
A Review of Potential Therapeutic Strategies for COVID-19
by
Jiajia Meng
1,†,
Ruijiao Li
1,2,†,
Zhiqi Zhang
1,
Jie Wang
1,2,
Qingwen Huang
1,
Dongxia Nie
1,
Kai Fan
1,
Wenbo Guo
1,
Zhihui Zhao
1 and
Zheng Han
1,2,* 1
Institute for Agro-Food Standards and Testing Technology, Shanghai Academy of Agricultural Sciences, Shanghai 201403, China
2
School of Health Science and Engineering, University of Shanghai for Science and Technology, Shanghai 200093, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this study.
Viruses 2022, 14(11), 2346; https://doi.org/10.3390/v14112346
Submission received: 18 September 2022
/
Revised: 21 October 2022
/
Accepted: 23 October 2022
/
Published: 25 October 2022
(This article belongs to the Section SARS-CoV-2 and COVID-19)
Abstract
:Coronavirus disease 2019 is a rather heterogeneous disease caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). The ongoing pandemic is a global threat with increasing death tolls worldwide. SARS-CoV-2 belongs to lineage B β-CoV, a subgroup of Sarbecovirus. These enveloped, large, positive-sense single-stranded RNA viruses are easily spread among individuals, mainly via the respiratory system and droplets. Although the disease has been gradually controlled in many countries, once social restrictions are relaxed the virus may rebound, leading to a more severe and uncontrollable situation again, as occurred in Shanghai, China, in 2022. The current global health threat calls for the urgent development of effective therapeutic options for the treatment and prevention of SARS-CoV-2 infection. This systematic overview of possible SARS-CoV-2 therapeutic strategies from 2019 to 2022 indicates three potential targets: virus entry, virus replication, and the immune system. The information provided in this review will aid the development of more potent and specific antiviral compounds.
1. Introduction
Coronavirus disease 2019 (COVID-19) is caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). It has spread worldwide and become a major health crisis [1,2]. Patients infected with SARS-CoV-2 display major symptoms that include fever, dyspnea, cough, and sore throat, while some patients show minor symptoms, such as dysgeusia, anosmia, gastrointestinal symptoms, headache, and skin lesions [3,4,5,6]. The condition of some patients may subsequently deteriorate rapidly, with multiple organ failure or even death within a short time [7]. Data regarding predictors of mortality in patients with COVID-19 are still scarce but are being actively investigated [8]. Person-to-person contact and respiratory droplets are the two major routes of transmission of COVID-19 infection to humans [9]. Angiotensin-converting enzyme 2 (ACE2) presented on the surface of alveolar cells in the lungs is the receptor for SARS-CoV-2 cell entry. ACE2 is also found on the mucosal cells of intestines, tubular epithelial cells of kidneys, and epithelial cells of renal tubules, which is presented as a variety of susceptible targets to SARS-CoV-2 infection [10,11]. Several studies suggested that patients with COVID-19 who use angiotensin-converting enzyme inhibitors/angiotensin receptor blockers had an increased risk of respiratory failure and death, but further randomized trials are needed to answer the question [12,13]. To date, COVID-19 has been reported from more than 200 countries; this human health threat has become the focus of global attention [14]. Actually, more than six hundred million cases have been reported with more than 6 million deaths [15].
The widespread occurrence of SARS-CoV-2 has been gradually controlled in many countries. However, once social restrictions are relaxed, the virus may rebound. This can lead to a more severe and uncontrollable situation again, such as has occurred in Shanghai, China, in 2022 [16,17]. Understanding the pathogenesis and potential targets of the virus to prevent SARS-CoV-2 infections is a global research priority. Numerous medicines have been investigated for the treatment of COVID-19 in previous studies in different parts of the world. Molnupiravir, nirmatrelvir/ritonavir, and remdesivir have been recognized as promising antiviral agents to manage COVID-19 and have been approved in different parts of the world. Moreover, some monoclonal antibodies, such as sotrovimab, casirivimab/imdevimab (Ronapreve®), and tixagevimab/cilgavimab (Evusheld®), have shown effective inhibition against COVID-19 and have permitted emergency use authorization and approval in many countries [18,19,20]. However, the benefit of these drugs is still a matter of debate due to less safety and efficacy and the variants of SARS-CoV-2.
This paper comprehensively reviews the effective recent therapeutic strategies proposed for the treatment and prevention of SARS-CoV-2 infections. Our aim is to provide insights into the development of potent and specific antivirals.
2. Infection Mechanisms of SARS-CoV-2
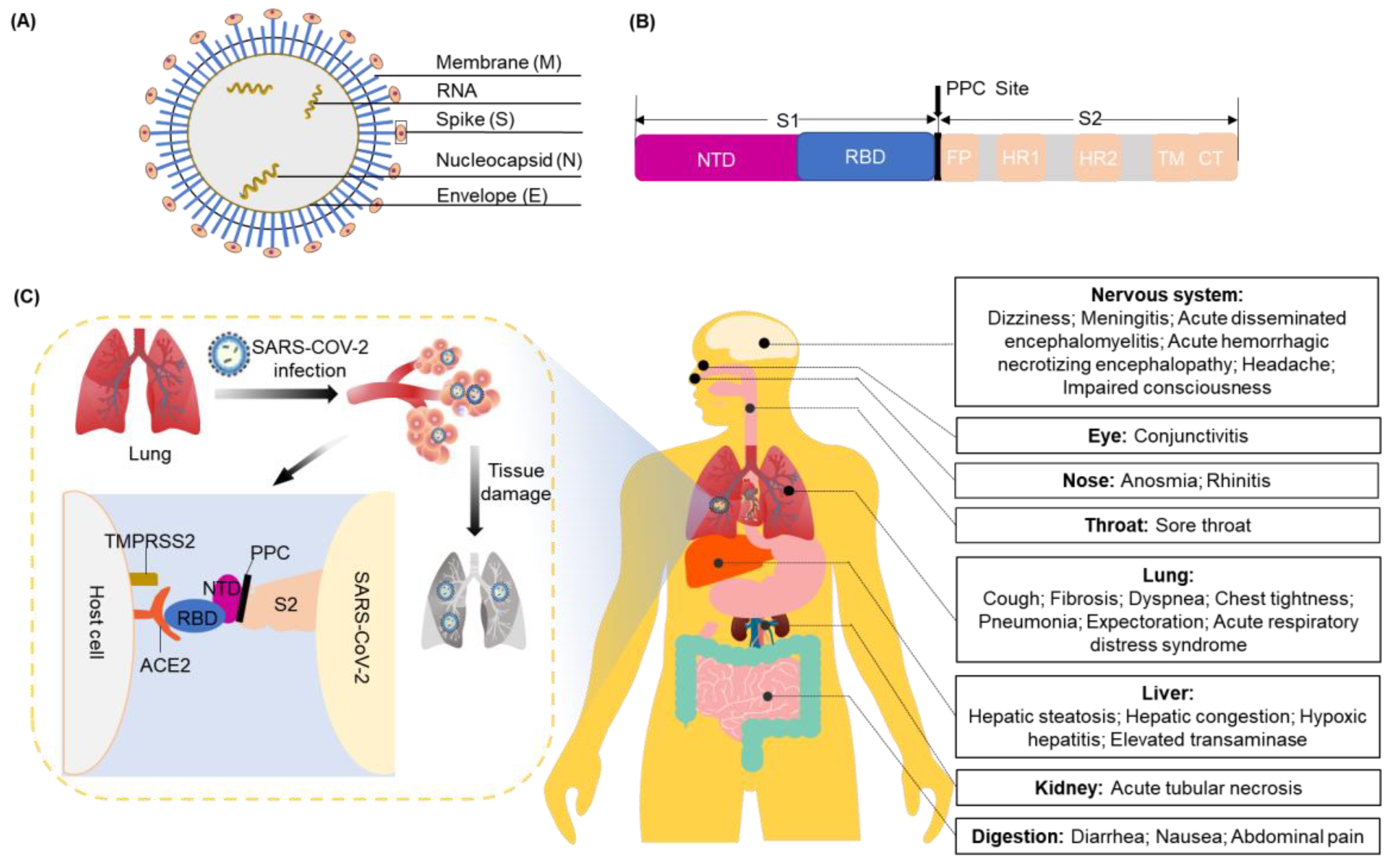
The coronavirus (CoV) genome ranges in length from 26 to 32 kilobases. It is the largest known viral RNA genome. Until now, seven types of CoVs that can cause fatal respiratory tract infections have been identified [21]. Among these, two are classified as α-CoV (229E and NL63) and five as β-CoV (OC43, HKU1, SARS-CoV, SARS-CoV-2, and Middle East Respiratory Syndrome (MERS)-CoV) [22]. SARS-CoV-2 belongs to lineage B of the β-CoVs, a subgroup of Sarbecovirus. SARS-CoV-2 is an enveloped, large, and positive-sense single-stranded RNA virus. It shares an 80% sequence identity with SARS-CoV and a 50% sequence identity with MERS-CoV. The genome of SARS-CoV-2 comprises 14 open reading frames that encode 27 proteins. These include major structural proteins: the spike (S) protein, membrane (M) protein, envelope (E) protein, and nucleocapsid (N) protein (Figure 1A).
ACE2 is responsible for allowing viral entry into cells via interactions with the S protein [23]. The S protein includes the S1 and S2 subunits, with the receptor-binding domain (RBD) on the tip of the S1 subunit. The proprotein convertase (PPC) site is discovered through the S-protein gene sequence of SARS-CoV-2, and it is the cutting point between S1 and S2 of SARS-CoV-2 (Figure 1B) [24]. Receptor binding induces the dissociation of S1 and ACE2, which elongates the structure of the S2 subunit so that the fusion peptide on it attaches to the host cell from the sub-fusion state to the fusion state. This sequence of events drives human infection. Another cellular factor important for viral entry is transmembrane protease serine 2 (TMPRSS2), which can cleave the protease ADM117 of ACE2 and activate the S protein for membrane fusion [25]. ACE2 and TMPRSS2 are mainly expressed in epithelial cells. Of these lung cells, 6.7% of type II cells express ACE2 and 3.8% co-express ACE2 and TMPRSS2 [26].
SARS-CoV-2 uses ACE2 as its receptor and mainly infects ciliated bronchial epithelial cells and lung cells [27]. When SARS-CoV-2 enters the respiratory tract, it irritates and damages tracheal mucosal cells. The resulting symptoms include coughing, fevering, fatiguing, etc. Along with the severe infections, especially after the gradual damage to the bronchioles and alveoli, the surrounding connective tissues can also be damaged due to inflammation. This can result in the leak of a large amount of protein-rich inflammatory exudate into the alveoli, occupying the space that normally holds air. Consequently, a carbon dioxide and oxygen exchange is difficult in the limited space, directly leading to difficulty in breathing and serious and life-threatening damage to the liver, kidney, nervous system, and other organs (Figure 1C) [28,29,30,31,32,33,34,35].
In the initial stage, the condition of SARS-CoV-2 patients is not serious. Later, failure of multiple organs may occur, and the severity of the disease may be related to the cytokine storm [36]. The specific molecules involved are still unclear.
3. Potential Therapeutic Strategies
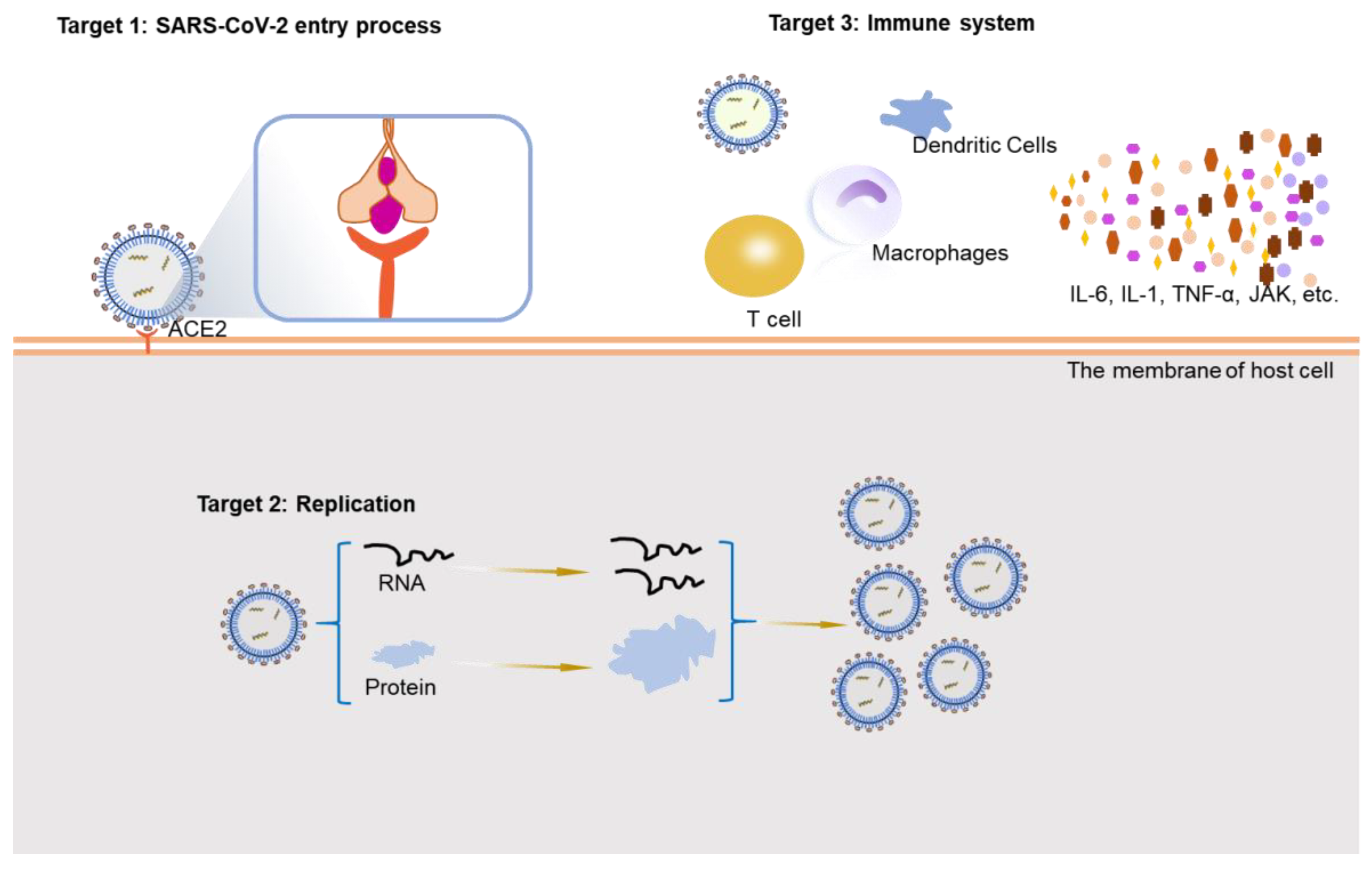
COVID-19 is a public health emergency of global concern. A large number of SARS-CoV-2-infected patients exhibit a mild-to-moderate illness, about 5–10% of the infections occur serious and life-threatening cases, and the mortality rate is about 2% [37]. The number of new confirmed patients continues to increase, highlighting the importance of the development of rapid and effective therapeutic strategies by the design of new specific drugs or the repurposing of existing drugs [38]. Herein, we summarize the therapeutic effects of the existing drugs and substances from three aspects based on potential targets: virus entry, virus replication, and the immune system (Figure 2). We also update the latest research on potential therapeutic strategies to provide new methods and ideas for future treatment methods development.
3.1. Entry of SARS-CoV-2
The S protein of SARS-CoV-2 binds to ACE2 in host cells to form the S protein–ACE2 complex. The complex is internalized by endocytosis and facilitates the entry of each virion into the cytoplasm. Inhibiting the binding of the S protein and ACE2 receptor to block the entry of SARS-CoV-2 into the host cells is an attractive approach.
3.1.1. S Protein of SARS-CoV-2
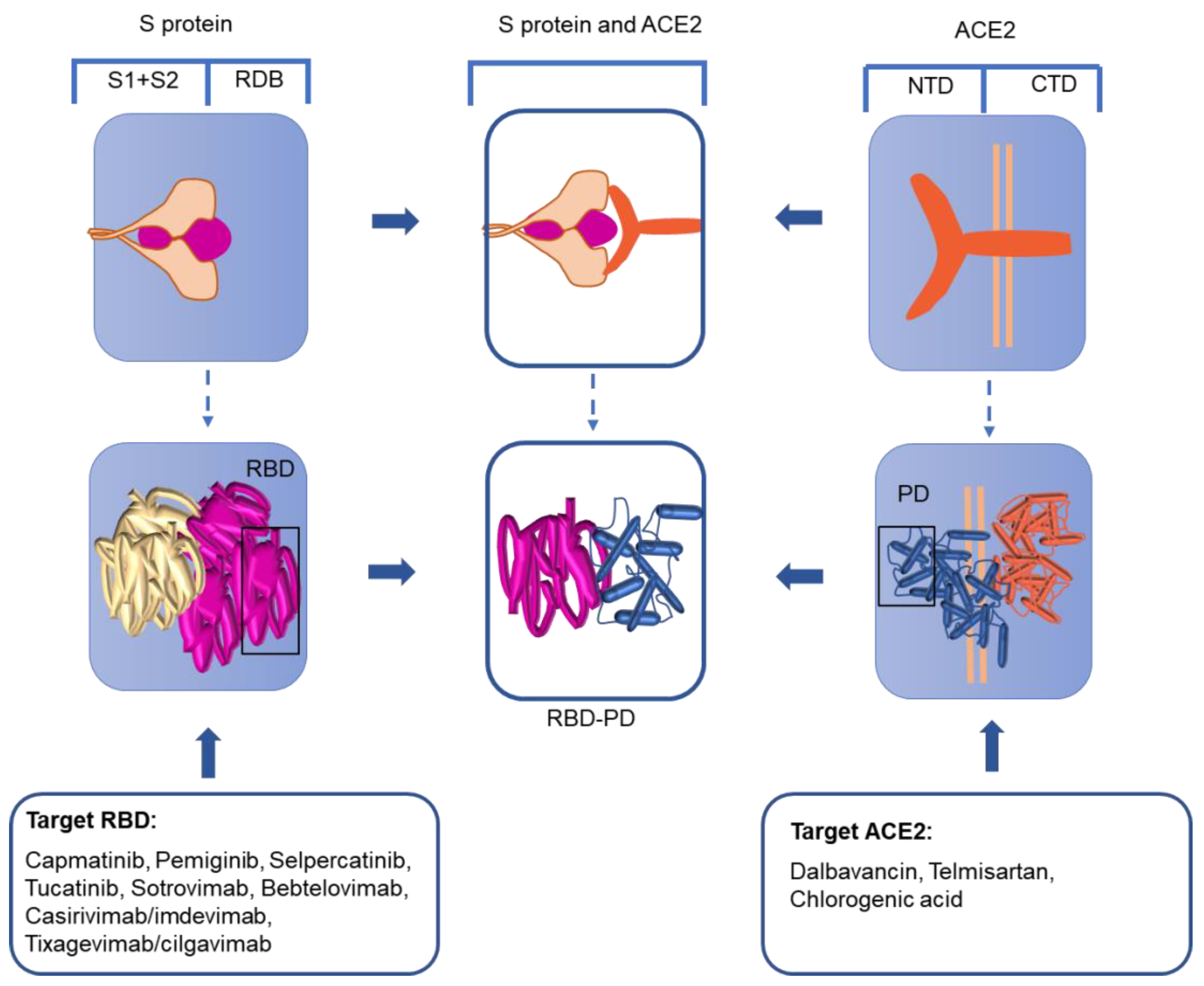
The trimer-shaped S protein with its’ S1 and S2 subunits on the surface of the virus harbors an RBD in the S-protein fusion active center [39]. When the S protein is fused, it undergoes structural reorganization with the RBD moving upward, leaving the area for receptor binding [40]. In this process, the S1 subunit falls off and the S2 subunit adopts the fused state [41,42,43]. Therefore, based on the idea that the S-protein RBD is the key binding site in human infection, a variety of strategies for the treatment of SARS-CoV-2 have been proposed (Figure 3).
Anticancer drugs may be beneficial for the treatment of SARS-CoV-2. Of the Food and Drug Administration (FDA)-approved anticancer drugs, four drugs (capmatinib, pemiginib, selpercatinib, and tucatinib) completely dock with the S protein of SARS-CoV-2. The docking prevents receptor binding, preventing virus infection [44].
Monoclonal antibodies (mAbs) therapy is another efficient approach for the treatment of SARS-CoV-2, given the 17 residue positions for antibodies that have been discovered in the RDB [45]. Various monoclonal antibodies have been screened for the RDB target of the S protein [46,47,48]. Sotrovimab is a recombinant human monoclonal immunoglobulin GI antibody that targets RBD epitopes outside the receptor-binding motif [49,50]. A COMET-ICE clinical trial demonstrated an 85% reduction in the primary endpoint of hospitalizations (more than 24 h) or death for those receiving sotrovimab compared to placebo. On 26 May 2021, sotrovimab was approved by the FDA for the treatment of mild and moderate symptoms of CoV-2 in people 12 years and older [51]. However, the efficacy of sotrovimab for the different new mutant strains would remain to be studied. Recently, the Omicron variant has become the dominant variant of SARS-CoV-2 globally, and bebtelovimab, which was shown to be effective against the different Omicron subvariants, was authorized in adults, those aged 12 years or older, and pediatric patients [52,53]. The emergence of virus-escape mutants and the development of antibody resistance have made the combined use of two or more antiviral monoclonal antibodies against different epitopes a critical aspect to improve efficacy. Casirivimab/imdevimab (REGN-COV2, Ronapreve), a cocktail of human antibodies against the S-protein RBD of SARS-CoV-2, was first authorized by the FAD for emergency use to treat patients with mild-to-moderate COVID-19 [54,55]. Evusheld (tixagevimab/cilgavimab), isolated from COVID-19 patients, can bind to different locations of the receptor-binding region of the SARS-CoV-2 S protein to neutralize SARS-CoV-2 [56,57]. These mAbs have indicated high effectiveness in trial studies with a decrease of 70–85% in hospitalization or death [58].
Nanobodies, which are composed of single-heavy-chain dimers, were first discovered in the serum of camels [59]. They are smaller, more soluble, stable, and permeable, with a higher affinity compared to traditional antibodies [59,60]. Nanobodies in the alpaca heavy-chain antibody library can bind to the RBD of the S protein, as demonstrated by surface plasmon resonance. The binding site is the same as that of ACE2. A stable nanobody–RDB complex is generated via a hydrogen bond or bivalent salt linkage with RBD residues. The complex can neutralize SARS-CoV-2 and has preventive and therapeutic effects on virus infection [61,62]. Wu et al. prepared a SARS-CoV-2 nanobody that can simultaneously bind to the RBD to block human infection [63].
An aptamer is a single-stranded oligonucleotide. The oligonucleotide is small, safe, and has been widely used to treat COVID-19. The ideal aptamer has excellent structural dynamics and blocking behavior, ensuring optimal affinity to the RBD of SARS-CoV-2. The prevention and treatment prowess of the aptamer rely on its ability to compete with ACE2. Aptamer CoV2-6 can compete and substitute with ACE2 in binding to the S-protein RBD. To improve the stability, affinity, and inhibition efficacy, CoV2-6 was further shortened and engineered as a circular bivalent aptamer, termed CoV2-6C3 [64]. Other oligonucleotide aptamers containing a conserved sequence motif that can block the interaction of the RBD-ACE2 have been designed to prevent infection [65].
3.1.2. ACE2 Receptor for SARS-CoV-2
ACE2 is comprised of an extracellular-facing N-terminal domain and a C-terminal transmembrane domain with a cytosolic tail. The N-terminal portion of the protein contains the claw-like protease domain (PD), which can recognize the SARS-CoV-2 RBD via polar residues. The C-terminal domain is termed the Collectrin-like domain. The SARS-CoV-2 RBD combines with the ACE2 PD to form an RBD-PD complex (Figure 3). Therefore, as the receptor of SARS-CoV-2, ACE2 is a possible therapeutic target that needs to be studied in depth.
In the treatment of SARS-CoV-2, in addition to drugs and antibodies developed against the virus target, drugs with a high affinity to ACE2 have also been discovered. Dalbavancin, in the peptide drug library, can directly bind with ACE2 through the action of ACE2 amino acids. This binding prevents ACE2 from interacting with the viral S protein [66]. Although dalbavancin has not been extensively investigated in clinical trials, there is great hope for its future application. Telmisartan has been evaluated in clinical trials as an angiotensin receptor blocker [67]. Chlorogenic acid can stably bind to the Glu329/Gln325 and Gln42/Asp38 sites in ACE2 as a potential inhibitor of COVID-19 [68]. Researchers envision administering an antibody or single-chain antibody fragment that can bind the host cell membrane ACE2 protein and inhibit the interaction of the S protein with ACE2 [69]. OM-85 (a standardized lysate of human airway bacteria) and B2R (the kinin B2 receptor) are antagonists that can reduce the expression level of ACE2 in humans and prevent the transmission of signaling pathways [70]. Wei et al. reported that the exogenous supplementation of ACE2 via mesenchymal stem cells (MSCs) had better therapeutic effects, which might be due to the increased activities of the secretory ACE2 [71]. Despite the increasing attention paid to ACE2, there is scant literature on the therapeutic strategies and efficient drugs that target ACE2. Further information is critical to establish effective prevention and control actions.
3.2. Preventing SARS-CoV-2 Reproduction
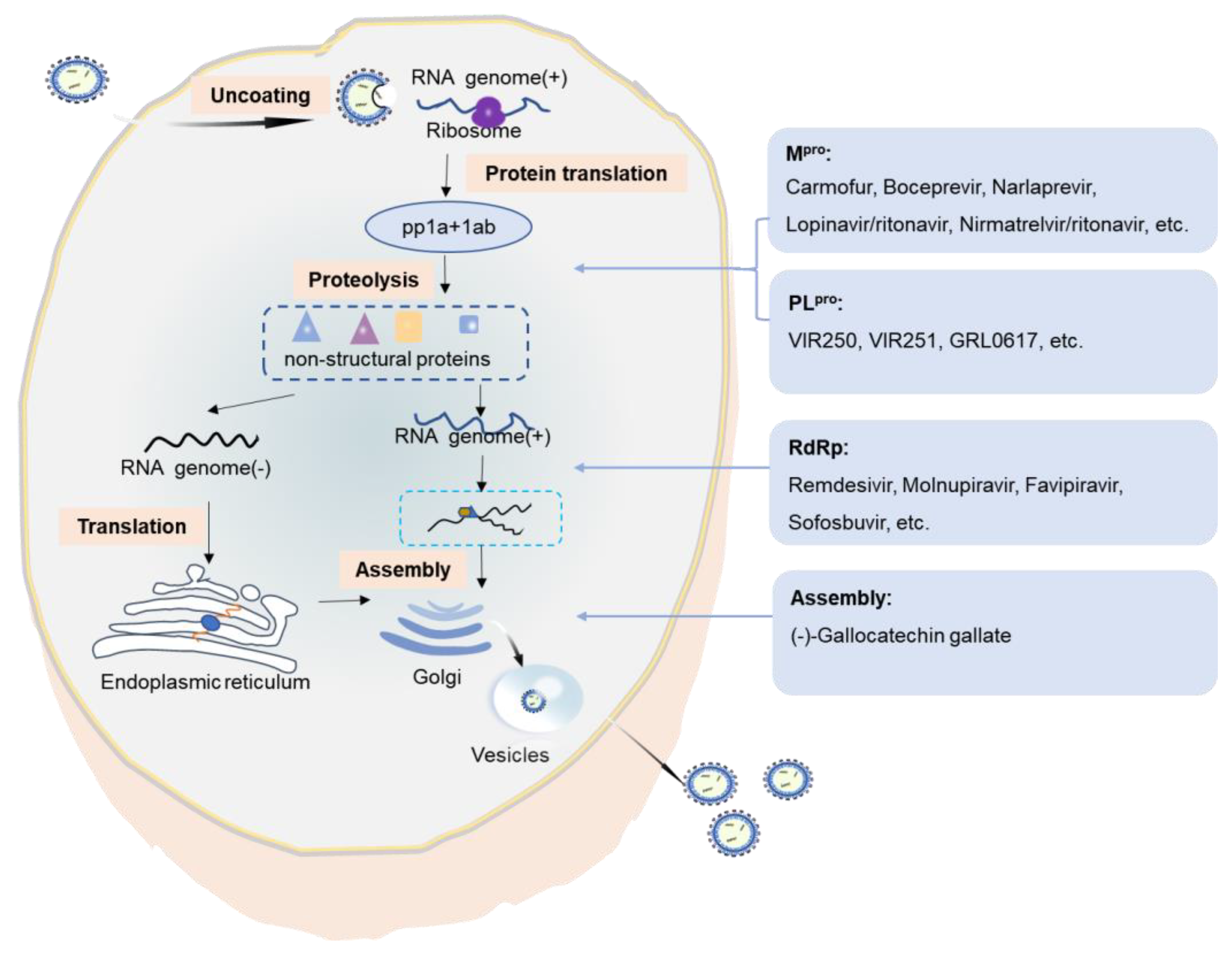
SARS-CoV-2 enters the host cell and replicates in the target cells. The SARS-CoV-2 genome is large and harbors two open reading frames (1a and 1b) at the 5′ end. These encode two large polyproteins, polyprotein 1a (pp1a) and polyprotein 1ab (pp1ab). The two polyproteins are cleaved and translated into a mature non-structural protein (NSP), which is responsible for the replication and transcription of the sub-genomic RNAs. The main protease, the 3C-like serine-type protease (3CLpro or Mpro) encoded by NSP5 and the papain-like proteinase (PLpro) encoded by NSP3, are important in the cleavage of polyproteins. PLpro cleaves the NSP1/2, NSP2/3, and NSP3/4 boundaries. The RNA-dependent RNA polymerase (RdRp; also known as NSP12) is a core component of the virus replicase–transcriptase complex responsible for the replication and transcription of viral RNA. Mpro, PLpro, and RdRp have indispensable roles in SARS-CoV-2 replication and are promising targets for drug design (Figure 4).
Mpro is essential for the cleavage of the viral polyprotein pp1ab at 11 discrete sites [72]. The released NSPs form a replicase–transcriptase complex, which in turn is responsible for viral replication [73]. The antineoplastic drug carmofur and anti-hepatitis C virus (HCV) drug boceprevir are approved drugs that are capable of inhibiting SARS-CoV-2 by targeting Mpro [74]. The carbonyl-reactive group of carmofur and trichostatin A can covalently bind to catalytic Cys145, while boceprevir occupies the substrate-binding pocket of Mpro and combines with the catalytic Cys145 to form a covalent bond [75,76]. Narlaprevir is a potent second-generation drug based on boceprevir. It inhibits the HCV NS3 protease and also has moderate inhibitory activity against SARS-CoV-2 Mpro by binding to the active site of Mpro through a C-S covalent bond with catalytic Cys145 [74]. 5-((1H-Imidazol-1-yl) methyl) quinolin-8-ol (DD1) is a derivative formed by the action of chloroquine and hydroxychloroquine. DD1 binds to the active residues in Mpro 6Y84 (GLN127, PHE223, and others) through hydrogen bond affinity. This can hinder the activity of the Mpro and inhibit the translation process [77,78]. Three modified dipeptides (11a, 11b, and 13b) were designed to inhibit the activity of Mpro. These compounds covalently block Cys145 by the formation of hemithioacetal/ketal [78,79]. The bioavailability of 11a was 87.8% in animal experiments [78]. Recently, two novel bicycloproline-containing Mpro inhibitors (MI-09 and MI-30) with excellent antiviral activity in cell-based assays were designed and synthesized. The inhibition modes of MI-09 and MI-30 are similar to that of 11a [80]. The only difference is that these compounds contain two cycloproline moieties, which obviously increase the exposure of the drugs in the body. Besides covalent inhibitors, baicalein is identified as a non-covalent inhibitor of SARS-CoV-2 Mpro. Baicalein occupies the core of the substrate-binding pocket, acting as a ‘shield’ in front of the catalytic dyad Cys145 to prevent the peptide substrate approaching the active site [81,82]. Through a combination of a structure-based drug design, Jin et al. analyzed over 10,000 compounds and 6 of those compounds (ebselen, shikonin, tideglusib, PX-12, disulfiram, and carmofur) were shown to inhibit Mpro [83]. Moreover, some drugs developed to treat HIV infections were studied as potential agents for COVID-19 [84]. Lopinavir/ritonavir (LPV/r), which is a prescription medicine approved by the FDA for the treatment of HIV infection, was proposed as a candidate agent due to its well binding to the SARS-CoV-2 Mpro [85]. In addition, nirmatrelvir/ritonavir (Paxlovid®) was approved on December 24, 2021, by the FDA for the emergency treatment of COVID-19 [86,87,88]. Nirmatrelvir inhibits the protein cleavage reaction of the Mpro of SARS-CoV-2, thereby inhibiting viral replication. Ritonavir acts as a booster by inhibiting a drug-metabolizing enzyme cytochrome p450 [89]. Paxlovid® would reduce the risk of hospitalization or death in 90% of subjects with mild-to-moderate disease [90].
SARS-CoV-2 PLpro is responsible for the cleavage of the N-termini of replicase polyproteins, which is critical in protein maturation. Compared with SARS-CoV-2 Mpro, there are fewer studies on SARS-CoV-2 PLpro inhibitors. Rut et al. used a combinatorial substrate library to perform a comprehensive activity profiling of SARS-CoV-2 PLpro. Two inhibitors (VIR250 and VIR251) designed to target SARS-CoV-2 occupy the S4-S1 pockets of the substrate-binding site, with high selectivity for SARS PLpro [91]. Naphthalene-based derivatives also inhibit SARS-CoV-2 replication by binding to the residues in the BL2 ring of PLpro [92]. A screen of a library of approved compounds revealed the activity of GRL0617 in preventing SARS-CoV-2 infection by binding to the catalytic domain and inhibiting the deacylation activity of PLpro [93]. Multiplexed enhanced protein dynamics (MePROD) revealed that the translation inhibitors cycloheximide and emetine can significantly inhibit the replication of SARS-CoV-2 [94]. MePROD can determine the changes in the proteome and translator in human cells. Cycloheximide is a translation extension inhibitor that can bind to the E site to block eEF2-mediated tRNA translocation. Emetine is a 40S ribosomal protein S14 inhibitor that inhibits the formation of peptide chains [95].
In addition, several drugs targeting virus RdRp have shown therapeutic potential against SARS-CoV-2. These drugs include favipiravir [96], sofosbuvir [97], ribavirin [98], galidesivir [99], remdesivir [100], molnupiravir [101], and so on. Among those, remdesivir, which was initially used to treat Ebola virus infections [100,102], is the first approval of the FDA for the emergency treatment of COVID-19 [103]. Remdesivir leads to the retention of the RNA 3ʹ-nucleotide at the substrate-binding site of the RNA polymerase, which interferes with the entry of the next nucleoside triphosphate and consequently inhibits RNA polymerization [104,105]. Up to now, although the drug has been approved for severe patients, the benefits of remdesivir for the treatment of COVID-19 remain debated as some clinical trials have shown discordant results [106,107,108]. The WHO recommends against the use of remdesivir for all patients, based on the results of the solidarity trial, which failed to observe the improvement in clinical mortality [109,110]. A meta-analysis concluded that there is a high probability that remdesivir reduces mortality for nonventilated patients with COVID-19 requiring supplemental oxygen therapy [107]. Similarly, molnupiravir, a prodrug for the ribonucleoside analogue β-d-N4-hydroxycytidine (NHC; EIDD-1931), has recently been approved in some countries for the patients with mild-to-moderate COVID-19 [18,111]. It was the first approval for oral drugs which have a major advantage over injectable drugs (remdesivir) against COVID-19 [112]. Suramin has stronger electronegativity than RNA polymerase and so can compete for binding to the gene sequence of the virus [113]. Suramin occupies two sites for RNA replication. One site prevents the primer from binding to the template strand [114]. The other site prevents the extension of the primer chain and hinders ATP from entering the catalytic site to provide energy [115]. Suramin is 20 times more powerful than remdesivir [114]. However, it is difficult to determine if the clinical benefits of suramin outweigh its toxic effects [116]. The suramin derivative NCTU-Alan-2026 is a small molecule that has less toxic effects. It is designed and synthesized to block protein interactions to inhibit cell proliferation [117]. The development of NCTU-Alan-2026 has provided new insights for the future development of antiviral drugs. Clofazimine also inhibits RNA replication by affecting the unwinding of RNA polymerase (NSP13) [118], which is essential for the CoV replication [119]. Interestingly, as an inhibitor of SARS-CoV-2 RdRp, Zn(II) may be a promising candidate, with direct inhibitory effects on the replicative cycle of SARS-CoV-2 [120]. In addition, some molecular docking studies showed that tenofovir and lamivudine, which are widely used to treat HIV infection, could be used effectively against SARS-CoV-2 [84].
In particular, during the replication process of SARS-CoV-2 in host cells, membrane and membrane-less organelles are formed to ensure the smooth progress of various biochemical processes [121]. Liquid–liquid phase separation (LLPS) is the basis for the formation of membrane-less organelles [122]. Protein and nucleic acid are condensed into a liquid-like condensate through LLPS and are assembled or disassembled in a spatially and temporally controlled manner [123]. Based on these findings, (−)-gallocatechin gallate, a green tea polyphenol, was recently discovered to efficiently prevent SARS-CoV-2 replication by blocking the LLPS of the N protein caused by RNA and affecting RNA and protein assembly [124].
3.3. Immunomodulatory Effects
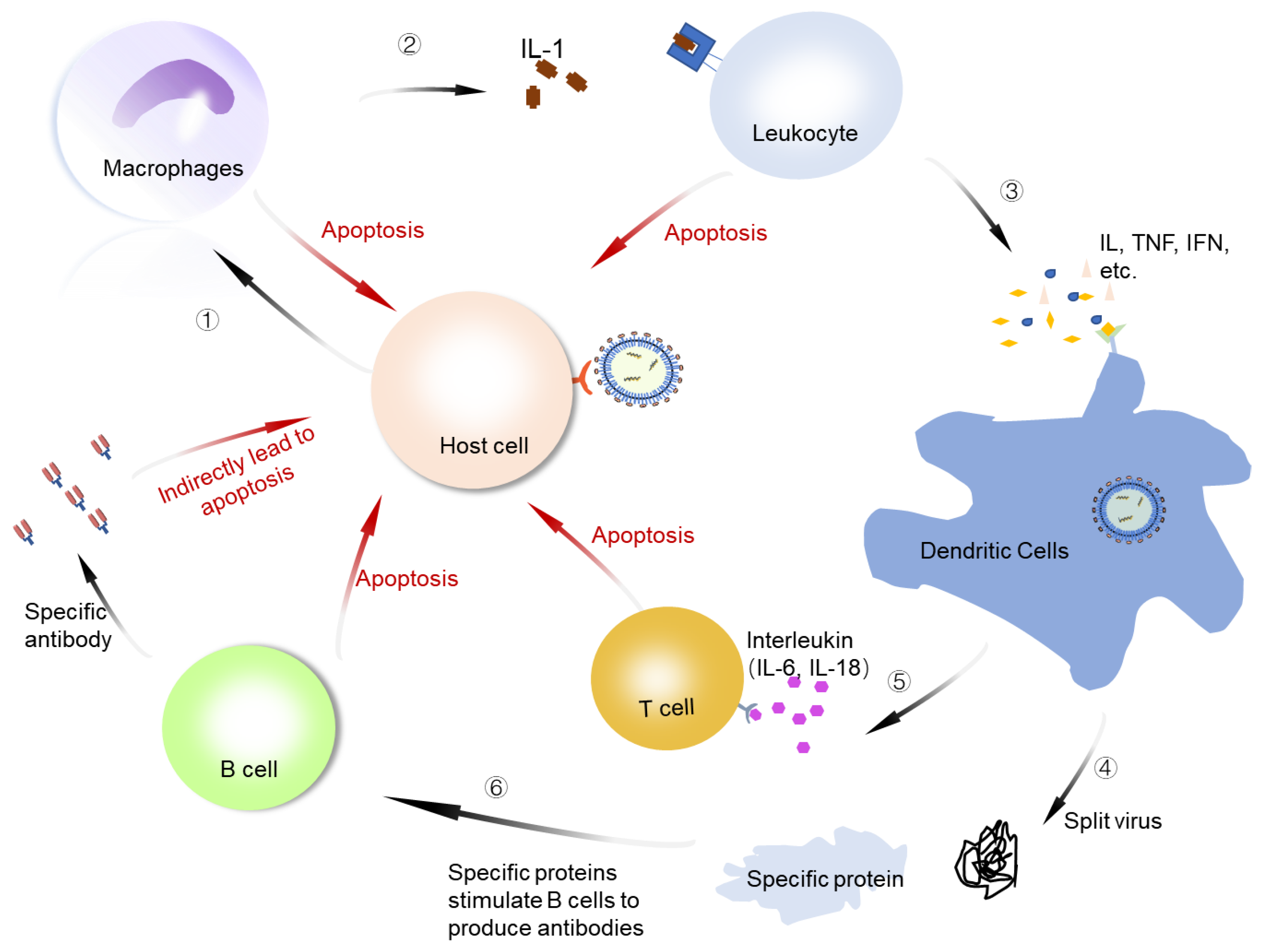
An important feature of COVID-19 is immune regulation disorder, which manifests as excessive inflammation. When SARS-CoV-2 infects the human body, it stimulates the body’s immune response line of defense. There are no serious clinical manifestations in the early stage of COVID-19 in patients who become critically ill or die. In the late stage, the viruses in the human body stimulate immune cells that include macrophages, dendritic cells, T cells, and white blood cells to activate anti-inflammatory factors that jointly respond to the invasion of viruses (Figure 5). The proinflammatory feed-back loop caused by SARS-CoV-2 infection could generate a massive production of chemokines, which orchestrate immune cell infiltration and the secretion of cytokines. The rapid rise of the concentrations of cytokines within a short period of time leads to a cytokine storm and acute respiratory distress syndrome. Some possible immune signatures in patients with COVID-19 and the targets for the treatment of the disease have been proposed. These include janus kinase (JAK), interleukins, tumor necrosis factor (TNF), and others [125]. The cytokine storm disrupts the immune function and is an important cause of severe illness and death. The production of SARS-CoV-2 symptoms is not clear, but it is closely related to specific cytokines [126,127]. The effective suppression of the cytokine storm should be an efficient way to prevent severe disease and death.
Clinical observations indicate that COVID-19 patients manifest an acute elevation of serum levels of inflammatory mediators, such as IL-6, IL-1, TNF-α, and JAK [128]. These elevations are associated with disease severity and progression. Therefore, the therapeutic strategies targeting IL-6, IL-1, TNF-α, and JAK have been proposed to quell the cytokine storm caused by SARS-CoV-2. A series of biologics have been designed that target IL-6, a key cytokine in the mediation of fever and the acute-phase response (Table 1). Among those, tocilizumab, which has been mainly used for the treatment of rheumatoid arthritis [129], is suggested to have beneficial effects in reducing the inflammatory response in severe COVID-19 patients. A meta-analysis of randomized clinical trials showed that tocilizumab was associated with a lower mortality rate in COVID-19 patients [130]. Moreover, the intravenous and subcutaneous forms of administration of tocilizumab almost have the same stability in treating patients with COVID-19 [131,132,133]. Sarilumab, a high-affinity anti-IL-6 receptor antibody, which has been approved for rheumatoid arthritis treatment by the FDA, has also showed efficiency in patients with COVID-19 [134]. Gremese et al. studied 53 patients who were treated with sarilumab, and the results showed that 89.7% of the patients significantly improved, 70.6% were discharged from the hospital, and 85.7% no longer needed oxygen therapy [135]. In addition, the same results were concluded by Benucci et al. [136]. Sarilumab is considered as an alternative regimen for the treatment of COVID-19 [137]. However, recently, some randomized trials showed that sarilumab treatment did not improve outcomes in patients with moderate-to-severe COVID-19 pneumonia [138,139]. So, further clinical studies with larger sample sizes and long-term follow-up are needed to assess the efficacy and safety of this drug. In addition, some traditional Chinese medicine, such as Glycyrrhizae Radix et Rhizoma (Gancao) and Pinelliae Rhizoma (Banxia), may act by suppressing the IL-6 amplifier and have been used to treat some moderately ill COVID-19 patients [140]. IL-1 is another important cytokine associated with inflammation. It is mainly produced by the activated mononuclear phagocytes. IL-1 can induce other proinflammatory cytokines, such as IL-6 and TNF-α. Several inhibitors that target IL-1, including canakinumab and anakinra, are effective for the treatment of the cytokine storm (Table 1) [141]. One report described patients from Saint Joseph’s hospital in Paris, France, who received anakinra treatment. The mortality rate and the need for invasive mechanical ventilation in the ICU were clearly reduced [142].
TNF-α is a key inflammatory factor that triggers a cytokine storm and is involved in mediating the inflammatory response in acute respiratory distress syndrome. Therefore, TNF-α inhibitors are expected to have promising therapeutic effects. A variety of TNF-α inhibitors, including adalimumab, etanercept, vitamin D, emapalumab, and infliximab, have been designed. Their effectiveness has not been proven in actual treatment (Table 1). JAK is an intracellular tyrosine kinase that mediates signals from cytokines, hormones, and growth factors. By blocking cytokine signaling and reducing excessive inflammatory responses, JAK inhibitors play an important role in the control of the cytokine storm in COVID-19 patients. Several JAK inhibitors, including baricitinib, ruxolitinib, tofacitinib, and fedratinib, are being studied for the treatment of severe COVID-19 (Table 1). Recently, two large clinical trials of baricitinib, involving 2558 patients hospitalized with COVID-19, showed significant clinical and survival benefits. Baricitinb has been recommended for clinical practice by multiple guidelines [143].
In addition to IL-6, IL-1, TNF-α, JAK, and other inflammatory mediators, such as the granulocyte-macrophage colony-stimulating factor (GM-CSF) and the complement protein C5, have been developed as therapeutic targets against the cytokine storm caused by SARS-CoV-2. GM-CSF is a cytokine with a cardinal role in the modulation of inflammation. Mavrilimumab is an anti-GM-CSF receptor-α monoclonal antibody that has improved clinical outcomes in patients with COVID-19. Earlier discharge from the hospital and no progression to death after mavrilimumab treatment were described [144]. Moreover, the complement system may be a valuable target for COVID-19 therapy, as it is an integral component of the innate immune response to virus infection. Eculizumab, a humanized mAb with a high affinity to complement factor C5, is developed for use in the treatment of COVID-19 [145,146].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Immunosuppressive agents are used to treat COVID-19 inflammatory phase.
| Target and Drugs | Initial Target | Administration Mode | Results | References |
|---|---|---|---|---|
| IL-6 | ||||
| Tocilizumab | Rheumatoid arthritis | Injection | Reduced mortality. | [147] |
| Babaodan | Infectious viral hepatitis | Oral | A large number of IL-6 and C-reactive proteins were eliminated in patients. | [148] |
| Methylprednisolone | Rheumatic disease | Injection | Patients with oxygen saturation improved rapidly and survival time became longer. | [149] |
| Olokizumab | Rheumatoid arthritis | Injection | The body temperature dropped to normal levels. | [150,151] |
| Sarilumab | Rheumatoid arthritis | Injection | A total of 89.7% of inpatients experienced significant improvement in symptoms and 70.6% were discharged. | [135] |
| Siltuximab | Castleman’s disease | Injection | Reduced mortality. | [152] |
| Clazakizumab | Rheumatoid arthritis | Injection | Symptoms, oxygen demand, radiologic findings, and inflammatory indicators improved markedly. | [153] |
| IL-1 | ||||
| Canakinumab | Rheumatologic disorders | Injection | Improved oxygenation, decreased serum C-reactive protein levels. | [154] |
| Anakinra | Rheumatoid arthritis | Injection | Relieved clinical signs in critically ill patients. | [155,156] |
| TNF-α | ||||
| Infiximab | Rheumatoid arthritis | Injection | Reduced serological response to SARS-CoV-2. | [157,158] |
| Etanercept | Psoriatic arthritis | Oral | Delayed the development of olfactory and taste dysfunction in patients. | [159,160] |
| Adalimumab | Rheumatoid arthritis | Injection | Radiological improvement of the lung without any complications. | [161,162] |
| Vitamin D | Anti-rickets | - | Inhibited the expression of proinflammatory cytokines and reduced viral load. | [163] |
| Emapalumab | Hemophagocytic | - | Effective in a refractory, persistent, and progressive cytokine storm. | [164] |
| JAK | ||||
| Ruxolitinib | Myelofibrosis | Oral | The cytokine content was reduced, and the patient’s chest CT improved by 80%. | [165] |
| Fedratinib | Myeloproliferative tumor | - | Reduced patient mortality. | [166] |
| Tofacitinib | Rheumatoid arthritis | Injection | Respiratory symptoms were relieved after 5 days. | [167] |
| Baricitinib | Rheumatoid arthritis | Oral | Accelerate improvement in clinical status and reduce 28-day mortality. | [168,169,170] |
| IL (IL-1, IL-3, IL-2, IL-5, IL-6, IL-8) and TNF-α | ||||
| Glucocorticoid | SARS; MERS | Injection | Significantly reduced risk of death by 62%. | [171,172] |
| Lidocaine/dexamethasone | - | - | Showed anti-inflammatory capacity on SARS-CoV-2-triggered immune pathways. | [173] |
| Fluvoxamine | Antidepressant | Injection | Reduced the need for advanced disease care in this high-risk population. | [174] |
| IL-6, IL-8, TNF-α | ||||
| Azithromycin | Macrolide antibiotics | Oral | Low mortality. | [175,176] |
| Melatonin | Immune system | - | Effectively improved inflammation. | [177] |
| Cyclosporine A | Hepatitis B virus | - | Reduced mortality in patients with moderate-to-severe disease. | [178] |
| GM-CSF | ||||
| Mavrilimumab | Refractory rheumatoid arthritis | Injection | Improved clinical outcomes, well tolerated without discomfort. | [144] |
| C5 | ||||
| Eculizumab | Orphan disease | Injection | Reduced inflammatory responses and improvedlung function and lymphocyte recovery. | [145,146] |
Abbreviations: TNF-α, tumor necrosis factor-α; IL, interleukin; JAK, Janus kinase; GM-CSF, granulocyte–macrophage colony-stimulating factor; CCR5, C–C chemokine receptor type 5.
4. Conclusions and Perspectives
The development of effective treatments for SARS-CoV-2 remains an urgent priority. This review summarizes possible treatment strategies that target the entry of the virus, virus replication, and the immune system. The aim is to inform the development of efficacious prevention and treatment strategies. The consideration of targeted treatment to prevent SARS-CoV-2 entry into human cells and the development of symptoms will hopefully bring novel insight into the treatment of COVID-19.
Some antiviral drugs have specific effects against SARS-CoV-2. The efficacy and safety of these promising candidate drugs in the treatment of SARS-CoV-2 need to be confirmed. Moreover, some monoclonal antibodies have demonstrated efficacy against COVID-19 and have been approved in some countries for the treatment and prevention of COVID-19, but the emergence of variant lineages is now one of the most difficult obstacles to controlling the COVID-19 pandemic.
The infection by SARS-CoV-2 may cause disseminated intravascular coagulation and venous thromboembolism in patients with some critically ill disease. Some research has demonstrated that anticoagulant therapy, mainly with low-molecular-weight heparin, has been associated with better outcomes in severe COVID-19, especially in the early treatment [179,180]. However, the use of anticoagulation or high doses of heparin is still controversial.
In addition, several studies showed the use of immune-suppressive drugs combined with the natural immune suppression caused by the virus can lead to increasing secondary bacterial and fungal infections [181,182,183], but more research about antimicrobial resistance and its correlation with antibiotic misuse in COVID-19 patients is required [184]. Moreover, the high rate of multidrug-resistant organisms among hospitalized patients alerted that it is urgent to use the antibiotics with a higher potential for resistance and to evaluate trends in antibiotic resistance [185].
With the rapid spread of SARS-CoV-2, as well as the rising global mortality rate of COVID-19, research has focused on the development of potential vaccine candidates for protection against SARS-CoV-2. To date, the total number of vaccine doses administered globally has reached 90.9 billion. Along with the virus spread, several mutant strains have emerged. These mutant strains may result in epitope changes that reduce the affinity of antibodies produced by neutralizing antibody therapy or vaccines. Therefore, developing effective, specific, and general antiviral drugs is crucial to end the COVID-19 pandemic and to prevent future social and economic hardships.
Author Contributions
Conceptualization, J.M. and Z.H.; writing—original draft preparation, J.M. and R.L.; writing—review and editing, J.M. and Z.H.; visualization, J.M., Z.Z. (Zhiqi Zhang), J.W., Q.H., D.N., K.F., W.G. and Z.Z. (Zhihui Zhao); project administration, Z.H.; funding acquisition, Z.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Shanghai Agriculture Applied Technology Development Program, China, grant number X20210302, and the National Natural Science Foundation of China, grant number 31972178.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, genetic recombination, and pathogenesis of coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forster, P.; Forster, L.; Renfrew, C.; Forster, M. Phylogenetic network analysis of SARS-CoV-2 genomes. Proc. Natl. Acad. Sci. USA 2020, 117, 9241–9243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vito, A.; Fiore, V.; Princic, E.; Geremia, N.; Panu Napodano, C.M.; Muredda, A.A.; Maida, I.; Madeddu, G.; Babudieri, S. Predictors of infection, symptoms development, and mortality in people with SARS-CoV-2 living in retirement nursing homes. PLoS ONE 2021, 16, e0248009. [Google Scholar] [CrossRef] [PubMed]
- Geremia, N.; De Vito, A.; Gunnella, S.; Fiore, V.; Princic, E.; Napodano, C.P.; Madeddu, G.; Babudieri, S. A case of vasculitis-like skin eruption associated with COVID-19. Infect. Dis. Clin. Prac. 2020, 28, E30–E31. [Google Scholar] [CrossRef]
- Vaira, L.A.; Hopkins, C.; Salzano, G.; Petrocelli, M.; Melis, A.; Cucurullo, M.; Ferrari, M.; Gagliardini, L.; Pipolo, C.; Deiana, G.; et al. Olfactory and gustatory function impairment in COVID-19 patients: Italian objective multicenter-study. Head Neck 2020, 42, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Hope, A.A.; Evering, T.H. Postacute sequelae of severe acute respiratory syndrome coronavirus 2 infection. Infect. Dis. Clin. N. Am. 2022, 36, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.J.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Cosentino, F.; Moscatt, V.; Marino, A.; Pampaloni, A.; Scuderi, D.; Ceccarelli, M.; Benanti, F.; Gussio, M.; Larocca, L.; Boscia, V.; et al. Clinical characteristics and predictors of death among hospitalized patients infected with SARS-CoV-2 in Sicily, Italy: A retrospective observational study. Biomed. Rep. 2022, 16, 34. [Google Scholar] [CrossRef]
- Rezaee, H.; Pourkarim, F.; Pourtaghi-Anvarian, S.; Entezari-Maleki, T.; Asvadi-Kermani, T.; Nouri-Vaskeh, M. Drug-drug interactions with candidate medications used for COVID-19 treatment: An overview. Pharmacol. Res. Perspect. 2021, 9, e00705. [Google Scholar] [CrossRef]
- Liu, L.; Wei, Q.; Alvarez, X.; Wang, H.; Du, Y.; Zhu, H.; Jiang, H.; Zhou, J.; Lam, P.; Zhang, L.; et al. Epithelial cells lining salivary gland ducts are early target cells of severe acute respiratory syndrome coronavirus infection in the upper respiratory tracts of rhesus macaques. J. Virol. 2011, 85, 4025–4030. [Google Scholar] [CrossRef]
- Guo, Y.; Korteweg, C.; McNutt, M.A.; Gu, J. Pathogenetic mechanisms of severe acute respiratory syndrome. Virus Res. 2008, 133, 4–12. [Google Scholar] [CrossRef] [PubMed]
- De Vito, A.; Geremia, N.; Princic, E.; Fanelli, C.; Panu Napodano, C.M.; Muredda, A.A.; Fiore, V.; Maida, I.; Fois, A.G.; Babudieri, S.; et al. Does angiotensin II receptor blockers increase the risk of SARS-CoV-2 infection? a real-life experience. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 523–526. [Google Scholar] [PubMed]
- Fosbol, E.L.; Butt, J.H.; Ostergaard, L.; Andersson, C.; Selmer, C.; Kragholm, K.; Schou, M.; Phelps, M.; Gislason, G.H.; Gerds, T.A.; et al. Association of angiotensin-converting enzyme inhibitor or angiotensin receptor blocker use with COVID-19 diagnosis and mortality. JAMA 2020, 324, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, R.T.; Lynch, J.B.; del Rio, C. Mild or moderate covid-19. N. Engl. J. Med. 2020, 383, 1757–1766. [Google Scholar] [CrossRef]
- COVID-19 Dashboard by the Center for Systems Science and Engineering at Johns Hopkins University. Available online: https://coronavirus.jhu.edu/map.html (accessed on 15 October 2022).
- Chen, Y.; Liu, Q.Y.; Guo, D.Y. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef] [Green Version]
- Kirtipal, N.; Bharadwaj, S.; Kang, S.G. From SARS to SARS-CoV-2, insights on structure, pathogenicity and immunity aspects of pandemic human coronaviruses. Infect. Genet. Evol. 2020, 85, 104502. [Google Scholar] [CrossRef]
- De Vito, A.; Colpani, A.; Bitti, A.; Zauli, B.; Meloni, M.C.; Fois, M.; Denti, L.; Bacciu, S.; Marcia, C.; Maida, I.; et al. Safety and efficacy of molnupiravir in SARS-CoV-2-infected patients: A real-life experience. J. Med. Virol. 2022, 94, 5582–5588. [Google Scholar] [CrossRef]
- Garcia-Lledo, A.; Gomez-Pavpn, J.; Gonzalez Del Castillo, J.; Hernandez-Sampelayo, T.; Martin-Delgado, M.C.; Martin Sanchez, F.J.; Martinez-Selles, M.; Molero Garcia, J.M.; Moreno Guillen, S.; Rodriguez-Artalejo, F.J.; et al. Pharmacological treatment of COVID-19: An opinion paper. Rev. Esp. Quimioter. 2022, 35, 115–130. [Google Scholar] [CrossRef]
- Atluri, K.; Aimlin, I.; Arora, S. Current effective therapeutics in management of COVID-19. J. Clin. Med. 2022, 11, 3838. [Google Scholar] [CrossRef]
- Hui, D.S.; Azhar, E.I.; Madani, T.A.; Ntoumi, F.; Kock, R.; Dar, O.; Ippolito, G.; Mchugh, T.D.; Memish, Z.A.; Drosten, C.; et al. The continuing 2019-nCoV epidemic threat of novel coronaviruses to global health-the latest 2019 novel coronavirus outbreak in Wuhan, China. Int. J. Infect. Dis. 2020, 91, 264–266. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The species severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar]
- South, A.M.; Diz, D.I.; Chappell, M.C. COVID-19, ACE2, and the cardiovascular consequences. Am. J. Physiol.-Heart C. 2020, 318, H1084–H1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef] [PubMed]
- Heurich, A.; Hofmann-Winkler, H.; Gierer, S.; Liepold, T.; Jahn, O.; Pohlmann, S. TMPRSS2 and ADAM17 cleave ACE2 differentially and only proteolysis by TMPRSS2 augments entry driven by the severe acute respiratory syndrome coronavirus spike protein. J. Virol. 2014, 88, 1293–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, C.G.K.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M.; et al. SARS-CoV-2 receptor ACE2 is an interferon-stimulated gene in human airway epithelial cells and is detected in specific cell subsets across tissues. Cell 2020, 181, 1016–1035. [Google Scholar] [CrossRef] [PubMed]
- Rendeiro, A.F.; Ravichandran, H.; Bram, Y.; Chandar, V.; Kim, J.; Meydan, C.; Park, J.; Foox, J.; Hether, T.; Warren, S.; et al. The spatial landscape of lung pathology during COVID-19 progression. Nature 2021, 593, 564–569. [Google Scholar] [CrossRef]
- Bamidis, A.D.; Koehler, P.; di Cristanziano, V.; Rasche, K.; Demirel, B.; Bacher, P.; Hallek, M.; Kochanek, M.; Klen, F.; Hofmann, S.C.; et al. First manifestation of adult-onset still’s disease after COVID-19. Lancet Rheumatol. 2021, 3, E319–E321. [Google Scholar] [CrossRef]
- Lechien, J.R.; Chiesa-Estomba, C.M.; De Siati, D.R.; Horoi, M.; Le Bon, S.D.; Rodriguez, A.; Dequanter, D.; Blecic, S.; Afia, F.E.; Distinguin, L.; et al. Olfactory and gustatory dysfunctions as a clinical presentation of mild-to-moderate forms of the coronavirus disease (COVID-19): A multicenter European study. Eur. Arch. Otorhinolaryngol. 2020, 277, 2251–2261. [Google Scholar] [CrossRef]
- Taquet, M.; Geddes, J.R.; Husain, M.; Luciano, S.; Harrison, P.J. 6-month neurological and psychiatric outcomes in 236379 survivors of COVID-19: A retrospective cohort study using electronic health records. Lancet Psychiat. 2021, 8, 416–427. [Google Scholar] [CrossRef]
- Weng, J.; Li, Y.; Li, J.; Shen, L.H.; Zhu, L.; Liang, Y.; Lin, X.; Jiao, N.; Cheng, S.; Huang, Y.; et al. Gastrointestinal sequelae 90 days after discharge for COVID-19. Lancet Gastroenterol. Hepatol. 2021, 6, 344–346. [Google Scholar] [CrossRef]
- Goldberg, M.F.; Goldberg, M.F.; Cerejo, R.; Tayal, A.H. Cerebrovascular disease in COVID-19. Am. J. Neuroradiol. 2020, 41, 1170–1172. [Google Scholar] [CrossRef]
- Poyiadji, N.; Shahin, G.; Noujaim, D.; Stone, M.; Patel, S.; Griffith, B. COVID-19-associated acute hemorrhagic necrotizing encephalopathy: Imaging features. Radiology 2020, 296, E119–E120. [Google Scholar] [CrossRef] [Green Version]
- Finsterer, J.; Scorza, F.A. Seizure and neuropsychiatric compromise as onset of SARS-CoV-2 associated encephalitis. Psychiatry Res. Commun. 2022, 2, 100029. [Google Scholar] [CrossRef]
- Radhakrishnan, R.K.; Kandasamy, M. SARS-CoV-2-mediated neuropathogenesis, deterioration of hippocampal neurogenesis and dementia. Am. J. Alzheimer’s Dis. 2022, 37, 15333175221078418. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.Z.; Dutta, D.; Ravichandiran, V.; Sukla, S. Monoclonal antibodies: A remedial approach to prevent SARS-CoV-2 infection. 3 Biotech 2022, 12, 227. [Google Scholar] [CrossRef]
- Mungroo, M.R.; Khan, N.A.; Siddiqui, R. The increasing importance of the novel coronavirus. Hosp. Pract. 2021, 49, 1–11. [Google Scholar] [CrossRef]
- Xiong, X.; Qu, K.; Ciazynska, K.A.; Hosmillo, M.; Carter, A.P.; Ebrahimi, S.; Ke, Z.; Scheres, S.H.W.; Bergamaschi, L.; Grice, G.L.; et al. A thermostable, closed SARS-CoV-2 spike protein trimer. Nat. Struct. Mol. Biol. 2020, 27, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Zhao, Y.; Sun, Y.; Zhang, B.; Wang, H.; Wu, Y.; Zhu, Y.; Zhu, C.; Hu, T.; Du, X.; et al. Structural basis for the inhibition of SARS-CoV-2 main protease by antineoplastic drug carmofur. Nat. Struct. Mol. Biol. 2020, 27, 529–532. [Google Scholar] [CrossRef]
- Ke, Z.; Oton, J.; Qu, K.; Cortese, M.; Zila, V.; McKeane, L.; Nakane, T.; Zivanov, J.; Neufeldt, C.J.; Cerikan, B.; et al. Structures and distributions of SARS-CoV-2 spike proteins on intact virions. Nature 2020, 588, 498–502. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef]
- Parveen, S.; Alnoman, R.B. Potential exploration of recent FDA-approved anticancer drugs against models of SARS-CoV-2’s main protease and spike glycoprotein: A computational study. Biointerface Res. App. 2021, 11, 10059–10073. [Google Scholar]
- Yuan, M.; Liu, H.J.; Wu, N.I.C.; Wilson, I.A. Recognition of the SARS-CoV-2 receptor binding domain by neutralizing antibodies. Biochem. Biophys. Res. Commun. 2021, 538, 192–203. [Google Scholar] [CrossRef]
- Zhao, G.; He, L.; Sun, S.; Qiu, H.; Tai, W.; Chen, J.; Li, J.; Chen, Y.; Guo, Y.; Wang, Y.; et al. A novel nanobody targeting middle east respiratory syndrome coronavirus (MERS-CoV) receptor-binding domain has potent cross-neutralizing activity and protective efficacy against MERS-CoV. J. Virol. 2018, 92, e00837-18. [Google Scholar] [CrossRef] [Green Version]
- Zost, S.J.; Gilchuk, P.; Chen, R.E.; Case, J.B.; Reidy, J.X.; Trivette, A.; Nargi, R.S.; Sutton, R.E.; Suryadevara, N.; Chen, E.C.; et al. Rapid isolation and profiling of a diverse panel of human monoclonal antibodies targeting the SARS-CoV-2 spike protein. Nat. Med. 2020, 26, 1422–1427. [Google Scholar] [CrossRef]
- Wrapp, D.; De Vlieger, D.; Corbett, K.S.; Torres, G.M.; Wang, N.; Van Breedam, W.; Roose, K.; van Schie, L.; VIB-CMB COVID-19 Response Team; Hoffmann, M.; et al. Structural basis for potent neutralization of betacoronaviruses by single-domain camelid antibodies. Cell 2020, 181, 1004–1015. [Google Scholar] [CrossRef]
- Gupta, A.; Gonzalez-Rojas, Y.; Juarez, E.; Crespo Casal, M.; Moya, J.; Falci, D.R.; Sarkis, E.; Solis, J.; Zheng, H.; Scott, N.; et al. Early treatment for Covid-19 with SARS-CoV-2 neutralizing antibody sotrovimab. N. Engl. J. Med. 2021, 385, 1941–1950. [Google Scholar] [CrossRef]
- Heo, Y.A. Sotrovimab: First approval. Drugs 2022, 82, 77–484. [Google Scholar] [CrossRef]
- Hwang, Y.C.; Lu, R.M.; Su, S.C.; Chiang, P.Y.; Ko, S.H.; Ke, F.Y.; Liang, K.H.; Hsieh, T.Y.; Wu, H.C. Monoclonal antibodies for COVID-19 therapy and SARS-CoV-2 detection. J. Biomed. Sci. 2022, 29, 1. [Google Scholar] [CrossRef]
- Westendorf, K.; Zentelis, S.; Wang, L.; Foster, D.; Vaillancourt, P.; Wiggin, M.; Lovett, E.; van der Lee, R.; Hendle, J.; Pustilnik, A. LY-CoV1404 (bebtelovimab) potently neutralizes SARS-CoV-2 variants. Cell Rep. 2022, 39, 110812. [Google Scholar] [CrossRef]
- Iketani, S.; Liu, L.; Guo, Y.; Liu, L.; Chan, J.F.; Huang, Y.; Wang, M.; Luo, Y.; Yu, J.; Chu, H.; et al. Antibody evasion properties of SARS-CoV-2 omicron sublineages. Nature 2022, 604, 553–556. [Google Scholar] [CrossRef]
- Deeks, E.D. Casirivimab/Imdevimab: First approval. Drugs. 2021, 81, 2047–2055. [Google Scholar] [CrossRef]
- Sidebottom, D.B.; Gill, D. Ronapreve for prophylaxis and treatment of covid-19. BMJ 2021, 374, n2136. [Google Scholar] [CrossRef]
- Loo, Y.M.; McTamney, P.M.; Arends, R.H.; Abram, M.E.; Aksyuk, A.A.; Diallo, S.; Flores, D.J.; Kelly, E.J.; Ren, K.; Roque, R.; et al. The SARS-CoV-2 monoclonal antibody combination, AZD7442, is protective in nonhuman primates and has an extended half-life in humans. Sci. Transl. Med. 2022, 14, eabl8124. [Google Scholar] [CrossRef]
- Levin, M.J.; Ustianowski, A.; De Wit, S.; Launay, O.; Avila, M.; Templeton, A.; Yuan, Y.; Seegobin, S.; Ellery, A.; Levinson, D.J.; et al. Intramuscular AZD7442 (Tixagevimab-Cilgavimab) for prevention of Covid-19. N. Engl. J. Med. 2022, 386, 2188–2200. [Google Scholar] [CrossRef]
- Kaplon, H.; Chenoweth, A.; Crescioli, S.; Reichert, J.M. Antibodies to watch in 2022. MAbs 2022, 14, 2014296. [Google Scholar] [CrossRef]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hamers, C.; Songa, E.B.; Bendahhman, N.; Hamers, R. Naturally occurring antibodies devoid of light chains. Nature 1993, 21, 446–448. [Google Scholar] [CrossRef]
- Xu, Y.; Xiong, L.; Li, Y.; Xiong, Y.; Tu, Z.; Fu, J.; Chen, B. Anti-idiotypic nanobody as citrinin mimotope from a naive alpaca heavy chain single domain antibody library. Anal. Bioanal. Chem. 2015, 407, 5333–5341. [Google Scholar] [CrossRef]
- Huo, J.; Le Bas, A.; Ruza, R.R.; Duyvesteyn, H.M.E.; Mikolajek, H.; Malinauska, T.; Tan, T.K.; Rijal, P.; Dumoux, M.; Ward, P.N.; et al. Neutralizing nanobodies bind SARS-CoV-2 spike RBD and block interaction with ACE2. Nat. Struct. Mol. Biol. 2020, 27, 846–854. [Google Scholar] [CrossRef]
- Nikiforuk, A.M.; Kuchinski, K.S.; Twa, D.D.W.; Lukac, C.D.; Sbihi, H.; Basham, C.A.; Steidl, C.; Prystajecky, N.A.; Jassem, A.N.; Krajden, M.; et al. The contrasting role of nasopharyngeal angiotensin converting enzyme 2 (ACE2) transcription in SARS-CoV-2 infection: A cross-sectional study of people tested for COVID-19 in British Columbia. EBioMedicine 2021, 66, 103316. [Google Scholar] [CrossRef]
- Wu, P.; Yang, Q.; Zhao, X.; Liu, Q.; Xi, J.; Zhang, F.; He, J.; Yang, H.; Zhang, C.; Ma, Z.; et al. A SARS-CoV-2 nanobody that can bind to the RBD region may be used for treatment in COVID-19 in animals. Res. Vet. Sci. 2022, 145, 46–49. [Google Scholar] [CrossRef]
- Sun, M.; Liu, S.; Wei, X.; Wan, S.; Huang, M.; Song, T.; Lu, Y.; Weng, X.; Lin, Z.; Chen, H.; et al. Aptamer blocking strategy inhibits SARS-CoV-2 virus infection. Angew. Chem. Int. Edit. 2021, 60, 10266–10272. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Wu, J.; Qi, J.; Zeng, Z.; Wan, Q.; Chen, Z.; Manandhar, P.; Cavener, V.S.; Fu, X.; et al. Neutralizing aptamers block S/RBD-ACE2 interactions and prevent host cell infection. Angew. Chem. Int. Ed. Engl. 2021, 60, 10273–10278. [Google Scholar] [CrossRef]
- Wang, G.; Yang, M.L.; Duan, Z.L.; Liu, F.L.; Jin, L.; Long, C.B.; Zhang, M.; Tang, X.P.; Xu, L.; Li, Y.C.; et al. Dalbavancin binds ACE2 to block its interaction with SARS-CoV-2 spike protein and is effective in inhibiting SARS-CoV-2 infection in animal models. Cell Res. 2021, 31, 17–24. [Google Scholar] [CrossRef]
- Rothlin, R.P.; Vetulli, H.M.; Duarte, M.; Pelorosso, F.G. Telmisartan as tentative angiotensin receptor blocker therapeutic for COVID-19. Drug Dev. Res. 2020, 81, 768–770. [Google Scholar] [CrossRef]
- Yu, J.; Wang, L.; Bao, L. Exploring the active compounds of traditional mongolian medicine in intervention of novel coronavirus (COVID-19) based on molecular docking method. J. Funct. Foods 2020, 71, 104016. [Google Scholar] [CrossRef]
- Kruse, R.L. Therapeutic strategies in an outbreak scenario to treat the novel coronavirus originating in Wuhan, China. F1000Research 2020, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Jakwerth, C.A.; Feuerherd, M.; Guerth, F.M.; Oelsner, M.; Schellhammer, L.; Giglberger, J.; Pechtold, L.; Jerin, C.; Kugler, L.; Mogler, C.; et al. Early reduction of SARS-CoV-2-replication in bronchial epithelium by kinin B2 receptor antagonism. J. Mol. Med. 2022, 100, 613–627. [Google Scholar] [CrossRef]
- Wei, J.; Shang, R.; Wang, J.; Zhu, S.; Yin, J.Q.; Chen, Y.; Zhao, Y.; Chen, G. ACE2 overexpressing mesenchymal stem cells alleviates COVID-19 lung injury by inhibiting pyroptosis. iScience 2022, 25, 104046. [Google Scholar] [CrossRef]
- Anand, K.; Palm, G.J.; Mesters, J.R.; Siddell, S.G.; Ziebuhr, J.; Hilgenfeld, R. Structure of coronavirus main proteinase reveals combination of a chymotrypsin fold with an extra α-helical domain. EMBO J. 2002, 21, 3213–3224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.; Zhou, F.; Huang, Z.; Ma, X.; Natarajan, K.; Zhang, M.; Huang, Y.; Su, H. Molecular insights into small-molecule drug discovery for SARS-CoV-2. Angew. Chem. Int. Ed. 2021, 60, 9789–9802. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Ye, F.; Feng, Y.; Liao, H.; Song, H.; Qi, J.; Gao, G.F.; Tan, W.; Fu, L.; Shi, Y. Structural basis for the inhibition of the SARS-CoV-2 main protease by the anti-HCV drug narlaprevir. Signal Transduct. Target. Ther. 2021, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Ye, F.; Feng, Y.; Yu, F.; Wang, Q.; Wu, Y.; Zhao, C.; Sun, H.; Huang, B.; Niu, P.; et al. Both boceprevir and GC376 efficaciously inhibit SARS-CoV-2 by targeting its main protease. Nat. Commun. 2020, 11, 4417. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Tang, K.M.; Chik, K.K.H.; Chan, C.C.Y.; Tsang, J.O.L.; Liang, R.; Gao, J.; Huang, Y.; Luo, C.; Cai, J.P.; et al. In silico structure-based discovery of a SARS-CoV-2 main protease inhibitor. Int. J. Biol. Sci. 2021, 17, 1555–1564. [Google Scholar] [CrossRef]
- Douche, D.; Sert, Y.; Brandan, S.A.; Kawther, A.A.; Bilmez, B.; Dege, N.; Louzi, A.E.; Bougrin, K.; Karrouchi, K.; Himmi, B. 5-((1H-imidazol-1-yl) methyl) quinolin-8-ol as potential antiviral SARS-CoV-2 candidate: Synthesis, crystal structure, hirshfeld surface analysis, DFT and molecular docking studies. J. Mol. Struct. 2021, 1232, 130005. [Google Scholar] [CrossRef]
- Dai, W.; Zhang, B.; Jiang, X.M.; Su, H.; Li, J.; Zhao, Y.; Xie, X.; Jin, Z.; Peng, J.; Liu, F.; et al. Structure-based design of antiviral drug candidates targeting the SARS-CoV-2 main protease. Science 2020, 368, 1331–1335. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Qiao, J.; Li, Y.S.; Zeng, R.; Liu, F.L.; Luo, R.H.; Huang, C.; Wang, Y.F.; Zhang, J.; Quan, B.; Shen, C.; et al. SARS-CoV-2 Mpro inhibitors with antiviral activity in a transgenic mouse model. Science 2021, 371, 1374–1378. [Google Scholar] [CrossRef]
- Liu, H.; Ye, F.; Sun, Q.; Liang, H.; Li, C.; Li, S.; Lu, R.; Huang, B.; Tan, W.; Lai, L. Scutellaria baicalensis extract and baicalein inhibit replication of SARS-CoV-2 and its 3C-like protease in vitro. J. Enzym. Inhib. Med. Chem. 2021, 36, 497–503. [Google Scholar] [CrossRef]
- Su, H.; Yao, S.; Zhao, W.; Li, M.; Liu, J.; Shang, W.; Xie, H.; Ke, C.; Hu, H.; Gao, M.; et al. Anti-SARS-Cov-2 activities in vitro of Shuanghuanglian preparations and bioactive ingredients. Acta Pharmacol. Sin. 2020, 41, 1167–1677. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Dallocchio, R.N.; Dessi, A.; De Vito, A.; Delogu, G.; Serra, P.A.; Madeddu, G. Early combination treatment with existing HIV antivirals: An effective treatment for COVID-19? Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 2435–2448. [Google Scholar] [PubMed]
- Elmekaty, E.Z.I.; Alibrahim, R.; Hassanin, R.; Eltaib, S.; Elsayed, A.; Rustom, F.; Ibrahim, M.I.M.; Abu Khattab, M.; Al Soub, H.; Al Maslamani, M.; et al. Darunavir-cobicistat versus lopinavir-ritonavir in the treatment of COVID-19 infection (DOLCI): A multicenter observational study. PLoS ONE 2022, 17, e0267884. [Google Scholar] [CrossRef]
- Hung, Y.P.; Lee, J.C.; Chiu, C.W.; Lee, C.C.; Tsai, P.J.; Hsu, I.L.; Ko, W.C. Oral nirmatrelvir/ritonavir therapy for COVID-19: The dawn in the dark? Antibiotics 2022, 11, 220. [Google Scholar] [CrossRef]
- Hammond, J.; Leister-Tebbe, H.; Gardner, A.; Abreu, P.; Bao, W.; Wisemandle, W.; Baniecki, M.; Hendrick, V.M.; Damle, B.; Simon-Campos, A. Oral nirmatrelvir for high-risk, nonhospitalized adults with Covid-19. N. Engl. J. Med. 2022, 386, 1397–1408. [Google Scholar] [CrossRef]
- Wong, C.K.H.; Au, I.C.H.; Lau, K.T.K.; Lau, E.H.Y.; Cowling, B.J.; Leung, G.M. Real-world effectiveness of early molnupiravir or nirmatrelvir-ritonavir in hospitalised patients with COVID-19 without supplemental oxygen requirement on admission during Hong Kong’s omicron BA.2 wave: A retrospective cohort study. Lancet Infect. Dis. 2022; in press. [Google Scholar] [CrossRef]
- NIH COVID-19 Treatment Guidelines. Ritonavir-Boosted Nirmatrelvir (Paxlovid). Available online: https://www.covid19treatmentguidelines.nih.gov/therapies/antiviral-therapy/ritonavir-boosted-nirmatrelvir--paxlovid-/ (accessed on 21 March 2022).
- Mayer, C.; VanHise, K.; Caskey, R.; Naqvi, M.; Burwick, R.M. Monoclonal antibodies casirivimab and imdevimab in pregnancy for coronavirus disease 2019 (COVID-19). Obstet. Gynecol. 2021, 138, 937–939. [Google Scholar] [CrossRef]
- Rut, W.; Lv, Z.; Zmudzinski, M.; Patchett, S.; Nayak, D.; Snipas, S.J.; Oualid, F.E.; Huang, T.T.; Bekes, M.; Drag, M.; et al. Activity profiling and structures of inhibitor-bound SARS-CoV-2-PLpro protease provides a framework for anti-COVID-19 drug design. Sci. Adv. 2020, 6, eabd4596. [Google Scholar] [CrossRef]
- Freitas, B.T.; Durie, I.A.; Murray, J.; Longo, J.E.; Miller, H.C.; Crich, D.; Hogan, R.J.; Tripp, R.A.; Pegan, S.D. Characterization and noncovalent inhibition of the deubiquitinase and deISGylase activity of SARS-CoV-2 papain-like protease. ACS Infect. Dis. 2020, 6, 099–109. [Google Scholar] [CrossRef]
- Fu, Z.; Huang, B.; Tang, J.; Liu, S.; Liu, M.; Ye, Y.; Liu, Z.; Xiong, Y.; Zhu, W.; Cao, D.; et al. The complex structure of GRL0617 and SARS-CoV-2 PLpro reveals a hot spot for antiviral drug discovery. Nat. Commun. 2021, 12, 488. [Google Scholar] [CrossRef] [PubMed]
- Bojkova, D.; Klann, K.; Koch, B.; Widera, M.; Krause, D.; Ciesek, S.; Cinatl, J.; Munch, C. Proteomics of SARS-CoV-2-infected host cells reveals therapy targets. Nature 2020, 583, 69–72. [Google Scholar] [CrossRef] [PubMed]
- de Loubresse, N.G.; Prokhorova, I.; Holtkamp, W.; Rodnina, M.V.; Yusupova, G.; Yusupov, M. Structural basis for the inhibition of the eukaryotic ribosome. Nature 2014, 513, 517–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivashchenko, A.A.; Dmitriev, K.A.; Vostokova, N.V.; Azarova, V.N.; Blinow, A.A.; Egorova, A.N.; Gordeev, I.G.; Ilin, A.P.; Karapetian, R.N.; Kravchenko, D.V.; et al. Avifavir for treatment of patients with moderate coronavirus disease 2019 (COVID-19): Interim results of a phase II/III multicenter randomized clinical trial. Clin. Infect. Dis. 2021, 73, 531–534. [Google Scholar] [CrossRef]
- Elfiky, A.A. Ribavirin, remdesivir, sofosbuvir, galidesivir, and tenofovir against SARS-CoV-2 RNA dependent RNA polymerase (RdRp): A molecular docking study. Life Sci. 2020, 253, 117592. [Google Scholar] [CrossRef]
- Lau, J.Y.N.; Tam, R.C.; Liang, T.J.; Hong, Z. Mechanism of action of ribavirin in the combination treatment of chronic HCV infection. Hepatology 2002, 35, 1002–1009. [Google Scholar] [CrossRef]
- Zhang, W.; Stephen, P.; Theriault, J.F.; Wang, R.; Lin, S.X. Novel coronavirus polymerase and nucleotidyl-transferase structures: Potential to target new outbreaks. J. Phys. Chem. Lett. 2020, 11, 4430–4435. [Google Scholar] [CrossRef]
- Gottlieb, R.L.; Vaca, C.E.; Paredes, R.; Mera, J.; Webb, B.J.; Perez, G.; Oguchi, G.; Ryan, P.; Nielsen, B.U.; Brown, M.; et al. Early to prevent progression to severe Covid-19 in outpatients. N. Engl. J. Med. 2022, 386, 305–315. [Google Scholar] [CrossRef]
- Bernal, A.J.; da Silva, M.M.G.; Musungaie, D.B.; Kovalchuk, E.; Gonzalez, A.; Delos Reyes, V.; Martín-Quiros, A.; Caraco, Y.; Williams-Diaz, A.; Brown, M.L.; et al. Molnupiravir for oral treatment of Covid-19 in nonhospitalized patients. N. Engl. J. Med. 2022, 386, 509–520. [Google Scholar] [CrossRef]
- Tchesnokov, E.P.; Feng, J.Y.; Porter, D.P.; Gotte, M. Mechanism of inhibition of ebola virus RNA-dependent RNA polymerase by remdesivir. Viruses 2019, 11, 326. [Google Scholar] [CrossRef] [Green Version]
- Lamb, Y.N. Remdesivir: First approval. Drugs. 2020, 80, 1355–1363. [Google Scholar] [CrossRef] [PubMed]
- Kokic, G.; Hillen, H.S.; Tegunov, D.; Dienemann, C.; Seitz, F.; Schmitzova, J.; Farnung, L.; Siewert, A.; Hobartner, C.; Cramer, P. Mechanism of SARS-CoV-2 polymerase stalling by remdesivir. Nat. Commun. 2021, 12, 279. [Google Scholar]
- Li, Y.; Zhang, H.; Luo, W.; Lam, C.W.K.; Wang, C.; Bai, L.; Wong, V.K.W.; Zhang, W.; Jiang, Z. Profiling ribonucleotide and deoxyribonucleotide pools perturbed by remdesivir in human bronchial epithelial cells. Front. Pharmacol. 2021, 12, 647280. [Google Scholar] [CrossRef] [PubMed]
- De Vito, A.; Poliseno, M.; Colpani, A.; Zauli, B.; Puci, M.V.; Santantonio, T.; Meloni, M.C.; Fois, M.; Fanelli, C.; Saderi, L.; et al. Reduced risk of death in people with SARS-CoV-2 infection treated with remdesivir: A nested case-control study. Curr. Med. Res. Opin. 2022, 13, 1–5. [Google Scholar] [CrossRef]
- Lee, T.C.; Murthy, S.; Del Corpo, O.; Senecal, J.; Butler-Laporte, G.; Sohani, Z.N.; Brophy, J.M.; McDonald, E.G. Remdesivir for the treatment of COVID-19: A systematic review and meta-analysis. Clin. Microbiol. Infect. 2022, 28, 1203–1210. [Google Scholar] [CrossRef]
- Rezagholizadeh, A.; Khiali, S.; Sarbakhsh, P.; Entezari-Maleki, T. Remdesivir for treatment of COVID-19: An updated systematic review and meta-analysis. Eur. J. Pharmacol. 2021, 897, 173926. [Google Scholar] [CrossRef]
- Pan, H.; Peto, R.; Henao-Restrepo, A.M.; Preziosi, M.P.; Sathiyamoorthy, V.; Abdool Karim, Q.; Alejandria, M.M.; Hernandez Garcia, C.; Kieny, M.P.; Malekzadeh, R. Repurposed antiviral drugs for Covid-19—Interim WHO solidarity trial results. N. Engl. J. Med. 2021, 384, 497–511. [Google Scholar]
- Colaneri, M.; Amarasinghe, N.; Rezzonico, L.; Pieri, T.C.; Segalini, E.; Sambo, M.; Roda, S.; Meloni, F.; Gregorini, M.; Rampino, T.; et al. Early remdesivir to prevent severe COVID-19 in recipients of solid organ transplant: A real-life study from Northern Italy. Int. J. Infect. Dis. 2022, 121, 157–160. [Google Scholar] [CrossRef]
- Pourkarim, F.; Pourtaghi-Anvarian, S.; Rezaee, H. Molnupiravir: A new candidate for COVID-19 treatment. Pharmacol. Res. Perspect. 2022, 10, e00909. [Google Scholar] [CrossRef]
- Hashemian, S.M.R.; Pourhanifeh, M.H.; Hamblin, M.R.; Shahrzad, M.K.; Mirzaei, H. RdRp inhibitors and COVID-19: Is molnupiravir a good option? Biomed. Pharmacother. 2022, 146, 112517. [Google Scholar] [CrossRef]
- Yu, P.; Zhu, J.; Zhang, Z.; Han, Y. A familial cluster of infection associated with the 2019 novel coronavirus indicating possible person-to-person transmission during the incubation period. J. Infect. Dis. 2020, 221, 1757–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, W.; Luan, X.; Li, Z.; Zhou, Z.; Wang, Q.; Gao, M.; Wang, X.; Zhou, F.; Shi, J.; You, E.; et al. Structural basis for inhibition of the SARS-CoV-2 RNA polymerase by suramin. Nat. Struct. Mol. Biol. 2021, 28, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Hohenegger, M.; Matyash, M.; Poussu, K.; Herrmann-Frank, A.; Sarkozi, S.; Lehmann-Horn, F.; Freissmuth, M. Activation of the skeletal muscle ryanodine receptor by suramin and suramin analogs. Mol. Pharmacol. 1996, 50, 1443–1453. [Google Scholar]
- Wiedemar, N.; Hauser, D.A.; Maser, P. 100 years of suramin. Antimicrob. Agents Chemother. 2020, 64, e01168-19. [Google Scholar] [CrossRef]
- Parveen, N.; Lin, Y.L.; Khan, M.I.; Chou, R.H.; Sun, C.M.; Yu, C. Suramin derivatives play an important role in blocking the interaction between FGF1 and FGFRD2 to inhibit cell proliferation. Eur. J. Med. Chem. 2020, 206, 112656. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Yin, X.; Meng, X.; Chan, J.F.W.; Ye, Z.W.; Riva, L.; Pache, L.; Chan, C.C.Y.; Lai, P.M.; Chan, C.C.S.; et al. Clofazimine broadly inhibits coronaviruses including SARS-CoV-2. Nature 2021, 593, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Malone, B.; Llewellyn, E.; Grasso, M.; Shelton, P.M.M.; Olinares, P.D.B.; Maruthi, K.; Eng, E.T.; Vatandaslar, H.; Chait, B.T.; et al. Structural basis for helicase-polymerase coupling in the SARS-CoV-2 replication-transcription complex. Cell 2020, 182, 1560–1573. [Google Scholar] [CrossRef]
- Hecel, A.; Ostrowska, M.; Stokowa-Soltys, K.; Watly, J.; Dudek, D.; Miller, A.; Potocki, S.; Matera-Witkiewicz, A.; Dominguez-Martin, A.; Kozlowski, H.; et al. Zinc(II)—The overlooked eminence grise of chloroquine’s fight against COVID-19? Pharmaceuticals 2020, 13, 228. [Google Scholar] [CrossRef]
- You, K.; Huang, Q.; Yu, C.; Shen, B.; Sevilla, C.; Shi, M.; Hermjakob, H.; Chen, Y.; Li, T. PhaSepDB: A database of liquid-liquid phase separation related proteins. Nucleic Acids Res. 2020, 48, D354–D359. [Google Scholar] [CrossRef]
- Zhang, H.; Ji, X.; Li, P.; Liu, C.; Lou, J.; Wang, Z.; Wen, W.; Xiao, Y.; Zhang, M.; Zhu, X. Liquid-liquid phase separation in biology: Mechanisms, physiological functions and human diseases. Sci. China Life Sci. 2020, 63, 953–985. [Google Scholar] [CrossRef]
- Hyman, A.A.; Weber, C.A.; Julicher, F. Liquid-liquid phase separation in biology. Annu. Rev. Cell Dev. Biol. 2014, 30, 39–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Yu, Y.; Sun, L.M.; Xing, J.Q.; Li, T.; Zhu, Y.; Wang, M.; Yu, Y.; Xue, W.; Xia, T.; et al. GCG inhibits SARS-CoV-2 replication by disrupting the liquid phase condensation of its nucleocapsid protein. Nat. Commun. 2021, 12, 2114. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.R.; Kaminski, J.J.; Kurt-Jones, E.A.; Fitzgerald, K.A. Pattern recognition receptors and the innate immune response to viral infection. Viruses 2011, 3, 920–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, C.M.; Foti, L. Anti-SARS-CoV-2 and anti-cytokine storm neutralizing antibody therapies against COVID-19: Update, challenges, and perspectives. Int. Immunopharmacol. 2021, 99, 108036. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.W.; Wang, T.; Zhang, X.Y.; Chen, H.L.; Yu, H.J.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef]
- Chen, Y.; Klein, S.L.; Garibaldi, B.T.; Li, H.; Wu, C.; Osevala, N.M.; Li, T.; Margolick, J.B.; Pawelec, G.; Leng, S.X. Aging in COVID-19: Vulnerability, immunity and intervention. Ageing Res. Rev. 2021, 65, 101205. [Google Scholar] [CrossRef]
- Kaly, L.; Rosner, M.I. Tocilizumab—A novel therapy for non-organ-specific autoimmune diseases. Best Pract. Res. Clin. Rheumatol. 2012, 26, 157–165. [Google Scholar] [CrossRef]
- Khan, F.A.; Stewart, I.; Fabbri, L.; Moss, S.; Robinson, K.; Smyth, A.R.; Jenkins, G. Systematic review and meta-analysis of anakinra, sarilumab, siltuximab and tocilizumab for COVID-19. Thorax 2021, 76, 907–919. [Google Scholar] [CrossRef]
- Mazzitelli, M.; Arrighi, E.; Serapide, F.; Pelle, M.C.; Tassone, B.; Lionello, R.; Marrazzo, G.; Lagana, D.; Costanzo, F.S.; Matera, G.; et al. Use of subcutaneous tocilizumab in patients with COVID-19 pneumonia. J. Med. Virol. 2021, 93, 32–34. [Google Scholar] [CrossRef]
- Navas, N.; Hermosilla, J.; Torrente-Lopez, A.; Hernandez-Jimenez, J.; Cabeza, J.; Perez-Robles, R.; Salmeron-Garcia, A. Use of subcutaneous tocilizumab to prepare intravenous solutions for COVID-19 emergency shortage: Comparative analytical study of physicochemical quality attributes. J. Pharm. Anal. 2020, 10, 532–545. [Google Scholar] [CrossRef]
- Di Giambenedetto, S.; Ciccullo, A.; Borghetti, A.; Gambassi, G.; Landi, F.; Visconti, E.; Dal Verme, L.Z.; Bernabei, R.; Tamburrini, E.; Cauda, R.; et al. Off-label use of tocilizumab in patients with SARS-CoV-2 infection. J. Med. Virol. 2020, 92, 1787–1788. [Google Scholar] [CrossRef] [PubMed]
- Khiali, S.; Rezagholizadeh, A.; Entezari-Maleki, T. A comprehensive review on sarilumab in COVID-19. Expert Opin. Biol. Ther. 2021, 21, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Gremese, E.; Cingolani, A.; Bosello, S.L.; Alivernini, S.; Tolusso, B.; Perniola, S.; Landi, F.; Pompili, M.; Murri, R.; Santoliquido, A.; et al. Sarilumab use in severe SARS-CoV-2 pneumonia. EClinicalMedicine 2020, 27, 100553. [Google Scholar] [CrossRef] [PubMed]
- Benucci, M.; Giannasi, G.; Cecchini, P.; Gobbi, F.L.; Damiani, A.; Grossi, V.; Infantino, M.; Manfredi, M. COVID-19 pneumonia treated with Sarilumab: A clinical series of eight patients. J. Med. Virol. 2020, 92, 2368–2370. [Google Scholar] [CrossRef]
- Marino, A.; Munafo, A.; Augello, E.; Bellanca, C.M.; Bonomo, C.; Ceccarelli, M.; Musso, N.; Cantarella, G.; Cacopardo, B.; Bernardini, R. Sarilumab administration in COVID-19 patients: Literature review and considerations. Infect. Dis. Rep. 2022, 14, 360–371. [Google Scholar] [CrossRef]
- Lescure, F.X.; Honda, H.; Fowler, R.A.; Lazar, J.S.; Shi, G.; Wung, P.; Patel, N.; Hagino, O. Sarilumab in patients admitted to hospital with severe or critical COVID-19: A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Respir. Med. 2021, 9, 522–532. [Google Scholar] [CrossRef]
- Mariette, F.; Hermine, O.; Tharaux, P.L.; Resche-Rigon, M.; Porcher, R.; Ravaud, P. Sarilumab in adults hospitalised with moderate-to-severe COVID-19 pneumonia (CORIMUNO-SARI-1): An open-label randomised controlled trial. Lancet Rheumatol. 2022, 4, e24–e32. [Google Scholar] [CrossRef]
- Luo, W.; Ding, R.; Guo, X.; Zhan, T.; Tang, T.; Fan, R.; Wang, Y. Clinical data mining reveals Gancao-Banxia as a potential herbal pair against moderate COVID-19 by dual binding to IL-6/STAT3. Comput. Biol. Med. 2022, 145, 105457. [Google Scholar] [CrossRef]
- Monteagudo, L.A.; Boothby, A.; Gertner, E. Continuous intravenous anakinra infusion to calm the cytokine storm in macrophage activation syndrome. ACR Open Rheumatol. 2020, 2, 276–282. [Google Scholar] [CrossRef]
- Huet, T.; Beaussier, H.; Voisin, O.; Jouveshomme, S.; Dauriat, G.; Lazareth, I.; Sacco, E.; Naccache, J.M.; Bezie, Y.; Laplanche, S.; et al. Anakinra for severe forms of COVID-19: A cohort study. Lancet Rheumatol. 2020, 2, E393–E400. [Google Scholar] [CrossRef]
- Florescu, D.F.; Kalil, A.C. Janus kinase inhibitors for the treatment of hospitalized patients with COVID-19. Curr. Opin. Crit. Care. 2021, 27, 493–496. [Google Scholar] [CrossRef] [PubMed]
- de Luca, G.; Cavalli, G.; Campochiaro, C.; Della-Torre, E.; Angelillo, P.; Tomelleri, A.; Boffini, N.; Tentori, S.; Mette, F.; Farina, N.; et al. GM-CSF blockade with mavrilimumab in severe COVID-19 pneumonia and systemic hyperinflammation: A single-centre, prospective cohort study. Lancet Rheumatol. 2020, 2, E465–E473. [Google Scholar] [CrossRef]
- Nilsson, B.; Persson, B.; Eriksson, O.; Fromell, K.; Hultström, M.; Frithiof, R.; Lipcsey, M.; Huber-Lang, M.; Ekdahl, K.N. How the innate immune system of the blood contributes to systemic pathology in COVID-19-induced ARDS and provides potential targets for treatment. Front. Immunol. 2022, 13, 840137. [Google Scholar] [CrossRef] [PubMed]
- Diurno, F.; Numis, F.G.; Porta, G.; Cirillo, F.; Maddaluno, S.; Ragozzino, A.; De Negri, P.; Di Gennaro, C.; Pagano, A.; Allegorico, E.; et al. Eculizumab treatment in patients with COVID-19: Preliminary results from real life ASL Napoli 2 Nord experience. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4040–4047. [Google Scholar]
- Xu, X.; Han, M.; Li, T.; Sun, W.; Wang, D.; Fu, B.; Zhou, Y.; Zheng, X.; Yang, Y.; Li, X.; et al. Effective treatment of severe COVID-19 patients with tocilizumab. Proc. Natl. Acad. Sci. USA 2020, 117, 10970–10975. [Google Scholar] [CrossRef]
- Qian, J.; Xu, H.; Lv, D.; Liu, W.; Chen, E.; Zhou, Y.; Wang, Y.; Ying, K.; Fan, X. Babaodan controls excessive immune responses and may represent a cytokine-targeted agent suitable for COVID-19 treatment. Biomed. Pharmacother. 2021, 139, 111586. [Google Scholar] [CrossRef]
- Edalatifard, M.; Akhtari, M.; Salehi, M.; Naderi, Z.; Jamshidi, A.; Mostafaei, S.; Najiafizadeh, S.R.; Farhadi, E.; Jalili, N.; Esfahani, M.; et al. Intravenous methylprednisolone pulse as a treatment for hospitalised severe COVID-19 patients: Results from a randomised controlled clinical trial. Eur. Respir. J. 2020, 56, 2002808. [Google Scholar] [CrossRef]
- Antonov, V.N.; Ignatova, G.L.; Pribytkova, O.V.; Sleptsova, S.S.; Strebkova, E.A.; Khudyakova, E.A.; Simakov, A.I.; Rabets, S.Y.; Tikhonova, E.P.; Kurmaeva, D.Y.; et al. Experience of olokizumab use in COVID-19 patients. Ter. Arkhiv 2020, 92, 148–154. [Google Scholar] [CrossRef]
- Buryachkovskaya, L.; Lomakin, N.; Melkumyants, A.; Docenko, J.; Serebruany, V. Impact of olokizumab on platelets, leukocytes and erythrocytes during mild COVID-19. Rev. Cardiovasc. Med. 2021, 22, 549–551. [Google Scholar] [CrossRef]
- Meira, F.; Albiach, L.; Carbonell, C.; Martín-Oterino, J.A.; Martin-Ordiales, M.; Linares, L.; Munell, I.M.; Aguero, D.; Ambrosioni, J.; Bodro, M.; et al. Experience with the use of siltuximab in patients with SARS-CoV-2 infection. Rev. Esp. Quimioter. 2021, 34, 337–341. [Google Scholar] [CrossRef]
- Vaidya, G.; Czer, L.S.C.; Kobashigawa, J.; Kittleson, M.; Patel, J.; Chang, D.; Kransdorf, E.; Shikhare, A.; Tran, H.; Vo, A.; et al. Successful treatment of severe COVID-19 pneumonia with clazakizumab in a heart transplant recipient: A case report. Transplant. Proc. 2020, 52, 2711–2714. [Google Scholar] [CrossRef] [PubMed]
- Caracciolo, M.; Macheda, S.; Labate, D.; Tescione, M.; La Scala, S.; Vadala, E.; Squillaci, R.; D’Aleo, F.; Morabito, A.; Garreffa, C.; et al. Case report: Canakinumab for the treatment of a patient with COVID-19 acute respiratory distress syndrome. Front. Immunol. 2020, 11, 1942. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Wang, F.; Huang, Z.; Chen, X.; Wang, Y.L. Perspectives on anti-IL-1 inhibitors as potential therapeutic interventions for severe COVID-19. Cytokine 2021, 143, 155544. [Google Scholar] [CrossRef] [PubMed]
- Franzetti, M.; Pozzetti, U.; Carugati, M.; Pandolfo, A.; Molteni, C.; Faccioli, P.; Castaldo, G.; Longoni, E.; Ormas, V.; Lemoli, V.; et al. Interleukin-1 receptor antagonist anakinra in association with anakinra in severe COVID-19: A case report. Int. J. Infect. Dis. 2020, 97, 215–218. [Google Scholar] [CrossRef]
- Feldmann, M. Anti-TNFα therapy of rheumatoid arthritis: What have we learned? Annu. Rev. Immunol. 2001, 19, 163–196. [Google Scholar] [CrossRef]
- Kennedy, N.A.; Goodhand, J.R.; Bewshea, C.; Nice, R.; Chee, D.; Lin, S.; Chanchlani, N.; Butterworth, J.; Cooney, R.; Croft, N.M.; et al. Anti-SARS-CoV-2 antibody responses are attenuated in patients with IBD treated with infliximab. Gut 2021, 70, 865–875. [Google Scholar] [CrossRef]
- Onsun, N.; Gunes, B.; Yabaci, A. Retention and survival rate of etanercept in psoriasis over 15 years and patient outcomes during the COVID-19 pandemic: The real-world experience of a single center. Dermatol. Ther. 2021, 34, e14623. [Google Scholar] [CrossRef]
- Lee, J.M.; Lee, S.J. Olfactory and gustatory dysfunction in a COVID-19 patient with ankylosing spondylitis treated with etanercept: Case report. J. Korean Med. Sci. 2020, 35, e201. [Google Scholar] [CrossRef]
- Surdea-Blaga, T.; David, L.; Pop, A.; Tantau, M.; Dumitrascu, D.L. Clinical and manometric characteristics of patients with achalasia: One disease with three presentations or three diseases with one presentation? J Gastrointest. Liver Dis. 2020, 29, 501–508. [Google Scholar] [CrossRef]
- Fakharian, A.; Barati, S.; Mohamadi, M.; Dastan, F. Successful management of COVID-19 with adalimumab in a post-coronary artery bypass graft surgery patient. J. Cardiothorac. Vasc. Anesth. 2022, 36, 1115–1117. [Google Scholar] [CrossRef]
- Ahmed, F. A network-based analysis reveals the mechanism underlying vitamin D in suppressing cytokine storm and virus in SARS-CoV-2 infection. Front. Immunol. 2020, 11, 590459. [Google Scholar] [CrossRef] [PubMed]
- Cure, E.; Kucuk, A.; Cure, M.C. Can emapalumab be life saving for refractory, recurrent, and progressive cytokine storm caused by COVID-19, which is resistant to anakinra, tocilizumab, and Janus kinase inhibitors. Indian J. Pharmacol. 2021, 53, 226–228. [Google Scholar] [PubMed]
- Cao, Y.; Wei, J.; Zou, L.; Jiang, T.; Wang, G.; Chen, L.; Huang, L.; Meng, F.; Huang, L.; Wang, N.; et al. Ruxolitinib in treatment of severe coronavirus disease 2019 (COVID-19): A multicenter, single-blind, randomized controlled trial. J. Allergy Clin. Immun. 2020, 146, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Yang, X.O. TH17 responses in cytokine storm of COVID-19: An emerging target of JAK2 inhibitor fedratinib. J. Microbiol. Immunol. 2020, 53, 368–370. [Google Scholar] [CrossRef]
- Jacobs, J.; Clark-Snustad, K.; Lee, S. Case report of a SARS-CoV-2 infection in a patient with ulcerative colitis on tofacitinib. Inflamm. Bowel Dis. 2020, 26, E64. [Google Scholar] [CrossRef]
- Kalil, A.C.; Patterson, T.F.; Mehta, A.K.; Tomashek, K.M.; Wolfe, C.R.; Ghazaryan, V.; Marconi, V.C.; Ruiz-Palacios, G.M.; Hsieh, L.; Kline, S.; et al. Baricitinib plus remdesivir for hospitalized adults with covid-19. N. Engl. J. Med. 2021, 384, 795–807. [Google Scholar] [CrossRef]
- Richardson, P.; Griffin, I.; Tucker, C.; Smith, D.; Oechsle, O.; Phelan, A.; Stebbing, J. Baricitinib as potential treatment for 2019-nCoV acute respiratory disease. Lancet 2020, 395, E30–E31. [Google Scholar] [CrossRef] [Green Version]
- Hoang, T.N.; Pino, M.; Boddapati, A.K.; Viox, E.G.; Starke, C.E.; Upadhyay, A.A.; Gumber, S.; Nekorchuk, M.; Busman-Sahay, K.; Strongin, Z.; et al. Baricitinib treatment resolves lower-airway macrophage inflammation and neutrophil recruitment in SARS-CoV-2-infected rhesus macaques. Cell 2021, 184, 460–475. [Google Scholar] [CrossRef]
- Qin, Y.Y.; Zhou, Y.H.; Lu, Y.Q.; Sun, F.; Yang, S.; Harypursat, V.; Chen, Y.K. Effectiveness of glucocorticoid therapy in patients with severe coronavirus disease 2019: Protocol of a randomized controlled trial. Chin. Med. J. 2020, 133, 1080–1086. [Google Scholar] [CrossRef]
- Veronese, N.; Demurtas, J.; Yang, L.; Tonelli, R.; Barbagallo, M.; Lopalco, P.; Lagolio, E.; Celotto, S.; Pizzol, D.; Zou, L.; et al. Use of corticosteroids in coronavirus disease 2019 pneumonia: A systematic review of the literature. Front. Med. 2020, 7, 170. [Google Scholar] [CrossRef]
- Elizagaray, M.L.; Mazitelli, I.; Pontoriero, A.; Baumeister, E.; Docena, G.; Raimondi, C.; Correger, E.; Rumbo, M. Lidocaine reinforces the anti-inflammatory action of dexamethasone on myeloid and epithelial cells activated by inflammatory cytokines or SARS-CoV-2 infection. Biomed. J. 2022; in press. [Google Scholar] [CrossRef]
- Reis, G.; Moreira-Silva, E.A.; Silva, D.C.M.; Thabane, L.; Milagres, A.C.; Ferreira, T.S.; Dos Santos, C.V.Q.; Campos, V.H.; Nogueira, A.M.R.; de Almeida, A.P.F.G.; et al. Effect of early treatment with fluvoxamine on risk of emergency care and hospitalisation among patients with COVID-19: The TOGETHER randomised, platform clinical trial. Lancet Glob. Health 2022, 10, e42–e51. [Google Scholar] [CrossRef]
- Million, M.; Lagier, J.C.; Gautret, P.; Colson, P.; Fournier, P.E.; Amrane, S.; Hocquart, M.; Mailhe, M.; Esteves-Vieira, V.; Doudier, B.; et al. Early treatment of COVID-19 patients with hydroxychloroquine and azithromycin: A retrospective analysis of 1061 cases in Marseille, France. Travel Med. Infect. Dis. 2020, 35, 101738. [Google Scholar] [CrossRef] [PubMed]
- Pani, A.; Lauriola, M.; Romandini, A.; Scaglione, F. Macrolides and viral infections: Focus on azithromycin in COVID-19 pathology. Int. J. Antimicrob. Agents 2020, 56, 106053. [Google Scholar] [CrossRef]
- Zarezadeh, M.; Khorshidi, M.; Emami, M.; Janmohammadi, P.; Kord-varkaneh, H.; Mousavi, S.M.; Mohammed, S.H.; Saedisomeolia, A. Melatonin supplementation and pro-inflammatory mediators: A systematic review and meta-analysis of clinical trials. Eur. J. Nutr. 2020, 59, 1803–1813. [Google Scholar] [CrossRef]
- Fenizia, C.; Galbiati, S.; Vanetti, C.; Vago, R.; Clerici, M.; Tacchetti, C.; Daniele, T. Cyclosporine a inhibits viral infection and release as well as cytokine production in lung cells by three SARS-CoV-2 variants. Microbiol. Spectr. 2022, 10, e01504–e01521. [Google Scholar] [CrossRef]
- Tang, N.; Bai, H.; Chen, X.; Gong, J.; Li, D.; Sun, Z. Anticoagulant treatment is associated with decreased mortality in severe coronavirus disease 2019 patients with coagulopathy. J. Thromb. Haemost. 2020, 18, 1094–1099. [Google Scholar] [CrossRef] [Green Version]
- DE Vito, A.; Saderi, L.; Fiore, V.; Geremia, N.; Princic, E.; Fanelli, C.; Muredda, A.A.; Panu Napodano, C.; Moi, G.; Maida, I.; et al. Early treatment with low-molecular-weight heparin reduces mortality rate in SARS-CoV-2 patients. Panminerva Med. 2022. [Google Scholar] [CrossRef]
- Scott, H.; Zahra, A.; Fernandes, R.; Fries, B.C.; Thode, H.C.; Singer, A.J. Bacterial Infections and death among patients with COVID-19 versus non COVID-19 patients with pneumonia. Am. J. Emerg. Med. 2022, 51, 1–5. [Google Scholar] [CrossRef]
- Cohen, R.; Finn, T.; Babushkin, F.; Geller, K.; Alexander, H.; Shapiro, M.; Uda, M.; Mostrchy, A.R.; Amash, R.; Shimoni, Z.; et al. High rate of bacterial respiratory tract co-infections upon admission amongst moderate to severe COVID-19 patients. Infect. Dis. 2022, 54, 134–144. [Google Scholar] [CrossRef]
- Hughes, S.; Mughal, N.; Moore, L.S.P. Procalcitonin to guide antibacterial prescribing in patients hospitalised with COVID-19. Antibiotics 2021, 10, 1119. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, M.; Marino, A.; Pulvirenti, S.; Coco, V.; Busa, B.; Nunnari, G.; Cacopardo, B.S. Bacterial and fungal co-infections and superinfections in a cohort of COVID-19 patients: Real-life data from an Italian third level hospital. Infect. Dis. Rep. 2022, 14, 372–382. [Google Scholar] [CrossRef] [PubMed]
- El-Sokkary, R.; Uysal, S.; Erdem, H.; Kullar, R.; Pekok, A.U.; Amer, F.; Grgic, S.; Carevic, B.; El-Kholy, A.; Liskova, A.; et al. Profiles of multidrug-resistant organisms among patients with bacteremia in intensive care units: An international ID-IRI survey. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 2323–2334. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The infection of SARS-CoV-2 into human cells. (A) Structure of SARS-CoV-2; (B) structure of the S protein. S1 includes an NTD and RBD. S2 plays a role in promoting cell entry in membrane fusion and contains an FP domain, two seven-step repeat domains (HR1 and HR2), a TM domain, and a CT domain; (C) schematic representation of the infection of SARS-CoV-2 into human cells and toxic effects. S protein: spike protein; NTD: N-terminal domain; RBD: receptor-binding domain; FP: fusion peptide; TM: transmembrane; CT: C-terminal; SARS-CoV-2: severe acute respiratory syndrome coronavirus 2.
Figure 1.
The infection of SARS-CoV-2 into human cells. (A) Structure of SARS-CoV-2; (B) structure of the S protein. S1 includes an NTD and RBD. S2 plays a role in promoting cell entry in membrane fusion and contains an FP domain, two seven-step repeat domains (HR1 and HR2), a TM domain, and a CT domain; (C) schematic representation of the infection of SARS-CoV-2 into human cells and toxic effects. S protein: spike protein; NTD: N-terminal domain; RBD: receptor-binding domain; FP: fusion peptide; TM: transmembrane; CT: C-terminal; SARS-CoV-2: severe acute respiratory syndrome coronavirus 2.

Figure 2.
Potential therapeutic targets for the treatment of COVID-19. ACE2: angiotensin-converting enzyme 2; RNA: ribonucleic acid.
Figure 2.
Potential therapeutic targets for the treatment of COVID-19. ACE2: angiotensin-converting enzyme 2; RNA: ribonucleic acid.

Figure 3.
S protein and RBD process. S protein: spike protein; RBD: receptor-binding domain; ACE2: angiotensin-converting enzyme 2; PD: protease domain; NTD: N-terminal domain; CTD: C-terminal domain.
Figure 3.
S protein and RBD process. S protein: spike protein; RBD: receptor-binding domain; ACE2: angiotensin-converting enzyme 2; PD: protease domain; NTD: N-terminal domain; CTD: C-terminal domain.

Figure 4.
Replication of SARS-CoV-2 in host cells. Mpro: main protease; pp1a: polyprotein 1a; pp1ab: polyprotein 1ab; PLpro: papain-like proteinase; RNA: ribonucleic acid; RdRp: RNA-dependent RNA polymerase.
Figure 4.
Replication of SARS-CoV-2 in host cells. Mpro: main protease; pp1a: polyprotein 1a; pp1ab: polyprotein 1ab; PLpro: papain-like proteinase; RNA: ribonucleic acid; RdRp: RNA-dependent RNA polymerase.

Figure 5.
Schematic representation of cytokine storm. ① A large number of viruses infect host cells and stimulate macrophages to fight the virus. ② Phagocytes can secrete cytokines to attract white blood cells to inhibit SARS-CoV-2. ③ Leukocytes secrete specific cytokines to stimulate cell pathways to attract the arrival of dendritic cells to reduce the number of viruses. ④ Dendritic cells degrade the virus into specific proteins and nucleic acids. ⑤ Dendritic cells also secrete IL-12 and IL-18, which activate signal molecules to stimulate T-cell proliferation and enhance immune effects. ⑥ Virus-specific proteins and nucleic acids induce B cells to differentiate into effector B cells that can secrete specific antibodies to fight the virus. IL: interleukin; IFN: interferon; TNF: tumor necrosis factor.
Figure 5.
Schematic representation of cytokine storm. ① A large number of viruses infect host cells and stimulate macrophages to fight the virus. ② Phagocytes can secrete cytokines to attract white blood cells to inhibit SARS-CoV-2. ③ Leukocytes secrete specific cytokines to stimulate cell pathways to attract the arrival of dendritic cells to reduce the number of viruses. ④ Dendritic cells degrade the virus into specific proteins and nucleic acids. ⑤ Dendritic cells also secrete IL-12 and IL-18, which activate signal molecules to stimulate T-cell proliferation and enhance immune effects. ⑥ Virus-specific proteins and nucleic acids induce B cells to differentiate into effector B cells that can secrete specific antibodies to fight the virus. IL: interleukin; IFN: interferon; TNF: tumor necrosis factor.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Meng, J.; Li, R.; Zhang, Z.; Wang, J.; Huang, Q.; Nie, D.; Fan, K.; Guo, W.; Zhao, Z.; Han, Z. A Review of Potential Therapeutic Strategies for COVID-19. Viruses 2022, 14, 2346. https://doi.org/10.3390/v14112346
AMA Style
Meng J, Li R, Zhang Z, Wang J, Huang Q, Nie D, Fan K, Guo W, Zhao Z, Han Z. A Review of Potential Therapeutic Strategies for COVID-19. Viruses. 2022; 14(11):2346. https://doi.org/10.3390/v14112346
Chicago/Turabian StyleMeng, Jiajia, Ruijiao Li, Zhiqi Zhang, Jie Wang, Qingwen Huang, Dongxia Nie, Kai Fan, Wenbo Guo, Zhihui Zhao, and Zheng Han. 2022. "A Review of Potential Therapeutic Strategies for COVID-19" Viruses 14, no. 11: 2346. https://doi.org/10.3390/v14112346
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.