
Humoral Responses against BQ.1.1 Elicited after Breakthrough Infection and SARS-CoV-2 mRNA Vaccination
 , , , , , ,
, , , , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Human Subjects
2.3. Plasma Samples and Antibodies
2.4. Plasmids
2.5. Cell Lines
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Cell Surface Staining and Flow Cytometry Analysis
2.8. Virus Neutralization Assay
2.9. Statistical Analysis
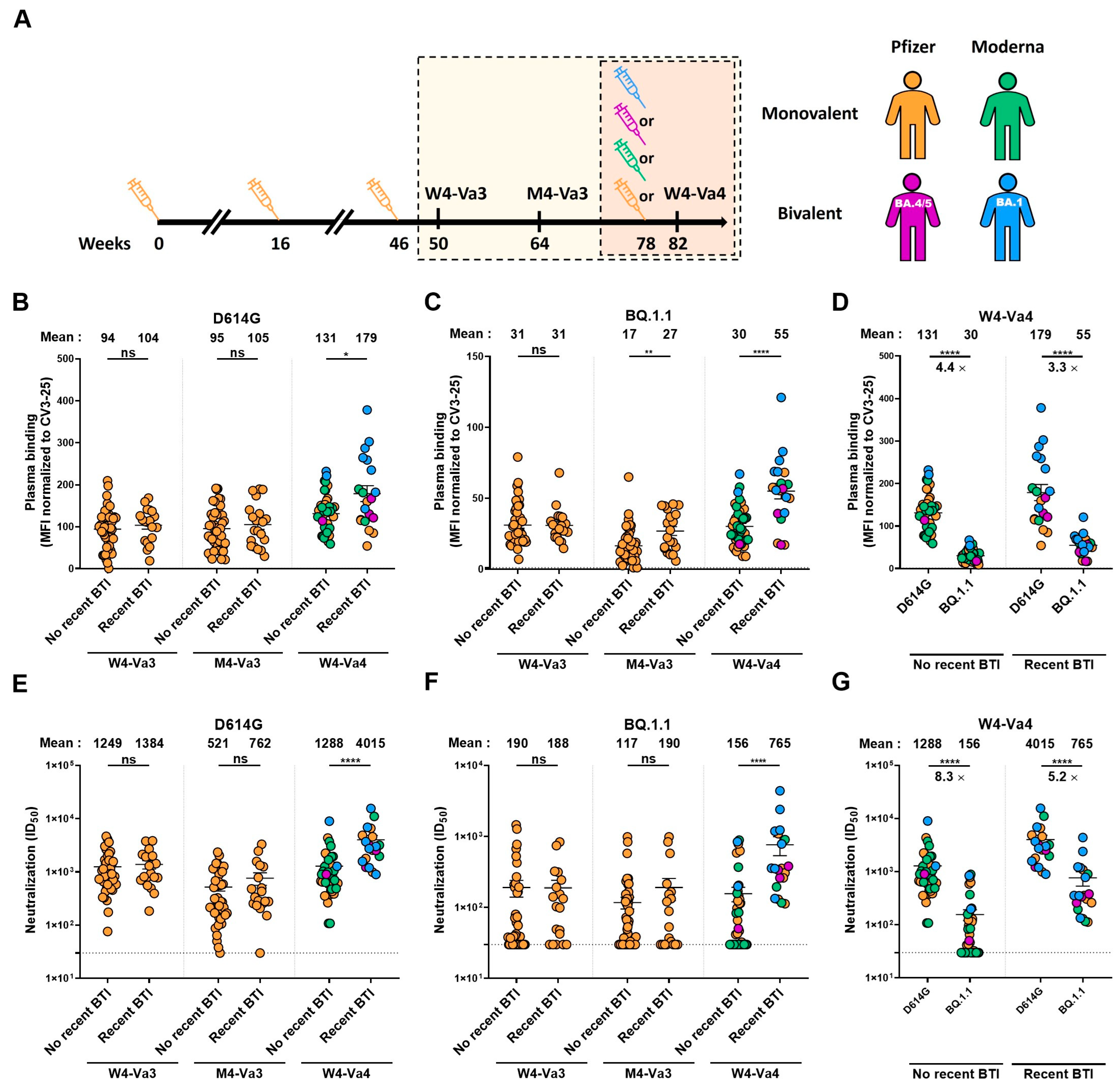
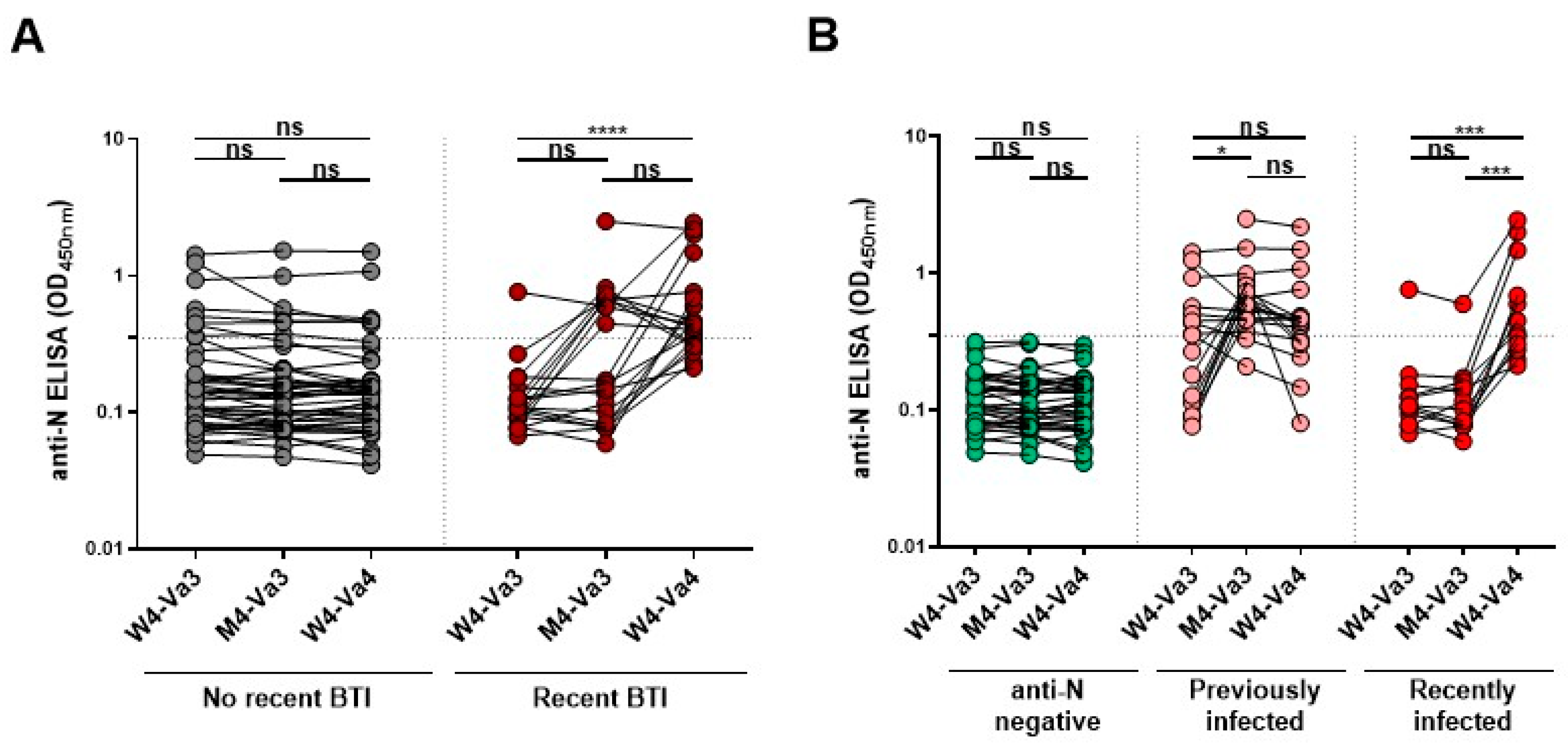
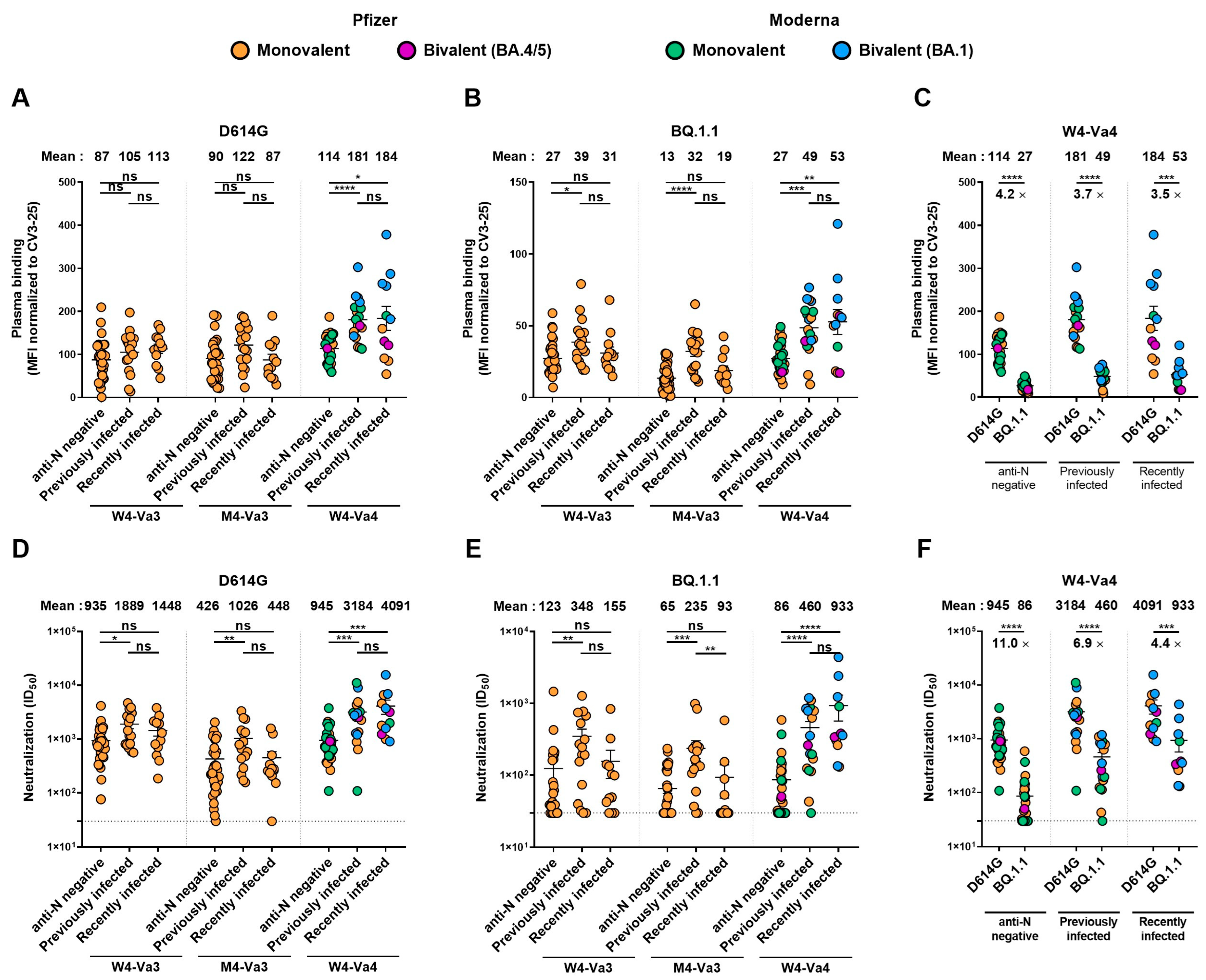
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Données sur les Variants du SRAS-CoV-2 au Québec. Available online: https://www.inspq.qc.ca/covid-19/donnees/variants (accessed on 6 December 2022).
- CDC. COVID Data Tracker Weekly Review. Available online: https://www.cdc.gov/coronavirus/2019-ncov/covid-data/covidview/index.html (accessed on 6 December 2022).
- Qu, P.; Evans, J.P.; Faraone, J.; Zheng, Y.-M.; Carlin, C.; Anghelina, M.; Stevens, P.; Fernandez, S.; Jones, D.; Lozanski, G.; et al. Enhanced Neutralization Resistance of SARS-CoV-2 Omicron Subvariants BQ.1, BQ.1.1, BA.4.6, BF.7 and BA.2.75.2. Cell Host Microbe 2022, 31, 9–17.e3. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.; Nicolas, A.; Ding, S.; Benlarbi, M.; Medjahed, H.; Chatterjee, D.; Dionne, K.; Gong, S.Y.; Gendron-Lepage, G.; Bo, Y.; et al. SARS-CoV-2 Omicron Subvariants Spike Recognition and Neutralization Elicited after the Third Dose of MRNA Vaccine. Cell Rep. 2023, 42, 111998. [Google Scholar] [CrossRef] [PubMed]
- Kurhade, C.; Zou, J.; Xia, H.; Liu, M.; Chang, H.C.; Ren, P.; Xie, X.; Shi, P.-Y. Low Neutralization of SARS-CoV-2 Omicron BA.2.75.2, BQ.1.1, and XBB.1 by Parental MRNA Vaccine or a BA.5-Bivalent Booster. Nat. Med. 2022. [Google Scholar] [CrossRef] [PubMed]
- FDA. Coronavirus (COVID-19) Update: FDA Authorizes Moderna, Pfizer-BioNTech Bivalent COVID-19 Vaccines for Use as a Booster Dose. Available online: https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-moderna-pfizer-biontech-bivalent-covid-19-vaccines-use (accessed on 6 December 2022).
- EMA. First Adapted COVID-19 Booster Vaccines Recommended for Approval in the EU. Available online: https://www.ema.europa.eu/en/news/first-adapted-covid-19-booster-vaccines-recommended-approval-eu (accessed on 6 December 2022).
- Health Canada. Health Canada Authorizes COVID-19 Vaccine Booster Targeting the Omicron BA.4/BA.5 Subvariants. Available online: https://www.canada.ca/en/health-canada/news/2022/10/health-canada-authorizes-covid-19-vaccine-booster-targeting-the-omicron-ba4ba5-subvariants.html (accessed on 6 December 2022).
- Pfizer and BioNTech Complete Submission to European Medicines Agency for Omicron BA.4/BA.5 Adapted Bivalent Vaccine|Pfizer. Available online: https://www.pfizer.com/news/press-release/press-release-detail/pfizer-and-biontech-complete-submission-european-medicines (accessed on 6 December 2022).
- Canada|SPIKEVAXTM Information. Available online: https://www.modernacovid19global.com/ca/ (accessed on 6 December 2022).
- Ntziora, F.; Kostaki, E.G.; Karapanou, A.; Mylona, M.; Tseti, I.; Sipsas, N.V.; Paraskevis, D.; Sfikakis, P.P. Protection of Vaccination versus Hybrid Immunity against Infection with COVID-19 Omicron Variants among Health-Care Workers. Vaccine 2022, 40, 7195–7200. [Google Scholar] [CrossRef] [PubMed]
- Altarawneh, H.N.; Chemaitelly, H.; Ayoub, H.H.; Tang, P.; Hasan, M.R.; Yassine, H.M.; Al-Khatib, H.A.; Smatti, M.K.; Coyle, P.; Al-Kanaani, Z.; et al. Effects of Previous Infection and Vaccination on Symptomatic Omicron Infections. N. Engl. J. Med. 2022, 387, 21–34. [Google Scholar] [CrossRef]
- Carazo, S.; Skowronski, D.M.; Brisson, M.; Barkati, S.; Sauvageau, C.; Brousseau, N.; Gilca, R.; Fafard, J.; Talbot, D.; Ouakki, M.; et al. Protection against Omicron (B.1.1.529) BA.2 Reinfection Conferred by Primary Omicron BA.1 or Pre-Omicron SARS-CoV-2 Infection among Health-Care Workers with and without MRNA Vaccination: A Test-Negative Case-Control Study. Lancet Infect. Dis. 2022, 23, 45–55. [Google Scholar] [CrossRef]
- Tauzin, A.; Nayrac, M.; Benlarbi, M.; Gong, S.Y.; Gasser, R.; Beaudoin-Bussières, G.; Brassard, N.; Laumaea, A.; Vézina, D.; Prévost, J.; et al. A Single Dose of the SARS-CoV-2 Vaccine BNT162b2 Elicits Fc-Mediated Antibody Effector Functions and T Cell Responses. Cell Host Microbe 2021, 29, 1137–1150.e6. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.; Gong, S.Y.; Beaudoin-Bussières, G.; Vézina, D.; Gasser, R.; Nault, L.; Marchitto, L.; Benlarbi, M.; Chatterjee, D.; Nayrac, M.; et al. Strong Humoral Immune Responses against SARS-CoV-2 Spike after BNT162b2 MRNA Vaccination with a 16-Week Interval between Doses. Cell Host Microbe 2022, 30, 97–109.e5. [Google Scholar] [CrossRef]
- Stamatatos, L.; Czartoski, J.; Wan, Y.-H.; Homad, L.J.; Rubin, V.; Glantz, H.; Neradilek, M.; Seydoux, E.; Jennewein, M.F.; MacCamy, A.J.; et al. MRNA Vaccination Boosts Cross-Variant Neutralizing Antibodies Elicited by SARS-CoV-2 Infection. Science 2021, 372, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.; Gong, S.Y.; Chatterjee, D.; Ding, S.; Painter, M.M.; Goel, R.R.; Beaudoin-Bussières, G.; Marchitto, L.; Boutin, M.; Laumaea, A.; et al. A Boost with SARS-CoV-2 BNT162b2 MRNA Vaccine Elicits Strong Humoral Responses Independently of the Interval between the First Two Doses. Cell Rep. 2022, 41, 111554. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.T.; Kwan, A.T.; Rodríguez-Barraquer, I.; Singer, B.J.; Park, H.J.; Lewnard, J.A.; Sears, D.; Lo, N.C. Infectiousness of SARS-CoV-2 Breakthrough Infections and Reinfections during the Omicron Wave. Nat. Med. 2023, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Woodbridge, Y.; Amit, S.; Huppert, A.; Kopelman, N.M. Viral Load Dynamics of SARS-CoV-2 Delta and Omicron Variants Following Multiple Vaccine Doses and Previous Infection. Nat. Commun. 2022, 13, 6706. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Tempia, S.; Kleynhans, J.; von Gottberg, A.; McMorrow, M.L.; Wolter, N.; Bhiman, J.N.; Moyes, J.; du Plessis, M.; Carrim, M.; et al. SARS-CoV-2 Transmission, Persistence of Immunity, and Estimates of Omicron’s Impact in South African Population Cohorts. Sci. Transl. Med. 2022, 14, eabo7081. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.S.V.; Gribben, C.; Bishop, J.; Hanlon, P.; Caldwell, D.; Wood, R.; Reid, M.; McMenamin, J.; Goldberg, D.; Stockton, D.; et al. Effect of Vaccination on Transmission of SARS-CoV-2. N. Engl. J. Med. 2021, 385, 1718–1720. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Hall, J.A.; Zaidi, A.; Andrews, N.J.; Dunbar, J.K.; Dabrera, G. Effect of Vaccination on Household Transmission of SARS-CoV-2 in England. N. Engl. J. Med. 2021, 385, 759–760. [Google Scholar] [CrossRef]
- COVID-19 Immunity Task Force. Seroprevalence in Canada. Available online: https://www.Covid19immunitytaskforce.ca/Seroprevalence-in-Canada/.COVID-19 Immunity Task Force (accessed on 30 November 2022).
- Bazin, R.; Rochette, S.; Perreault, J.; Fournier, M.-J.; Grégoire, Y.; Boivin, A.; Lewin, A.; Germain, M.; Renaud, C. Development and Use of a Method Based on the Anti-N Reactivity of Longitudinal Samples to Better Estimate SARS-CoV-2 Seroprevalence in a Vaccinated Population. medRxiv 2022. [Google Scholar] [CrossRef]
- Germain, M.; Lewin, A.; Bazin, R.; Dieudé, M.; Perreault, J.; Boivin, A.; Grégoire, Y.; Renaud, C. Cohort Profile: A Québec-Based Plasma Donor Biobank to Study COVID-19 Immunity (PlasCoV). medRxiv 2022. [Google Scholar] [CrossRef]
- Prévost, J.; Richard, J.; Gasser, R.; Ding, S.; Fage, C.; Anand, S.P.; Adam, D.; Vergara, N.G.; Tauzin, A.; Benlarbi, M.; et al. Impact of Temperature on the Affinity of SARS-CoV-2 Spike Glycoprotein for Host ACE2. J. Biol. Chem. 2021, 297, 101151. [Google Scholar] [CrossRef]
- Jennewein, M.F.; MacCamy, A.J.; Akins, N.R.; Feng, J.; Homad, L.J.; Hurlburt, N.K.; Seydoux, E.; Wan, Y.-H.; Stuart, A.B.; Edara, V.V.; et al. Isolation and Characterization of Cross-Neutralizing Coronavirus Antibodies from COVID-19+ Subjects. Cell Rep. 2021, 36, 109353. [Google Scholar] [CrossRef]
- Gong, S.Y.; Chatterjee, D.; Richard, J.; Prévost, J.; Tauzin, A.; Gasser, R.; Bo, Y.; Vézina, D.; Goyette, G.; Gendron-Lepage, G.; et al. Contribution of Single Mutations to Selected SARS-CoV-2 Emerging Variants Spike Antigenicity. Virology 2021, 563, 134–145. [Google Scholar] [CrossRef]
- Chatterjee, D.; Tauzin, A.; Marchitto, L.; Gong, S.Y.; Boutin, M.; Bourassa, C.; Beaudoin-Bussières, G.; Bo, Y.; Ding, S.; Laumaea, A.; et al. SARS-CoV-2 Omicron Spike Recognition by Plasma from Individuals Receiving BNT162b2 MRNA Vaccination with a 16-Week Interval between Doses. Cell Rep. 2022, 38, 110429. [Google Scholar] [CrossRef] [PubMed]
- Prévost, J.; Gasser, R.; Beaudoin-Bussières, G.; Richard, J.; Duerr, R.; Laumaea, A.; Anand, S.P.; Goyette, G.; Benlarbi, M.; Ding, S.; et al. Cross-Sectional Evaluation of Humoral Responses against SARS-CoV-2 Spike. Cell Rep. Med. 2020, 1, 100126. [Google Scholar] [CrossRef]
- Perreault, J.; Tremblay, T.; Fournier, M.-J.; Drouin, M.; Beaudoin-Bussières, G.; Prévost, J.; Lewin, A.; Bégin, P.; Finzi, A.; Bazin, R. Waning of SARS-CoV-2 RBD Antibodies in Longitudinal Convalescent Plasma Samples within 4 Months after Symptom Onset. Blood 2020, 136, 2588–2591. [Google Scholar] [CrossRef] [PubMed]
- Qu, P.; Faraone, J.; Evans, J.P.; Zou, X.; Zheng, Y.-M.; Carlin, C.; Bednash, J.S.; Lozanski, G.; Mallampalli, R.K.; Saif, L.J.; et al. Neutralization of the SARS-CoV-2 Omicron BA.4/5 and BA.2.12.1 Subvariants. N. Engl. J. Med. 2022, 386, 2526–2528. [Google Scholar] [CrossRef] [PubMed]
- Tuekprakhon, A.; Nutalai, R.; Dijokaite-Guraliuc, A.; Zhou, D.; Ginn, H.M.; Selvaraj, M.; Liu, C.; Mentzer, A.J.; Supasa, P.; Duyvesteyn, H.M.E.; et al. Antibody Escape of SARS-CoV-2 Omicron BA.4 and BA.5 from Vaccine and BA.1 Serum. Cell 2022, 185, 2422–2433.e13. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Guo, Y.; Iketani, S.; Nair, M.S.; Li, Z.; Mohri, H.; Wang, M.; Yu, J.; Bowen, A.D.; Chang, J.Y.; et al. Antibody Evasion by SARS-CoV-2 Omicron Subvariants BA.2.12.1, BA.4, & BA.5. Nature 2022, 4, 2022–2025. [Google Scholar] [CrossRef]
- Sheward, D.J.; Kim, C.; Fischbach, J.; Sato, K.; Muschiol, S.; Ehling, R.A.; Björkström, N.K.; Hedestam, G.B.K.; Reddy, S.T.; Albert, J.; et al. Omicron Sublineage BA.2.75.2 Exhibits Extensive Escape from Neutralising Antibodies. Lancet Infect. Dis. 2022, 22, 1538–1540. [Google Scholar] [CrossRef]
- Arora, P.; Kempf, A.; Nehlmeier, I.; Schulz, S.R.; Jäck, H.-M.; Pöhlmann, S.; Hoffmann, M. Omicron Sublineage BQ.1.1 Resistance to Monoclonal Antibodies. Lancet Infect. Dis. 2022, 23, 22–23. [Google Scholar] [CrossRef]
- Scheaffer, S.M.; Lee, D.; Whitener, B.; Ying, B.; Wu, K.; Liang, C.-Y.; Jani, H.; Martin, P.; Amato, N.J.; Avena, L.E.; et al. Bivalent SARS-CoV-2 MRNA Vaccines Increase Breadth of Neutralization and Protect against the BA.5 Omicron Variant in Mice. Nat. Med. 2022, 1–11. [Google Scholar] [CrossRef]
- Hoffmann, M.; Behrens, G.M.N.; Arora, P.; Kempf, A.; Nehlmeier, I.; Cossmann, A.; Manthey, L.; Dopfer-Jablonka, A.; Pöhlmann, S. Effect of Hybrid Immunity and Bivalent Booster Vaccination on Omicron Sublineage Neutralisation. Lancet Infect. Dis. 2022, 23, 25–28. [Google Scholar] [CrossRef]
- Davis-Gardner, M.E.; Lai, L.; Wali, B.; Samaha, H.; Solis, D.; Lee, M.; Porter-Morrison, A.; Hentenaar, I.T.; Yamamoto, F.; Godbole, S.; et al. MRNA Bivalent Booster Enhances Neutralization against BA.2.75.2 and BQ.1.1. Biorxiv 2022. [Google Scholar] [CrossRef]
- WHO. COVID-19 Weekly Epidemiological Update; WHO: Geneva, Switzeland, 2022; Available online: https://www.who.int/publications/m/item/covid-19-weekly-epidemiological-update---21-december-2022 (accessed on 21 December 2022).




{kind=link}
{kind=link}
{kind=link}
| Entire Cohort | No Recent BTI | Recent BTI | ||
|---|---|---|---|---|
| Number (n) a | 63 | 43 | 20 | |
| Age ** b | 59 (47–65) | 63 (49–67) | 54 (41–60) | |
| Sex a | Female (n) | 38 | 27 | 11 |
| Male (n) | 25 | 16 | 9 | |
| Days between the third and fourth doses **** b | 186 (134–229) | 155 (121–215) | 268 (209–288) | |
| Fourth dose (n) a | Pfizer monovalent | 28 | 22 | 6 |
| Moderna monovalent | 21 | 18 | 3 | |
| Pfizer BA.4/5 | 4 | 1 | 3 | |
| Moderna BA.1 | 10 | 2 | 8 | |
| Days between the third dose and W4-Va3 b | 26 (22–33) | 25 (21–31) | 28 (22–34) | |
| Days between the third dose and M4-Va3 b | 120 (113–126) | 120 (111–126) | 122 (117–129) | |
| Days between the fourth dose and W4-Va4 b | 28 (22–35) | 28 (23–36) | 25 (21–30) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tauzin, A.; Benlarbi, M.; Medjahed, H.; Grégoire, Y.; Perreault, J.; Gendron-Lepage, G.; Gokool, L.; Morrisseau, C.; Arlotto, P.; Tremblay, C.; et al. Humoral Responses against BQ.1.1 Elicited after Breakthrough Infection and SARS-CoV-2 mRNA Vaccination. Vaccines 2023, 11, 242. https://doi.org/10.3390/vaccines11020242
Tauzin A, Benlarbi M, Medjahed H, Grégoire Y, Perreault J, Gendron-Lepage G, Gokool L, Morrisseau C, Arlotto P, Tremblay C, et al. Humoral Responses against BQ.1.1 Elicited after Breakthrough Infection and SARS-CoV-2 mRNA Vaccination. Vaccines. 2023; 11(2):242. https://doi.org/10.3390/vaccines11020242
Chicago/Turabian StyleTauzin, Alexandra, Mehdi Benlarbi, Halima Medjahed, Yves Grégoire, Josée Perreault, Gabrielle Gendron-Lepage, Laurie Gokool, Chantal Morrisseau, Pascale Arlotto, Cécile Tremblay, and et al. 2023. "Humoral Responses against BQ.1.1 Elicited after Breakthrough Infection and SARS-CoV-2 mRNA Vaccination" Vaccines 11, no. 2: 242. https://doi.org/10.3390/vaccines11020242