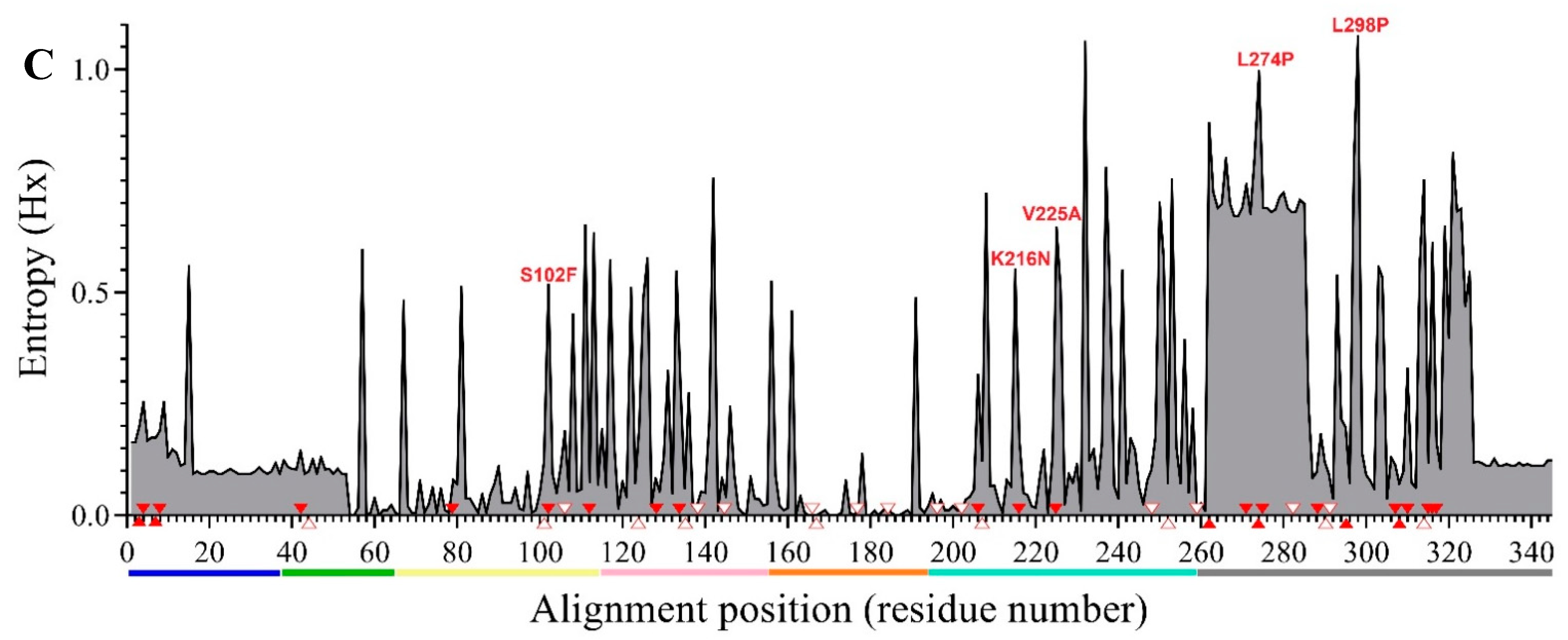
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bont, L.; Checchia, P.A.; Fauroux, B.; Figueras-Aloy, J.; Manzoni, P.; Paes, B.; Simões, E.A.; Carbonell-Estrany, X. Defining the Epidemiology and Burden of Severe Respiratory Syncytial Virus Infection Among Infants and Children in Western Countries. Infect. Dis. Ther. 2016, 5, 271–298. [Google Scholar] [CrossRef] [Green Version]
- Manzoni, P.; Figueras-Aloy, J.; Simões, E.A.F.; Checchia, P.A.; Fauroux, B.; Bont, L.; Paes, B.; Carbonell-Estrany, X. Defining the Incidence and Associated Morbidity and Mortality of Severe Respiratory Syncytial Virus Infection Among Children with Chronic Diseases. Infect. Dis. Ther. 2017, 6, 383–411. [Google Scholar] [CrossRef] [Green Version]
- Simoes, E.A.; Carbonell-Estrany, X. Impact of severe disease caused by respiratory syncytial virus in children living in developed countries. Pediatr. Infect. Dis. J. 2003, 22, S13–S18, discussion S18–S20. [Google Scholar] [CrossRef] [PubMed]
- Nair, H.; Nokes, D.J.; Gessner, B.D.; Dherani, M.; Madhi, S.A.; Singleton, R.J.; O’Brien, K.L.; Roca, A.; Wright, P.F.; Bruce, N.; et al. Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: A systematic review and meta-analysis. Lancet 2010, 375, 1545–1555. [Google Scholar] [CrossRef] [Green Version]
- Karron, R.A. Vaccines, 6th ed.; Plotkin, S.A., Orenstein, W.A., Offit, P.A., Eds.; Elsevier Inc. Saunders: Philadelphia, PA, USA, 2012; pp. 1146–1153. [Google Scholar]
- Sominina, A.A.; Pisareva, M.M.; Buzitskaya, Z.V.; Osidak, L.V.; Sukhovetskaya, V.F.; Afanasyeva, O.I.; Voloshchuk, L.V.; Afanas’eva, V.S.; Obraztsova, E.V.; Golovacheva, E.G.; et al. Peculiarities of Etiology of Respiratory Virus Infections in Hospitalized Patients Depending on the Demographic, Socio-Economic Factors and Previous Vaccination. Epidemiol. I Vakcinoprofilaktika 2015, 14, 74–83. (In Russian) [Google Scholar] [CrossRef] [Green Version]
- Anderson, L.J.; Hierholzer, J.C.; Tsou, C.; Hendry, R.M.; Fernie, B.F.; Stone, Y.; McIntosh, K. Antigenic Characterization of Respiratory Syncytial Virus Strains with Monoclonal Antibodies. J. Infect. Dis. 1985, 151, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Zhu, R.; Qian, Y.; Deng, J.; Zhao, L.; Sun, Y.; Wang, F. Genetic Variation in Attachment Glycoprotein Genes of Human Respiratory Syncytial Virus Subgroups A and B in Children in Recent Five Consecutive Years. PLoS ONE 2013, 8, e75020. [Google Scholar] [CrossRef]
- Nagasawa, K.; Hirano, E.; Kobayashi, M.; Ryo, A.; Oishi, K.; Obuchi, M.; Ishiwada, N.; Noda, M.; Kuroda, M.; Shimojo, N.; et al. Molecular evolution of the hypervariable region of the attachment glycoprotein gene in human respiratory syncytial virus subgroup B genotypes BA9 and BA10. Infect. Genet. Evol. 2015, 36, 217–273. [Google Scholar] [CrossRef]
- Duvvuri, V.R.; Granados, A.; Rosenfeld, P.; Bahl, J.; Eshaghi, A.; Gubbay, J.B. Genetic diversity and evolutionary insights of respiratory syncytial virus A ON1genotype: Global and local transmission dynamics. Sci. Rep. 2015, 5, e14268. [Google Scholar] [CrossRef] [Green Version]
- Tabatabai, J.; Prifert, C.; Pfeil, J.; Grulich-Henn, J.; Schnitzler, P. Novel Respiratory Syncytial Virus (RSV) Genotype ON1 Predominates in Germany during Winter Season 2012–13. PLoS ONE 2014, 9, e109191. [Google Scholar] [CrossRef] [Green Version]
- Esposito, S.; Piralla, A.; Zampiero, A.; Bianchini, S.; Pietro, G.D.; Scala, A.; Pinzani, R.; Fossali, E.; Baldanti, F.; Principi, N. Characteristics and Their Clinical Relevance of Respiratory Syncytial Virus Types and Genotypes Circulating in Northern Italy in Five Consecutive Winter Seasons. PLoS ONE 2015, 10, e0129369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vos, L.M.; Oosterheert, J.J.; Kuil, S.D.; Viveen, M.; Bont, L.J.; Hoepelman, A.I.M.; Coenjaerts, F.E.J. High epidemic burden of RSV disease coinciding with genetic alterations causing amino acid substitutions in the RSV G-protein during the 2016/2017 season in The Netherlands. J. Clin. Virol. 2019, 112, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Bohmwald, K.; Gálvez, N.M.S.; Canedo-Marroquín, G.; Pizarro-Ortega, M.S.; Andrade-Parra, C.; Gómez-Santander, F.; Kalergis, A.M. Contribution of Cytokines to Tissue Damage During Human Respiratory Syncytial Virus Infection. Front. Immunol. 2019, 10, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorquera, P.A.; Anderson, L.; Tripp, R.A. Human Respiratory Syncytial Virus. In Methods and Protocols/Methods in Molecular Biology; Springer Science and Business Media: New York, NY, USA, 2016; Volume 1442, pp. 1–12. [Google Scholar]
- Hijano, D.R.; Vu, L.D.; Kauvar, L.M.; Tripp, R.A.; Polack, F.P.; Cormier, S.A. Role of type I interferon (IFN) in the respiratory syncytial virus (RSV) immune response and disease severity. Front. Immunol. 2019, 10, 566. [Google Scholar] [CrossRef] [Green Version]
- Cane, P.A. Analysis of linear epitopes recognised by the primary human antibody response to a variable region of the attachment (G) protein of respiratory syncytial virus. J. Med. Virol. 1997, 51, 297–304. [Google Scholar] [CrossRef]
- Kim, S.; Joo, D.H.; Lee, J.B.; Shim, B.S.; Cheon, I.S.; Jang, J.E.; Song, H.H.; Kim, K.H.; Song, M.K.; Chang, J. Dual Role of Respiratory Syncytial Virus Glycoprotein Fragment as a Mucosal Immunogen and Chemotactic Adjuvant. PLoS ONE 2012, 7, e32226. [Google Scholar] [CrossRef]
- Lee, J.; Klenow, L.; Coyle, E.M.; Golding, H.; Khurana, S. Protective antigenic sites in respiratory syncytial virus G attachment protein outside the central conserved and cysteine noose domains. PLoS Pathog. 2018, 14, e1007262. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Yu, G. Using ggtree to Visualize Data on Tree-Like Structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef]
- Charif, D.; Lobry, J. SeqinR 1.0-2: A contributed package to the R project for statistical computing devoted to biological sequences retrieval and analysis. In Structural Approaches to Sequence Evolution: Molecules, Networks, Populations, Series Biological and Medical Physics, Biomedical Engineering; Bastolla, U., Porto, M., Roman, H., Vendruscolo, M., Eds.; Springer Verlag: New York, NY, USA, 2007; pp. 207–232. ISBN 978-3-540-35305-8. [Google Scholar]
- Trento, A.; Ábrego, L.; Rodriguez-Fernandez, R.; González-Sánchez, M.I.; González-Martínez, F.; Delfraro, A.; Pascale, J.M.; Arbiza, J.; Melero, J.A. Conservation of G-Protein Epitopes in Respiratory Syncytial Virus (Group A) Despite Broad Genetic Diversity: Is Antibody Selection Involved in Virus Evolution? J. Virol. 2015, 89, 7776–7785. [Google Scholar] [CrossRef] [Green Version]
- Agoti, C.N.; Otieno, J.R.; Gitahi, C.W.; Cane, P.A.; Nokes, D.J. Rapid Spread and Diversification of Respiratory Syncytial Virus Genotype ON1, Kenya. Emerg. Infect. Dis. 2014, 20, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Korsun, N.; Angelova, S.; Tzotcheva, I.; Georgieva, I.; Lazova, S.; Parina, S.; Alexiev, I.; Perenovska, P. Prevalence and genetic characterisation of respiratory syncytial viruses circulating in Bulgaria during the 2014/15 and 2015/16 winter seasons. Pathog. Glob. Health 2017, 111, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thongpan, I.; Mauleekoonphairoj, J.; Vichiwattana, P.; Korkong, S.; Wasitthankasem, R.; Vongpun-sawad, S.; Poovorawan, Y. Respiratory syncytial virus genotypes NA1, ON1, and BA9 are prevalent in Thailand, 2012–2015. Peer. J. 2017, 5, e3970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, B.; Liu, H.; Tabor, D.E.; Tovchigrechko, A.; Qi, Y.; Ruzin, A.; Esser, M.T.; Jin, H. Emergence of new antigenic epitopes in the glycoproteins of human respiratory syncytial virus collected from a US surveillance study, 2015–17. Sci. Rep. 2019, 9, 3898. [Google Scholar]
- Ren, L.; Xia, Q.; Xiao, Q.; Zhou, L.; Zang, N.; Long, X.; Xie, X.; Deng, Y.; Wang, L.; Fu, Z.; et al. The genetic variability of glycoproteins among respiratory syncytial virus subtype A in China between 2009 and 2013. Infect. Genet. Evol. 2014, 27, 339–347. [Google Scholar] [CrossRef]
- Fall, A.; Dia, N.; Cisseel, H.A.; Kiori, D.E.; Sarr, F.D.; Sy, S.; Goudiaby, D.; Richard, V.; Niang, M.N. Epidemiology and Molecular Characterization of Human Respiratory Syncytial Virus in Senegal after Four Consecutive Years of Surveillance, 2012–2015. PLoS ONE 2016, 11, e0157163. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, S.; Coyle, E.M.; Beeler, J.; Golding, H.; Khurana, S. Antigenic Fingerprinting following Primary RSV Infection in Young Children Identifies Novel Antigenic Sites and Reveals Unlinked Evolution of Human Antibody Repertoires to Fusion and Attachment Glycoproteins. PLoS Pathog. 2016, 12, e1005554. [Google Scholar] [CrossRef]
- Walsh, E.E.; Hall, C.B.; Schlesinger, J.J.; Brandriss, M.W.; Hildreth, S.; Paradiso, P. Immunization with glycoprotein subunits of respiratory syncytial virus to protect cotton rats against viral infection. J. Gen. Virol. 1989, 70, 2953–2961. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, J.Y.; Park, M.H.; Kim, J.Y.; Chang, J. Monoclonal Antibody against G Glycoprotein Increases Respiratory Syncytial Virus Clearance In Vivo and Prevents Vaccine-Enhanced Diseases. PLoS ONE 2017, 12, e0169139. [Google Scholar] [CrossRef] [Green Version]
- Hancock, G.E.; Tebbey, P.W.; Scheuer, C.A.; Pryharski, K.S.; Heers, K.M.; LaPierre, N.A. Immune responses to the nonglycosylated ectodomain of respiratory syncytial virus attachment glycoprotein mediate pulmonary eosinophilia in inbred strains of mice with different MHC haplotypes. J. Med. Virol. 2003, 70, 301–308. [Google Scholar] [CrossRef]
- Johnson, T.R.; Varga, S.M.; Braciale, T.J.; Graham, B.S. V14β T Cells Mediate the Vaccine-Enhanced Disease Induced by Immunization with Respiratory Syncytial Virus (RSV) G Glycoprotein but Not with Formalin-Inactivated RSV. J. Virol. 2004, 78, 8753–8760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Mason, C.S.; Jones, L.P.; Crabtree, J.; Jorquera, P.A.; Tripp, R.A. Antibodies to the Central Conserved Region of Respiratory Syncytial Virus (RSV) G Protein Block RSV G Protein CX3C-CX3CR1 Binding and Cross-Neutralize RSV A and B Strains. Viral Immunol. 2012, 25, 193–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, B.; Chirkova, T.; Boukhvalova, M.S.; Sun, H.Y.; Walsh, E.E.; Anderson, C.S.; Mariani, T.J.; An-derson, L.J. Mutation of Respiratory Syncytial Virus G Protein’s CX3C Motif Attenuates Infection in Cotton Rats and Primary Human Airway Epithelial Cells. Vaccines 2019, 7, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparer, T.E.; Matthews, S.; Hussell, T.; Rae, A.J.; Garcia-Barreno, B.; Malero, J.A.; Openshaw, P. Eliminating a region of respiratory syncytial virus attachment protein allows induction of protective immunity without vaccine-enhanced lung eosinophilia. J. Exp. Med. 1998, 187, 1921–1926. [Google Scholar] [CrossRef] [Green Version]
- Tebbey, P.W.; Hagen, M.; Hancock, G.E. Atypical pulmonary eosinophilia is mediated by a specific amino acid sequence of the attachment (G) protein of respiratory syncytial virus. J. Exp. Med. 1998, 188, 1967–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cane, P.A.; Pringle, C.R. Evolution of subgroup A respiratory syncytial virus: Evidence for progressive accumulation of amino acid changes in the attachment protein. J. Virol. 1995, 69, 2918–2925. [Google Scholar] [CrossRef] [Green Version]
- Shields, B.; Mills, J.; Ghildyal, R.; Gooley, P.; Meanger, J. Multiple heparin binding domains of respiratory syncytial virus G mediate binding to mammalian cells. Arch. Virol. 2003, 148, 1987–2003. [Google Scholar] [CrossRef]
- Elliott, M.B.; Pryharski, K.S.; Yu, Q.; Boutilier, L.A.; Campeol, N.; Melville, K.; Laughlin, T.S.; Gupta, C.K.; Lerch, R.A.; Randolph, V.B.; et al. Characterization of Recombinant Respiratory Syncytial Viruses with the Region Responsible for Type 2 T-Cell Responses and Pulmonary Eosinophilia Deleted from the Attachment (G) Protein. J. Virol. 2004, 78, 8446–8454. [Google Scholar] [CrossRef] [Green Version]
- Eshaghi, A.; Duvvuri, V.R.; Lai, R.; Nadarajah, J.T.; Li, A.; Patel, S.N.; Low, D.E.; Gubbay, J.B. Genetic Variability of Human Respiratory Syncytial Virus A Strains Circulating in Ontario: A Novel Genotype with a 72 Nucleotide G Gene Duplication. PLoS ONE 2012, 7, e32807. [Google Scholar] [CrossRef] [Green Version]
- Collins, P.L.; Chanock, R.M.; Murphy, B.R. Fields Virology, 4th ed.; Fields, B.N., Howley, P.M., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2001; Volume 1, pp. 1182–1215. [Google Scholar]
- Collins, P.L.; Karron, R.A. Fields Virology, 6th ed.; Fields, B.N., Knipe, D.M., How-Ley, P.M., Eds.; Wolters Kluwer Health; Lippincott Williams, and Wilkins: Philadelphia, PA, USA, 2013; Volume 1, pp. 1086–1123. [Google Scholar]
- Melero, J.A.; Mas, V.; McLellan, J.S. Structural, antigenic and immunogenic features of respiratory syncytial virus glycoproteins relevant for vaccine development. Vaccines 2017, 35, 461–468. [Google Scholar] [CrossRef] [Green Version]
- Botosso, V.F.; Zanotto, P.M.D.A.; Ueda, M.; Arruda, E.; Gilio, A.E.; Vieira, S.E.; Stewien, K.E.; Peret, T.C.T.; Jamal, L.F.; Pardini, M.I.D.M.C.; et al. Positive Selection Results in Frequent Reversible Amino Acid Replacements in the G Protein Gene of Human Respiratory Syncytial Virus. PLoS Pathog. 2009, 5, e1000254. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Yi, L.; Wu, J.; Song, Y.; Huang, G.; Zhang, X.; Liang, L.; Ni, H.; Pybus, O.G.; Ke, C.; et al. Evolution and Transmission of Respiratory Syncytial Group A (RSV-A) Viruses in Guangdong, China 2008–2015. Front. Microbiol. 2016, 7, e1263. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| M13F-hRSVAB-G-F | 5′-TGTAAAACGACGGCCAGTGCAAATGCAAACATGTCCAAA-3′ |
| M13R-hRSVA-G-R | 5′-CAGGAAACAGCTATGCAACYATACGCTTTTTAAATGACTA-3′ |
| Strain | GenBank Accession No. | Strain | GenBank Accession No. |
|---|---|---|---|
| ON67-1210A | JN257693 | MON-1-89 | Z33422 |
| ATCC VR-26 (Long) | AY911262 | MON-5-90 | Z33427 |
| A2 | KT992094 | MON-1-90 | Z33494 |
| RSVA/Homo_sapiens/USA/79E-495-01/1979 | KP856969 | MON-4-90 | Z33426 |
| RSVA/Homo_sapiens/USA/81E-025-01/1981 | KP856967 | MON-9-91 | Z33431 |
| RSVA/Homo_sapiens/USA/84E-004-01/1984 | KP258733 | MAD-3-92 | Z33455 |
| RSVA/Homo_sapiens/USA/86E-007-01/1986 | KP258723 | VH220557 | KM402663 |
| MAD-1-93 | Z33414 | HN_6560 | KT781349 |
| MAD/GM2_14/12 | KP792373 | RSV_A_14163_GD-CHN_2014 | KX009698 |
| MAD/GM2_1/12 | KP792361 | RSA_073_GD-CHN_2015 | KX009700 |
| MAD/GM2_2/12 | KP792362 | RSA_029_GD-CHN_2015 | KX009699 |
| MAD/GM2_5/12 | KP792365 | RSA_173_GD-CHN_2015 | KX009686 |
| RSVA/Homo_sapiens/USA/TH_10603/2013 | KU950596 | A/Shiraz/12/2016 | KU716108 |
| MON-3-88 | Z33425 | A/Shiraz/19/2016 | KU716109 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krivitskaya, V.; Komissarova, K.; Pisareva, M.; Sverlova, M.; Fadeev, A.; Petrova, E.; Timonina, V.; Sominina, A.; Danilenko, D. Respiratory Syncytial Virus G Protein Sequence Variability among Isolates from St. Petersburg, Russia, during the 2013–2014 Epidemic Season. Viruses 2021, 13, 119. https://doi.org/10.3390/v13010119
Krivitskaya V, Komissarova K, Pisareva M, Sverlova M, Fadeev A, Petrova E, Timonina V, Sominina A, Danilenko D. Respiratory Syncytial Virus G Protein Sequence Variability among Isolates from St. Petersburg, Russia, during the 2013–2014 Epidemic Season. Viruses. 2021; 13(1):119. https://doi.org/10.3390/v13010119
Chicago/Turabian StyleKrivitskaya, Vera, Kseniya Komissarova, Maria Pisareva, Maria Sverlova, Artem Fadeev, Ekaterina Petrova, Veronika Timonina, Anna Sominina, and Daria Danilenko. 2021. "Respiratory Syncytial Virus G Protein Sequence Variability among Isolates from St. Petersburg, Russia, during the 2013–2014 Epidemic Season" Viruses 13, no. 1: 119. https://doi.org/10.3390/v13010119