
Development of a T Cell-Based COVID-19 Vaccine Using a Live Attenuated Influenza Vaccine Viral Vector

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses and Peptides
2.1.1. Cells
2.1.2. Viruses
2.1.3. Peptides
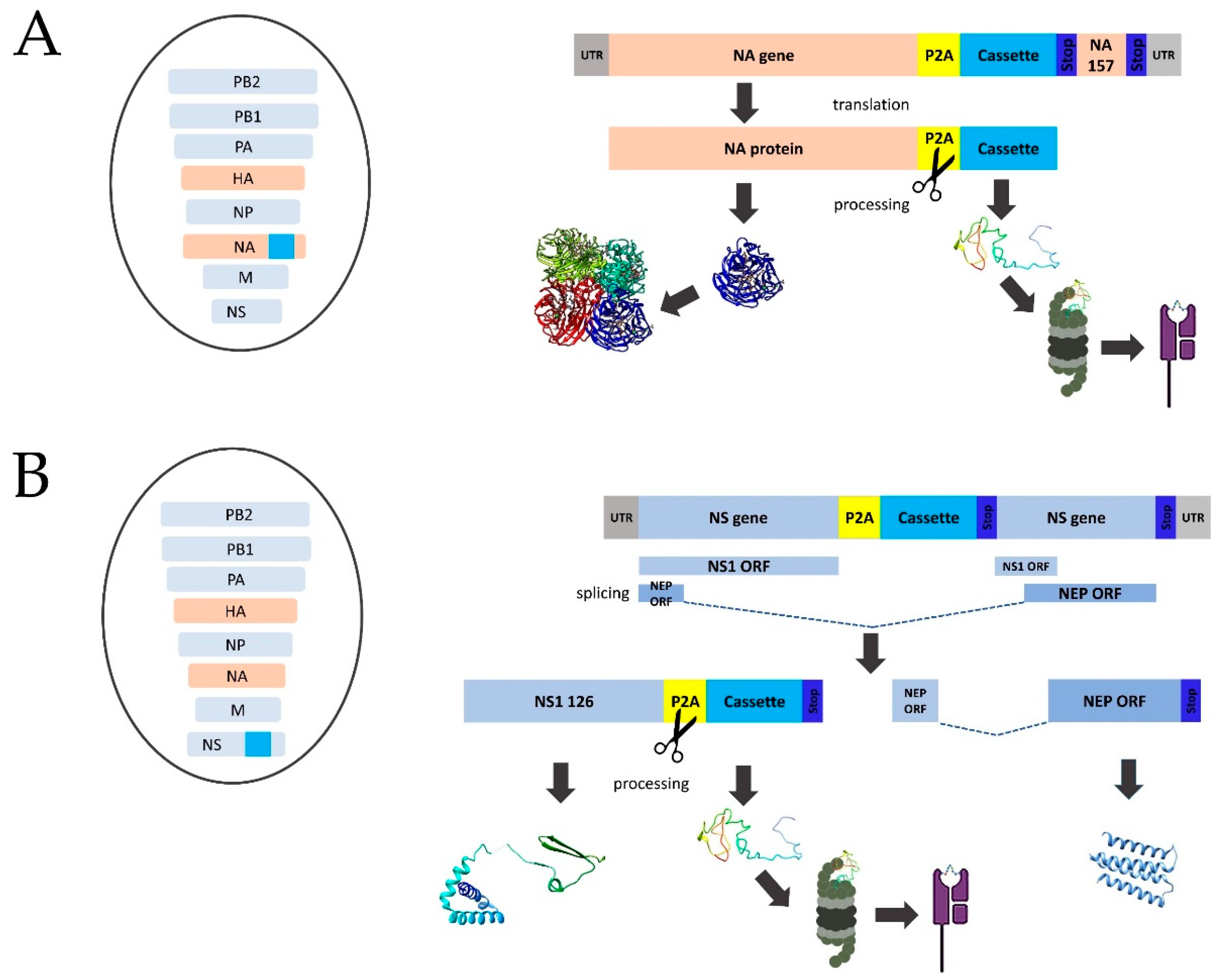
2.2. Designing of SARS-CoV-2 T-Cell Cassettes for Insertion into LAIV Genome
2.3. Generation of Recombinant LAIV Viruses Expressing SARS-CoV-2 Polyepitope Cassettes
2.4. Assessment of T-Cell Activation by Recombinant LAIV Viruses Using PBMCs of COVID-19 Convalescents
2.5. Immunization of HLA-A2.1 Transgenic Mice with LAIV/SARS-CoV-2 Vaccine Candidates
2.6. Immunization and Viral Challenge of Golden Syrian Hamsters
2.7. Assessment of Virus-Specific Antibody and Cellular Immune Responses
2.7.1. Virus-Specific Antibody Immune Responses
2.7.2. Assessment of Cellular Responses by ICS Assay
2.7.3. Assessment of Cellular Responses by ELISpot Assay
2.8. Histopathological Studies
- (a)
- Airway pathology comprised of tree parameters: % airway affected (0—none, 1—<10%, 2—10–25%, 3—25–50%, 4—50+%); airway severity (0—minimal peribronchial/peribronchiolar mononuclear infiltrates, 1—mild peribronchitis/bronchiolitis, 2—mild to moderate mononuclear to mixed peribronchiolitis/lumens contain low numbers of inflammatory cells/multifocal single cell necrosis of airway epithelium, 3—moderate to marked mixed peribronchiolitis/large foci of bronchiolar epithelial necrosis/occasional atypical or multinucleated cells; 4—marked bronchiolitis and widespread epithelial necrosis +/− rupture of bronchiolar epithelium, and/or frequent atypical/syncytial cells); and bronchiolar epithelial hyperplasia (0—none, 1—sporadic bronchiolar epithelial hyperplasia < 10% section’s airways, 2—mild to moderate bronchiolar epithelial hyperplasia 10–25% section’s airways, 3 -widespread bronchiolar epithelial hyperplasia and/or multinucleated syncytial cells taking up 25+% section’s airways).
- (b)
- Lung/alveolar pathology comprised of tree parameters: % alveoli affected (0—none, 1—<10%, 2—10–25%, 3—25–50%, 4—50+%); alveolar severity (0—within normal margins (rare/minimal peribronchial/peribronchiolar mononuclear infiltrates), 1—mild peribronchiolar primary mononuclear inflammatory infiltrates, extending into adjacent alveolar septa/spaces, 2—mild to moderate, mononuclear to mixed inflammation (>3 cell layer), expands alveolar septa or spaces/obscures normal septal architecture, 3—moderate mixed interstitial inflammation, and/or alveolar damage characterized by type I pneumocyte necrosis/loss with replacement by hemorrhage, fibrin, edema, necrotic debris (reminiscent of hyaline membranes) and/or scattered atypical/syncytial cells, 4—marked alveolar inflammation (mixed), alveolar septal damage (all above) + loss of normal septal architecture with frequent syncytial cells); and type II pneumocyte hyperplasia (0—none, 1—scattered type II pneumocyte hyperplasia taking up <10% of the section, 2—mild to moderate type II pneumocyte hyperplasia taking up 10–15% + atypical multinucleated cells, 3—widespread type II pneumocyte hyperplasia taking up 25+% of the section).
- (c)
- Vascular damage comprised of two parameters: % vessels affected (0—none, 1—<10%, 2—10–25%, 3—25–50%, 4—50+%) and vascular/perivascular lesions (0—none, 1—multifocal perivascular edema/mild mononuclear perivascular inflammation, 2—moderate mononuclear to mixed perivascular inflammation, edema or fibrin with leukocytes occasionally transmigrating the vessel wall/multifocal endotheliitis, 3—severe mixed perivascular infiltration, expanding/replacing vessel wall and/or marked frequent endotheliitis).
2.9. Statistical Analysis
3. Results
3.1. Selection of SARS-CoV-2 Fragments Enriched with Conserved T-Cell Epitopes and Generation of Recombinant LAIV/SARS-CoV-2 Viruses
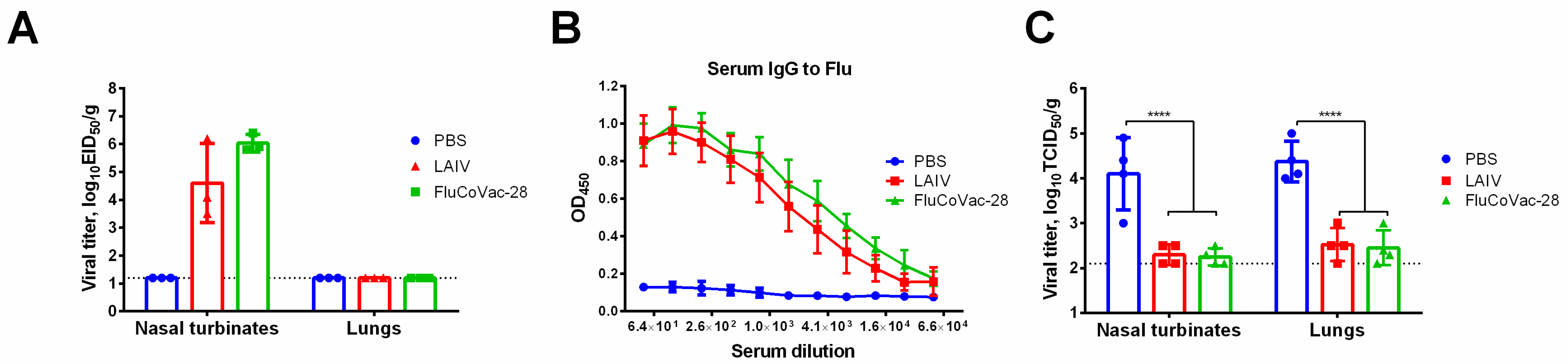
3.2. Replicative Properties of Recombinant LAIV Viruses In Vitro and In Vivo
3.3. Immunogenicity of the Recombinant LAIV-SARS-CoV-2 Viruses in Transgenic Mice
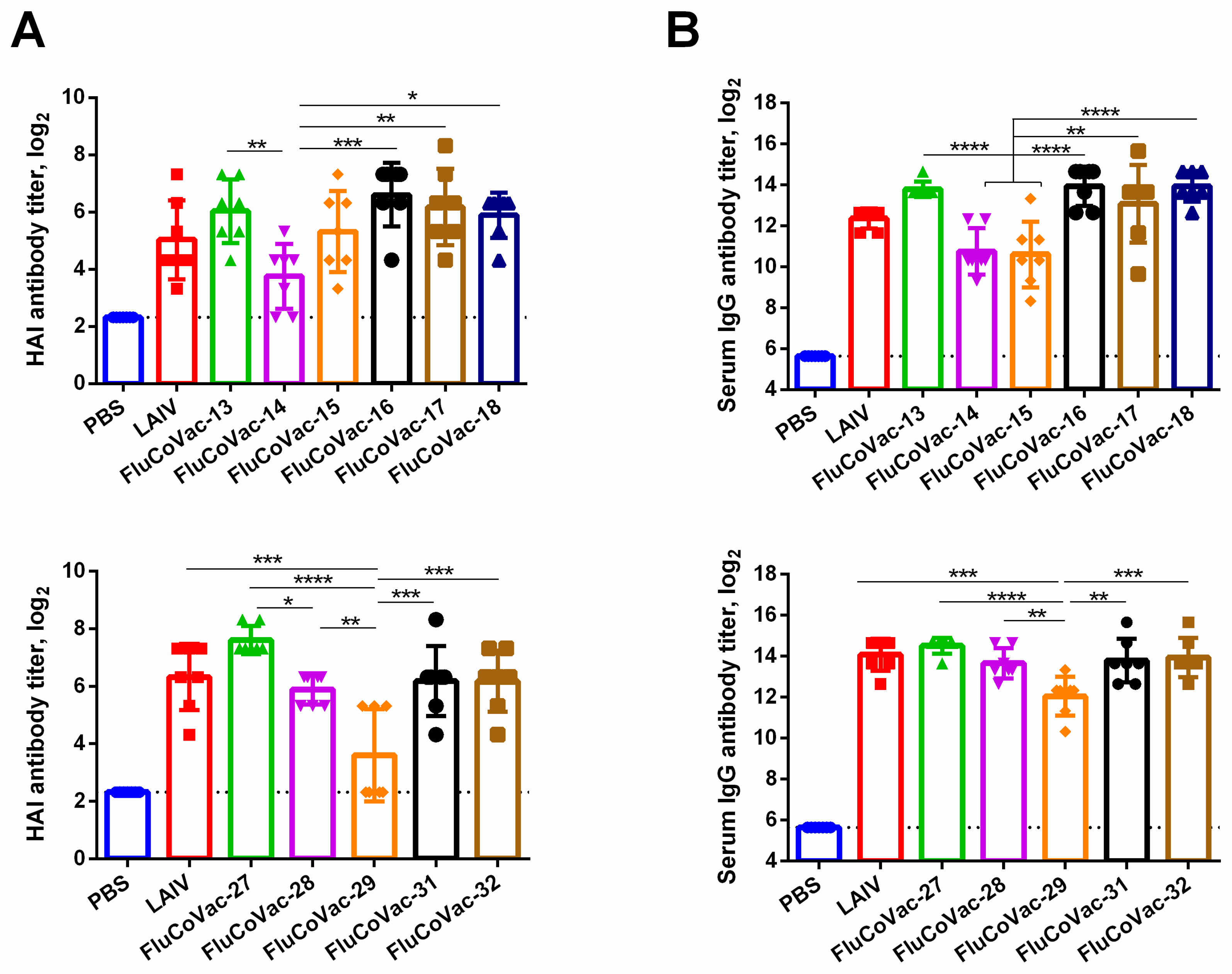
3.3.1. Influenza-Specific Antibody Immune Responses
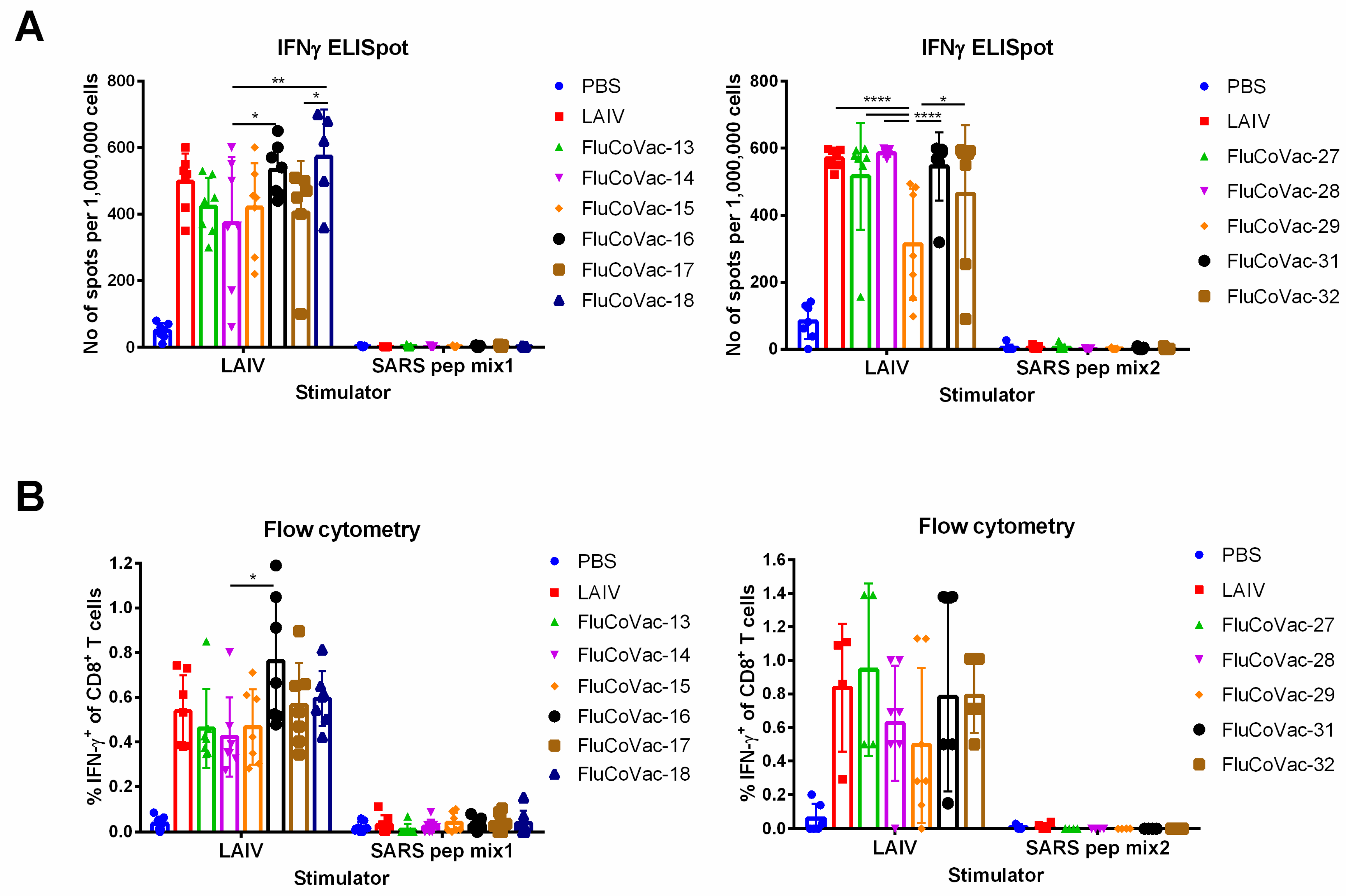
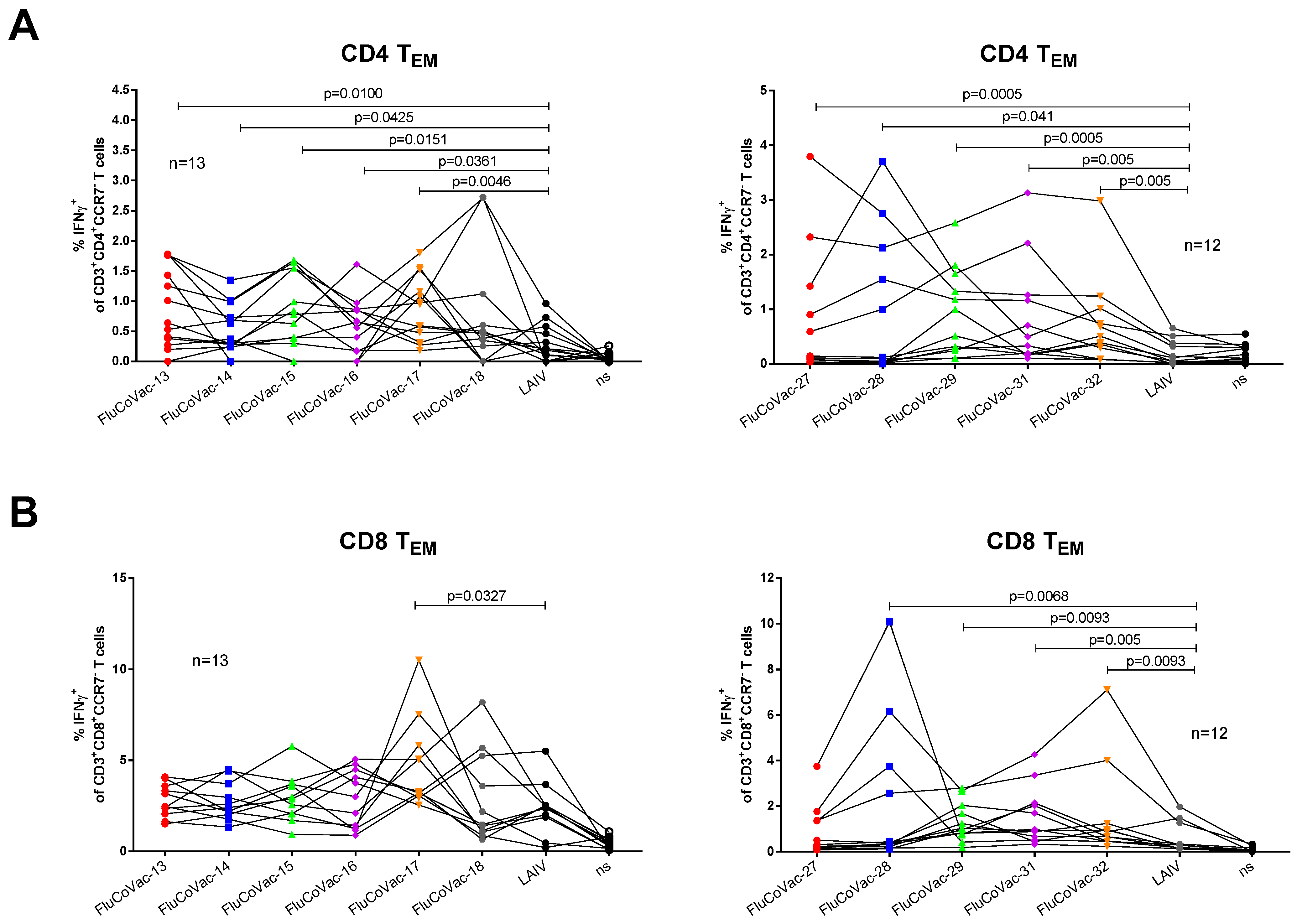
3.3.2. Influenza- and SARS-CoV-2-Specific T-Cell Responses
3.4. Assessment of Recombinant LAIV Viruses in ICS Assay with COVID-19 Convalescents Samples
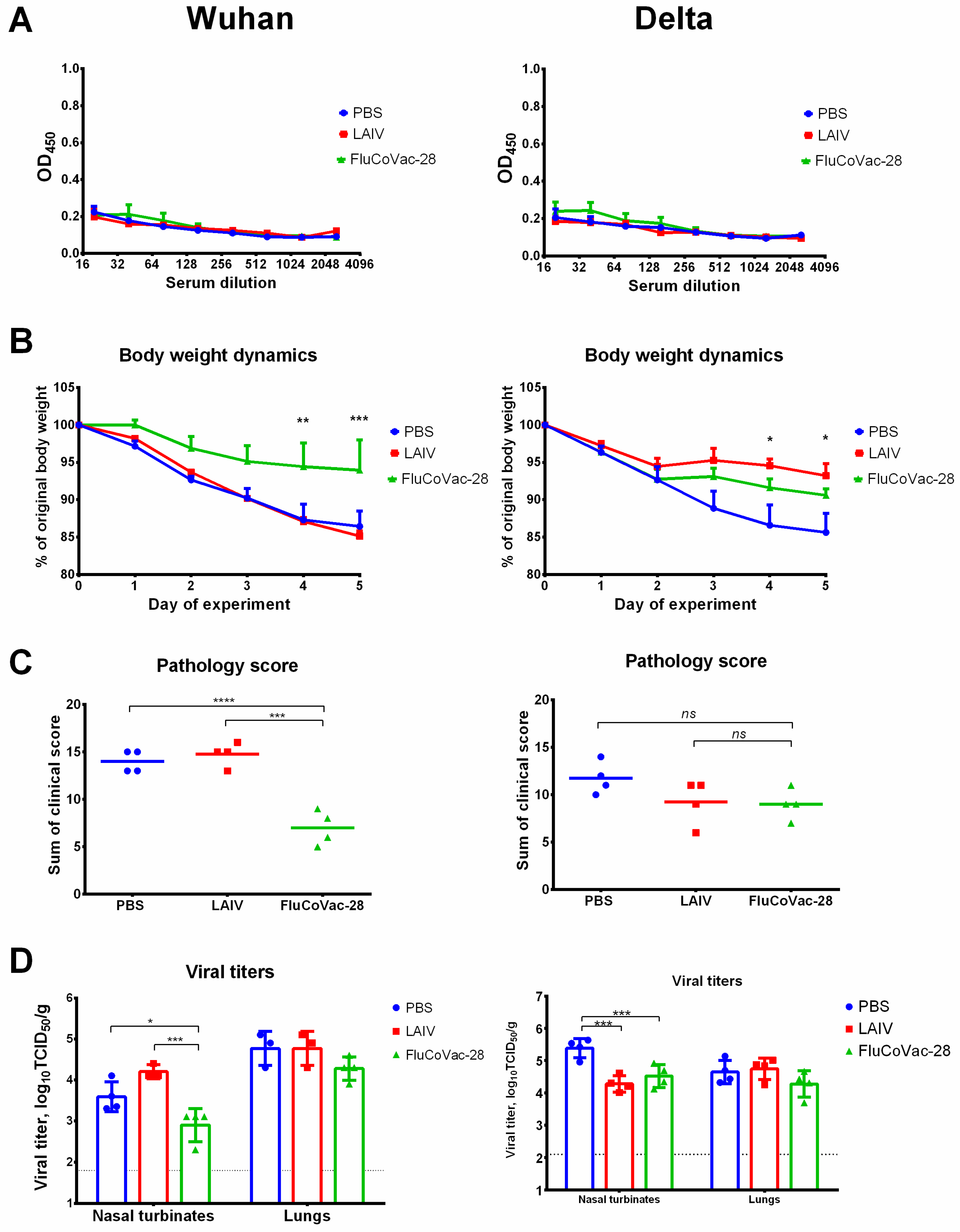
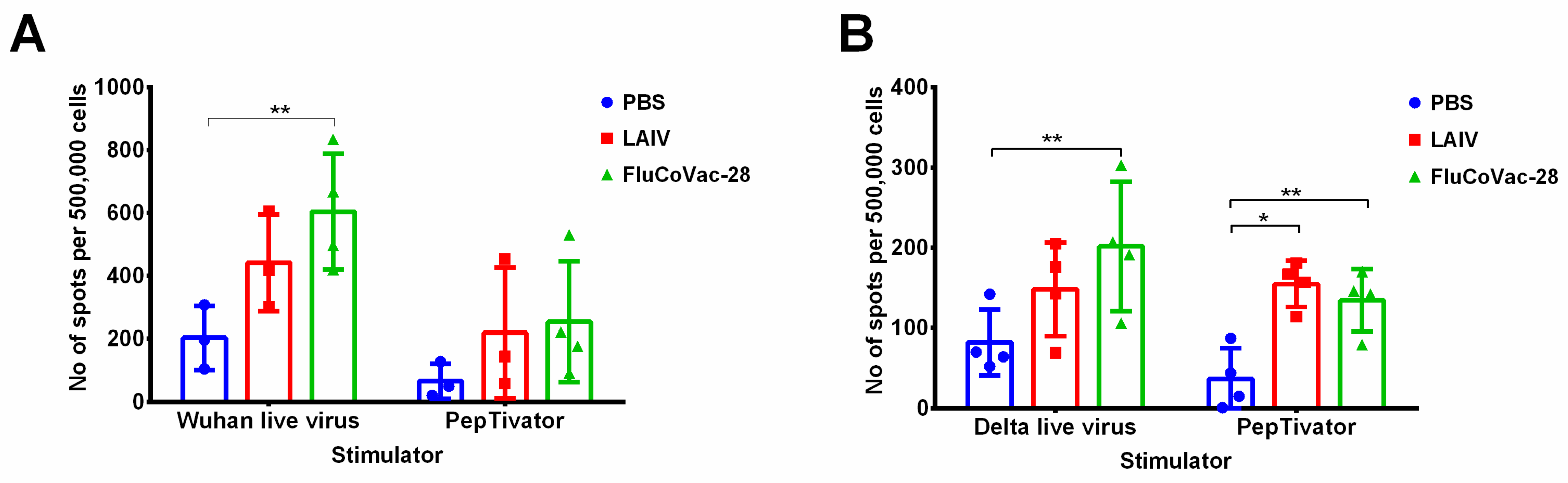
3.5. Immunogenicity and Protective Activity of a Selected LAIV/SARS-CoV-2 Vaccine Prototype in Syrian Hamsters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- COVID-19 Dashboard by the Center for Systems Science and Engineering (CSSE) at Johns Hopkins University (JHU). 2022. Available online: https://coronavirus.jhu.edu/map.html (accessed on 1 May 2022).
- Kyriakidis, N.C.; Lopez-Cortes, A.; Gonzalez, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. NPJ Vaccines 2021, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Sadeghalvad, M.; Mansourabadi, A.H.; Noori, M.; Nejadghaderi, S.A.; Masoomikarimi, M.; Alimohammadi, M.; Rezaei, N. Recent developments in SARS-CoV-2 vaccines: A systematic review of the current studies. Rev. Med. Virol. 2022, e2359. [Google Scholar] [CrossRef]
- Zheng, C.; Shao, W.; Chen, X.; Zhang, B.; Wang, G.; Zhang, W. Real-world effectiveness of COVID-19 vaccines: A literature review and meta-analysis. Int. J. Infect. Dis. 2022, 114, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Wang, S.H.; Tang, Y.; Sheng, W.; Zuo, C.J.; Wu, D.W.; Fang, H.; Du, Q.; Li, N. Landscape and progress of global COVID-19 vaccine development. Hum. Vaccines Immunother. 2021, 17, 3276–3280. [Google Scholar] [CrossRef] [PubMed]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Tatsi, E.B.; Filippatos, F.; Michos, A. SARS-CoV-2 variants and effectiveness of vaccines: A review of current evidence. Epidemiol. Infect. 2021, 149, e237. [Google Scholar] [CrossRef]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef]
- Kedzierska, K.; Thomas, P.G. Count on us: T cells in SARS-CoV-2 infection and vaccination. Cell Rep. Med. 2022, 3, 100562. [Google Scholar] [CrossRef]
- Tarke, A.; Coelho, C.H.; Zhang, Z.; Dan, J.M.; Yu, E.D.; Methot, N.; Bloom, N.I.; Goodwin, B.; Phillips, E.; Mallal, S.; et al. SARS-CoV-2 vaccination induces immunological T cell memory able to cross-recognize variants from Alpha to Omicron. Cell 2022, 185, 847–859.e11. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.R.; Gotz, R.; Mayr, V.; Lohse, M.J.; Holthoff, H.P.; Ungerer, M. SARS-CoV2 wild type and mutant specific humoral and T cell immunity is superior after vaccination than after natural infection. PLoS ONE 2022, 17, e0266701. [Google Scholar] [CrossRef] [PubMed]
- Taus, E.; Hofmann, C.; Ibarrondo, F.J.; Hausner, M.A.; Fulcher, J.A.; Krogstad, P.; Ferbas, K.G.; Tobin, N.H.; Rimoin, A.W.; Aldrovandi, G.M.; et al. Dominant CD8+ T Cell Nucleocapsid Targeting in SARS-CoV-2 Infection and Broad Spike Targeting From Vaccination. Front. Immunol. 2022, 13, 835830. [Google Scholar] [CrossRef] [PubMed]
- de Silva, T.I.; Liu, G.; Lindsey, B.B.; Dong, D.; Moore, S.C.; Hsu, N.S.; Shah, D.; Wellington, D.; Mentzer, A.J.; Angyal, A.; et al. The impact of viral mutations on recognition by SARS-CoV-2 specific T cells. iScience 2021, 24, 103353. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, L.; Schank, M.; Dang, X.; Lu, Z.; Cao, D.; Khanal, S.; Nguyen, L.N.; Nguyen, L.N.T.; Zhang, J.; et al. SARS-CoV-2 specific memory T cell epitopes identified in COVID-19-recovered subjects. Virus Res. 2021, 304, 198508. [Google Scholar] [CrossRef]
- Heitmann, J.S.; Bilich, T.; Tandler, C.; Nelde, A.; Maringer, Y.; Marconato, M.; Reusch, J.; Jager, S.; Denk, M.; Richter, M.; et al. A COVID-19 peptide vaccine for the induction of SARS-CoV-2 T cell immunity. Nature 2022, 601, 617–622. [Google Scholar] [CrossRef]
- Somogyi, E.; Csiszovszki, Z.; Molnar, L.; Lorincz, O.; Toth, J.; Pattijn, S.; Schockaert, J.; Mazy, A.; Miklos, I.; Pantya, K.; et al. A Peptide Vaccine Candidate Tailored to Individuals’ Genetics Mimics the Multi-Targeted T Cell Immunity of COVID-19 Convalescent Subjects. Front. Genet. 2021, 12, 684152. [Google Scholar] [CrossRef]
- Chen, J.; Deng, Y.; Huang, B.; Han, D.; Wang, W.; Huang, M.; Zhai, C.; Zhao, Z.; Yang, R.; Zhao, Y.; et al. DNA Vaccines Expressing the Envelope and Membrane Proteins Provide Partial Protection Against SARS-CoV-2 in Mice. Front. Immunol. 2022, 13, 827605. [Google Scholar] [CrossRef]
- Dangi, T.; Class, J.; Palacio, N.; Richner, J.M.; Penaloza MacMaster, P. Combining spike-and nucleocapsid-based vaccines improves distal control of SARS-CoV-2. Cell Rep. 2021, 36, 109664. [Google Scholar] [CrossRef]
- Matchett, W.E.; Joag, V.; Stolley, J.M.; Shepherd, F.K.; Quarnstrom, C.F.; Mickelson, C.K.; Wijeyesinghe, S.; Soerens, A.G.; Becker, S.; Thiede, J.M.; et al. Cutting Edge: Nucleocapsid Vaccine Elicits Spike-Independent SARS-CoV-2 Protective Immunity. J. Immunol. 2021, 207, 376–379. [Google Scholar] [CrossRef]
- Isakova-Sivak, I.; Matyushenko, V.; Stepanova, E.; Matushkina, A.; Kotomina, T.; Mezhenskaya, D.; Prokopenko, P.; Kudryavtsev, I.; Kopeykin, P.; Sivak, K.; et al. Recombinant Live Attenuated Influenza Vaccine Viruses Carrying Conserved T-Cell Epitopes of Human Adenoviruses Induce Functional Cytotoxic T-Cell Responses and Protect Mice against Both Infections. Vaccines 2020, 8, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotomina, T.; Isakova-Sivak, I.; Matyushenko, V.; Kim, K.H.; Lee, Y.; Jung, Y.J.; Kang, S.M.; Rudenko, L. Recombinant live attenuated influenza vaccine viruses carrying CD8 T-cell epitopes of respiratory syncytial virus protect mice against both pathogens without inflammatory disease. Antivir. Res. 2019, 168, 9–17. [Google Scholar] [CrossRef]
- Kotomina, T.; Korenkov, D.; Matyushenko, V.; Prokopenko, P.; Rudenko, L.; Isakova-Sivak, I. Live attenuated influenza vaccine viral vector induces functional cytotoxic T-cell immune response against foreign CD8+ T-cell epitopes inserted into NA and NS1 genes using the 2A self-cleavage site. Hum. Vaccines Immunother. 2018, 14, 2964–2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matyushenko, V.; Kotomina, T.; Kudryavtsev, I.; Mezhenskaya, D.; Prokopenko, P.; Matushkina, A.; Sivak, K.; Muzhikyan, A.; Rudenko, L.; Isakova-Sivak, I. Conserved T-cell epitopes of respiratory syncytial virus (RSV) delivered by recombinant live attenuated influenza vaccine viruses efficiently induce RSV-specific lung-localized memory T cells and augment influenza-specific resident memory T-cell responses. Antivir. Res. 2020, 182, 104864. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, E.A.; Kotomina, T.S.; Matyushenko, V.A.; Smolonogina, T.A.; Shapovalova, V.S.; Rudenko, L.G.; Isakova-Sivak, I.N. Amino Acid Substitutions N123D and N149D in Hemagglutinin Molecule Enhance Immunigenicity of Live Attenuated Influenza H7N9 Vaccine Strain in Experiment. Bull. Exp. Biol. Med. 2019, 166, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Matyushenko, V.; Isakova-Sivak, I.; Kudryavtsev, I.; Goshina, A.; Chistyakova, A.; Stepanova, E.; Prokopenko, P.; Sychev, I.; Rudenko, L. Detection of IFNγ-Secreting CD4+ and CD8+ Memory T Cells in COVID-19 Convalescents after Stimulation of Peripheral Blood Mononuclear Cells with Live SARS-CoV-2. Viruses 2021, 13, 1490. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, O.; Wall, J.B.J.; Zheng, M.; Zhou, Y.; Wang, L.; Vaseghi, H.R.; Qian, L.; Liu, J. Systematic comparison of 2A peptides for cloning multi-genes in a polycistronic vector. Sci. Rep. 2017, 7, 2193. [Google Scholar] [CrossRef]
- Isakova-Sivak, I.; Chen, L.M.; Matsuoka, Y.; Voeten, J.T.; Kiseleva, I.; Heldens, J.G.; den Bosch, H.; Klimov, A.; Rudenko, L.; Cox, N.J.; et al. Genetic bases of the temperature-sensitive phenotype of a master donor virus used in live attenuated influenza vaccines: A/Leningrad/134/17/57 (H2N2). Virology 2011, 412, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Kozlovskaya, L.; Piniaeva, A.; Ignatyev, G.; Selivanov, A.; Shishova, A.; Kovpak, A.; Gordeychuk, I.; Ivin, Y.; Berestovskaya, A.; Prokhortchouk, E.; et al. Isolation and phylogenetic analysis of SARS-CoV-2 variants collected in Russia during the COVID-19 outbreak. Int. J. Infect. Dis. 2020, 99, 40–46. [Google Scholar] [CrossRef]
- Kittel, B.; Ruehl-Fehlert, C.; Morawietz, G.; Klapwijk, J.; Elwell, M.R.; Lenz, B.; O'Sullivan, M.G.; Roth, D.R.; Wadsworth, P.F.; Group, R.; et al. Revised guides for organ sampling and trimming in rats and mice—Part 2: A joint publication of the RITA and NACAD groups. Exp. Toxicol. Pathol. 2004, 55, 413–431. [Google Scholar] [CrossRef]
- Hsia, C.C.W.; Hyde, D.M.; Ochs, M.; Weibel, E.R. An official research policy statement of the American Thoracic Society/European Respiratory Society: Standards for quantitative assessment of lung structure. Am. J. Respir. Crit. Care Med. 2010, 181, 394–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, T.; Fox, D.; van Doremalen, N.; Ball, E.; Morris, M.K.; Sotomayor-Gonzalez, A.; Servellita, V.; Rustagi, A.; Yinda, C.K.; Fritts, L.; et al. The B.1.427/1.429 (epsilon) SARS-CoV-2 variants are more virulent than ancestral B.1 (614G) in Syrian hamsters. PLoS Pathog. 2022, 18, e1009914. [Google Scholar] [CrossRef] [PubMed]
- Mattoo, S.U.; Myoung, J. T cell responses to SARS-CoV-2 in humans and animals. J. Microbiol. 2022, 60, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Shomuradova, A.S.; Vagida, M.S.; Sheetikov, S.A.; Zornikova, K.V.; Kiryukhin, D.; Titov, A.; Peshkova, I.O.; Khmelevskaya, A.; Dianov, D.V.; Malasheva, M.; et al. SARS-CoV-2 Epitopes Are Recognized by a Public and Diverse Repertoire of Human T Cell Receptors. Immunity 2020, 53, 1245–1257.e5. [Google Scholar] [CrossRef] [PubMed]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Stralin, K.; Gorin, J.B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158–168.e14. [Google Scholar] [CrossRef]
- Tarke, A.; Sidney, J.; Kidd, C.K.; Dan, J.M.; Ramirez, S.I.; Yu, E.D.; Mateus, J.; da Antunes, R.S.; Moore, E.; Rubiro, P.; et al. Comprehensive analysis of T cell immunodominance and immunoprevalence of SARS-CoV-2 epitopes in COVID-19 cases. Cell Reports. Med. 2021, 2, 100204. [Google Scholar] [CrossRef]
- Croxford, J.L.; Ercolini, A.M.; Degutes, M.; Miller, S.D. Structural requirements for initiation of cross-reactivity and CNS autoimmunity with a PLP139-151 mimic peptide derived from murine hepatitis virus. Eur. J. Immunol. 2006, 36, 2671–2680. [Google Scholar] [CrossRef]
- Mathieu, P.A.; Gomez, K.A.; Coutelier, J.P.; Retegui, L.A. Sequence similarity and structural homologies are involved in the autoimmune response elicited by mouse hepatitis virus A59. J. Autoimmun. 2004, 23, 117–126. [Google Scholar] [CrossRef]
- Harndahl, M.; Lamberth, K.; Roder, G.; Justesen, S.; Madsen, M.; Nielsen, M.; Lundegaard, C.; Larsen, M.V.; Tang, S.; Brunak, S.; et al. Large Scale Analysis of Peptide-HLA Class I Interactions (Direct IEDB Submission 1019519). 2010. Available online: http://www.iedb.org/reference/1019519 (accessed on 7 March 2020).
- Sidney, J.; Botten, J.; Neuman, B.; Buchmeier, M.; Sette, A. HLA DRB1*01:01 Binding Capacity of Selected SARS-Derived Peptides. 2006. Available online: http://www.iedb.org/reference/1000399 (accessed on 25 February 2020).
- Sidney, J.; Botten, J.; Neuman, B.; Buchmeier, M.; Sette, A. Large Scale Analysis of Peptide-HLA Class I Interactions (Direct IEDB Submissions 1000402, 1000425). 2006. Available online: http://www.iedb.org/reference/1000425 (accessed on 25 February 2020).
- Rasmussen, M.; Harndahl, M.N.; Nielsen, L.L.; Stryhn, A.; Nielsen, M.; Buus, S. Describing the Peptide Binding Specificity of HLA-C Molecules (Direct IEDB Submissions 1028228, 1028231, 1028232, 1028233, 1028234, 1028235, 1028286). 2014. Available online: http://www.iedb.org/reference/1028232 (accessed on 23 April 2020).
- Rasmussen, M.; Harndahl, M.N.; Kristensen, A.B.; Nielsen, I.K.; Jorgensen, K.W.; Stryhn, A.; Nielsen, M.; Buus, S. Large Scale Analysis of Peptide—HLA-I Stability (Direct IEDB Submissions 1028282, 1028285, 1028287, 1028288, 1028289, 1028290, 1028291, 1028292, 1028293). 2014. Available online: http://www.iedb.org/reference/1028282 (accessed on 23 April 2020).
- Lee, E.; Sandgren, K.; Duette, G.; Stylianou, V.V.; Khanna, R.; Eden, J.S.; Blyth, E.; Gottlieb, D.; Cunningham, A.L.; Palmer, S. Identification of SARS-CoV-2 Nucleocapsid and Spike T-Cell Epitopes for Assessing T-Cell Immunity. J. Virol. 2021, 95, e02002-20. [Google Scholar] [CrossRef]
- Zhou, B.; Li, Y.; Belser, J.A.; Pearce, M.B.; Schmolke, M.; Subba, A.X.; Shi, Z.; Zaki, S.R.; Blau, D.M.; Garcia-Sastre, A.; et al. NS-based live attenuated H1N1 pandemic vaccines protect mice and ferrets. Vaccine 2010, 28, 8015–8025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferko, B.; Stasakova, J.; Sereinig, S.; Romanova, J.; Katinger, D.; Niebler, B.; Katinger, H.; Egorov, A. Hyperattenuated recombinant influenza A virus nonstructural-protein-encoding vectors induce human immunodeficiency virus type 1 Nef-specific systemic and mucosal immune responses in mice. J. Virol. 2001, 75, 8899–8908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Guan, J.; Handel, A.; Tscharke, D.C.; Sidney, J.; Sette, A.; Wakim, L.M.; Sng, X.Y.X.; Thomas, P.G.; Croft, N.P.; et al. Quantification of epitope abundance reveals the effect of direct and cross-presentation on influenza CTL responses. Nat. Commun. 2019, 10, 2846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kedzierska, K.; Guillonneau, C.; Gras, S.; Hatton, L.A.; Webby, R.; Purcell, A.W.; Rossjohn, J.; Doherty, P.C.; Turner, S.J. Complete modification of TCR specificity and repertoire selection does not perturb a CD8+ T cell immunodominance hierarchy. Proc. Natl. Acad. Sci. USA 2008, 105, 19408–19413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohandas, S.; Yadav, P.D.; Shete, A.; Nyayanit, D.; Sapkal, G.; Lole, K.; Gupta, N. SARS-CoV-2 Delta Variant Pathogenesis and Host Response in Syrian Hamsters. Viruses 2021, 13, 1773. [Google Scholar] [CrossRef]
- Nathan, A.; Rossin, E.J.; Kaseke, C.; Park, R.J.; Khatri, A.; Koundakjian, D.; Urbach, J.M.; Singh, N.K.; Bashirova, A.; Tano-Menka, R.; et al. Structure-guided T cell vaccine design for SARS-CoV-2 variants and sarbecoviruses. Cell 2021, 184, 4401–4413.e10. [Google Scholar] [CrossRef]
- Altmann, D.M.; Boyton, R.J. SARS-CoV-2 T cell immunity: Specificity, function, durability, and role in protection. Sci. Immunol. 2020, 5, eabd6160. [Google Scholar] [CrossRef]
- Ng, O.-W.; Chia, A.; Tan, A.T.; Jadi, R.S.; Leong, H.N.; Bertoletti, A.; Tan, Y.J. Memory T cell responses targeting the SARS coronavirus persist up to 11 years post-infection. Vaccine 2016, 34, 2008–2014. [Google Scholar] [CrossRef]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef]
- Loyal, L.; Braun, J.; Henze, L.; Kruse, B.; Dingeldey, M.; Reimer, U.; Kern, F.; Schwarz, T.; Mangold, M.; Unger, C.; et al. Cross-reactive CD4+ T cells enhance SARS-CoV-2 immune responses upon infection and vaccination. Science 2021, 374, eabh1823. [Google Scholar] [CrossRef]
- Low, J.S.; Vaqueirinho, D.; Mele, F.; Foglierini, M.; Jerak, J.; Perotti, M.; Jarrossay, D.; Jovic, S.; Perez, L.; Cacciatore, R.; et al. Clonal analysis of immunodominance and cross-reactivity of the CD4 T cell response to SARS-CoV-2. Science 2021, 372, 1336–1341. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S.; Srivastava, R.; Coulon, P.G.; Dhanushkodi, N.R.; Chentoufi, A.A.; Tifrea, D.F.; Edwards, R.A.; Figueroa, C.J.; Schubl, S.D.; Hsieh, L.; et al. Genome-Wide B Cell, CD4+, and CD8+ T Cell Epitopes That Are Highly Conserved between Human and Animal Coronaviruses, Identified from SARS-CoV-2 as Targets for Preemptive Pan-Coronavirus Vaccines. J. Immunol. 2021, 206, 2566–2582. [Google Scholar] [CrossRef] [PubMed]
- Knierman, M.D.; Lannan, M.B.; Spindler, L.J.; McMillian, C.L.; Konrad, R.J.; Siegel, R.W. The Human Leukocyte Antigen Class II Immunopeptidome of the SARS-CoV-2 Spike Glycoprotein. Cell Rep. 2020, 33, 108454. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; He, M.; Wang, X.; Sun, Q.; Kuang, M. Specific CD8+ TCR Repertoire Recognizing Conserved Antigens of SARS-CoV-2 in Unexposed Population: A Prerequisite for Broad-Spectrum CD8+ T Cell Immunity. Vaccines 2021, 9, 1093. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Shen, M.; Han, X.; Chen, Q.; Li, L.; Chen, S.; Zhang, J.; Gao, F.; Wang, W.; Wang, Y.; et al. Identification of cross-reactive CD8+ T cell receptors with high functional avidity to a SARS-CoV-2 immunodominant epitope and its natural mutant variants. Genes Dis. 2022, 9, 216–229. [Google Scholar] [CrossRef]
- Eisenlohr, L.C.; Yewdell, J.W.; Bennink, J.R. Flanking sequences influence the presentation of an endogenously synthesized peptide to cytotoxic T lymphocytes. J. Exp. Med. 1992, 175, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Steers, N.J.; Currier, J.R.; Jobe, O.; Tovanabutra, S.; Ratto-Kim, S.; Marovich, M.A.; Kim, J.H.; Michael, N.L.; Alving, C.R.; Rao, M. Designing the epitope flanking regions for optimal generation of CTL epitopes. Vaccine 2014, 32, 3509–3516. [Google Scholar] [CrossRef]
- Korenkov, D.; Isakova-Sivak, I.; Rudenko, L. Basics of CD8 T-cell immune responses after influenza infection and vaccination with inactivated or live attenuated influenza vaccine. Expert Rev. Vaccines 2018, 17, 977–987. [Google Scholar] [CrossRef]
- Isakova-Sivak, I.; Grigorieva, E.; Rudenko, L. Insights into current clinical research on the immunogenicity of live attenuated influenza vaccines. Expert Rev. Vaccines 2020, 19, 43–55. [Google Scholar] [CrossRef]
- Rudenko, L.; Isakova-Sivak, I.; Naykhin, A.; Kiseleva, I.; Stukova, M.; Erofeeva, M.; Korenkov, D.; Matyushenko, V.; Sparrow, E.; Kieny, M.P. H7N9 live attenuated influenza vaccine in healthy adults: A randomised, double-blind, placebo-controlled, phase 1 trial. Lancet. Infect. Dis. 2016, 16, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Vasilyev, K.; Shurygina, A.P.; Sergeeva, M.; Stukova, M.; Egorov, A. Intranasal Immunization with the Influenza A Virus Encoding Truncated NS1 Protein Protects Mice from Heterologous Challenge by Restraining the Inflammatory Response in the Lungs. Microorganisms 2021, 9, 690. [Google Scholar] [CrossRef] [PubMed]
- Pica, N.; Langlois, R.A.; Krammer, F.; Margine, I.; Palese, P. NS1-truncated live attenuated virus vaccine provides robust protection to aged mice from viral challenge. J. Virol. 2012, 86, 10293–10301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isakova-Sivak, I.; Stepanova, E.; Mezhenskaya, D.; Matyushenko, V.; Prokopenko, P.; Sychev, I.; Wong, P.F.; Rudenko, L. Influenza vaccine: Progress in a vaccine that elicits a broad immune response. Expert Rev. Vaccines 2021, 20, 1097–1112. [Google Scholar] [CrossRef] [PubMed]
- Kotturi, M.F.; Assarsson, E.; Peters, B.; Grey, H.; Oseroff, C.; Pasquetto, V.; Sette, A. Of mice and humans: How good are HLA transgenic mice as a model of human immune responses? Immunome Res. 2009, 5, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, T.M.; Gittelman, R.M.; Klinger, M.; May, D.H.; Osborne, E.J.; Taniguchi, R.; Zahid, H.J.; Kaplan, I.M.; Dines, J.N.; Noakes, M.T.; et al. Magnitude and Dynamics of the T-Cell Response to SARS-CoV-2 Infection at Both Individual and Population Levels. medRxiv 2020. [Google Scholar] [CrossRef]
- Jin, X.; Ding, Y.; Sun, S.; Wang, X.; Zhou, Z.; Liu, X.; Li, M.; Chen, X.; Shen, A.; Wu, Y.; et al. Screening HLA-A-restricted T cell epitopes of SARS-CoV-2 and the induction of CD8+ T cell responses in HLA-A transgenic mice. Cell. Mol. Immunol. 2021, 18, 2588–2608. [Google Scholar] [CrossRef]
- Deng, J.; Pan, J.; Qiu, M.; Mao, L.; Wang, Z.; Zhu, G.; Gao, L.; Su, J.; Hu, Y.; Luo, O.J.; et al. Identification of HLA-A2 restricted CD8+ T cell epitopes in SARS-CoV-2 structural proteins. J. Leukoc. Biol. 2021, 110, 1171–1180. [Google Scholar] [CrossRef]
- Johansson, A.M.; Malhotra, U.; Kim, Y.G.; Gomez, R.; Krist, M.P.; Wald, A.; Koelle, D.M.; Kwok, W.W. Cross-reactive and mono-reactive SARS-CoV-2 CD4+ T cells in prepandemic and COVID-19 convalescent individuals. PLoS Pathog. 2021, 17, e1010203. [Google Scholar] [CrossRef]
- Kingstad-Bakke, B.; Lee, W.; Chandrasekar, S.S.; Gasper, D.J.; Salas-Quinchucua, C.; Cleven, T.; Sullivan, J.A.; Talaat, A.; Osorio, J.E.; Suresh, M. Vaccine-induced systemic and mucosal T cell immunity to SARS-CoV-2 viral variants. Proc. Natl. Acad. Sci. USA 2022, 119, e2118312119. [Google Scholar] [CrossRef]
- Mahajan, S.; Kode, V.; Bhojak, K.; Karunakaran, C.; Lee, K.; Manoharan, M.; Ramesh, A.; Hv, S.; Srivastava, A.; Sathian, R.; et al. Immunodominant T-cell epitopes from the SARS-CoV-2 spike antigen reveal robust pre-existing T-cell immunity in unexposed individuals. Sci. Rep. 2021, 11, 13164. [Google Scholar] [CrossRef]
- Pallikkuth, S.; Williams, E.; Pahwa, R.; Hoffer, M.; Pahwa, S. Association of Flu specific and SARS-CoV-2 specific CD4 T cell responses in SARS-CoV-2 infected asymptomatic heath care workers. Vaccine 2021, 39, 6019–6024. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, P.; Yuan, L.; Zhang, L.; Zhang, L.; Zhao, H.; Chen, C.; Wang, X.; Han, J.; Chen, Y.; et al. A live attenuated virus-based intranasal COVID-19 vaccine provides rapid, prolonged, and broad protection against SARS-CoV-2. Sci. Bull. 2022, 67, 1372–1387. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recombinant Influenza Virus ID | SARS-CoV-2 T Cassette | Cassette Size, aa | Influenza Gene Modified | Mean Viral Titer, log10EID(TCID)50/mL | |||
|---|---|---|---|---|---|---|---|
| Eggs | MDCK Cells | Nasal Turbinates † | Lungs † | ||||
| H7N9 LAIV | - | - | - | 9.8 | 8.2 | 3.2 | 1.6 |
| FluCoVac-13 | Cas #1 | 96 | NS | 7.3 * | 4.1 * | 1.6 * | ≤1.2 |
| FluCoVac-14 | Cas #2 | 96 | NS | 6.8 * | 4.2 * | ≤1.2 * | ≤1.2 |
| FluCoVac-15 | Cas #3 | 122 | NS | 8.0 * | 4.1 * | ≤1.2 * | ≤1.2 |
| FluCoVac-16 | Cas #4 | 105 | NA | 8.3 * | 5.4 * | 2.9 | ≤1.2 |
| FluCoVac-17 | Cas #5 | 122 | NA | 8.1 * | 5.7 * | 3.7 | ≤1.2 |
| FluCoVac-18 | Cas #6 | 101 | NA | 8.4 * | 6.8 * | 3.3 | ≤1.2 |
| FluCoVac-27 | Cas #7 | 161 | NA | 8.3 * | 6.6 * | 3.6 | 1.3 |
| FluCoVac-28 | Cas #8 | 174 | NA | 9.7 | 8.3 | 3.8 | 1.5 |
| FluCoVac-29 | Cas #9 | 122 | NS | 7.5 * | 4.1 * | ≤1.2 * | ≤1.2 |
| FluCoVac-31 | Cas #10 | 147 | NS | 7.7 * | 4.3 * | ≤1.2 * | ≤1.2 |
| FluCoVac-32 | Cas #7 | 161 | NS | 8.2 * | 4.1 * | ≤1.2 * | ≤1.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isakova-Sivak, I.; Stepanova, E.; Matyushenko, V.; Niskanen, S.; Mezhenskaya, D.; Bazhenova, E.; Krutikova, E.; Kotomina, T.; Prokopenko, P.; Neterebskii, B.; et al. Development of a T Cell-Based COVID-19 Vaccine Using a Live Attenuated Influenza Vaccine Viral Vector. Vaccines 2022, 10, 1142. https://doi.org/10.3390/vaccines10071142
Isakova-Sivak I, Stepanova E, Matyushenko V, Niskanen S, Mezhenskaya D, Bazhenova E, Krutikova E, Kotomina T, Prokopenko P, Neterebskii B, et al. Development of a T Cell-Based COVID-19 Vaccine Using a Live Attenuated Influenza Vaccine Viral Vector. Vaccines. 2022; 10(7):1142. https://doi.org/10.3390/vaccines10071142
Chicago/Turabian StyleIsakova-Sivak, Irina, Ekaterina Stepanova, Victoria Matyushenko, Sergei Niskanen, Daria Mezhenskaya, Ekaterina Bazhenova, Elena Krutikova, Tatiana Kotomina, Polina Prokopenko, Bogdan Neterebskii, and et al. 2022. "Development of a T Cell-Based COVID-19 Vaccine Using a Live Attenuated Influenza Vaccine Viral Vector" Vaccines 10, no. 7: 1142. https://doi.org/10.3390/vaccines10071142