
A Clinical, Pathological, Epidemiological and Molecular Investigation of Recent Outbreaks of Peste des Petits Ruminants Virus in Domestic and Wild Small Ruminants in the Abu Dhabi Emirate, United Arab Emirates
 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
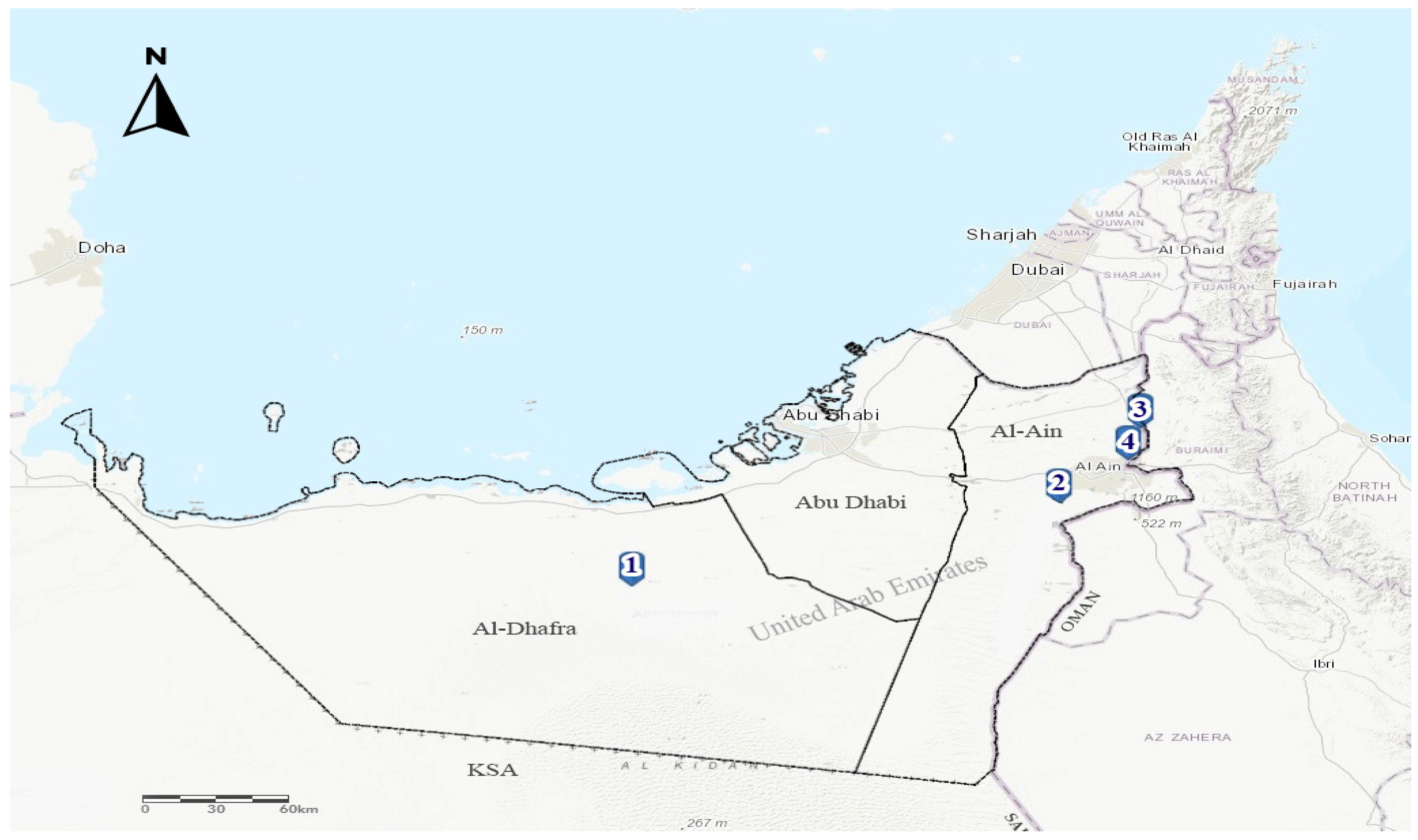
2.1. Epidemiological Data, Necropsy and Sampling
2.2. Histopathological Examination
2.3. Quantitative Real-Time PCR (RT-qPCR) Based Detection of PPRV
2.4. N Gene and F Gene Sequencing and Phylogenetic Analysis
2.4.1. Amplification of N Gene and F Gene
2.4.2. Sanger Sequencing
2.4.3. Sequence Alignment and Phylogenetic Analysis
3. Results
3.1. Outbreak Investigations
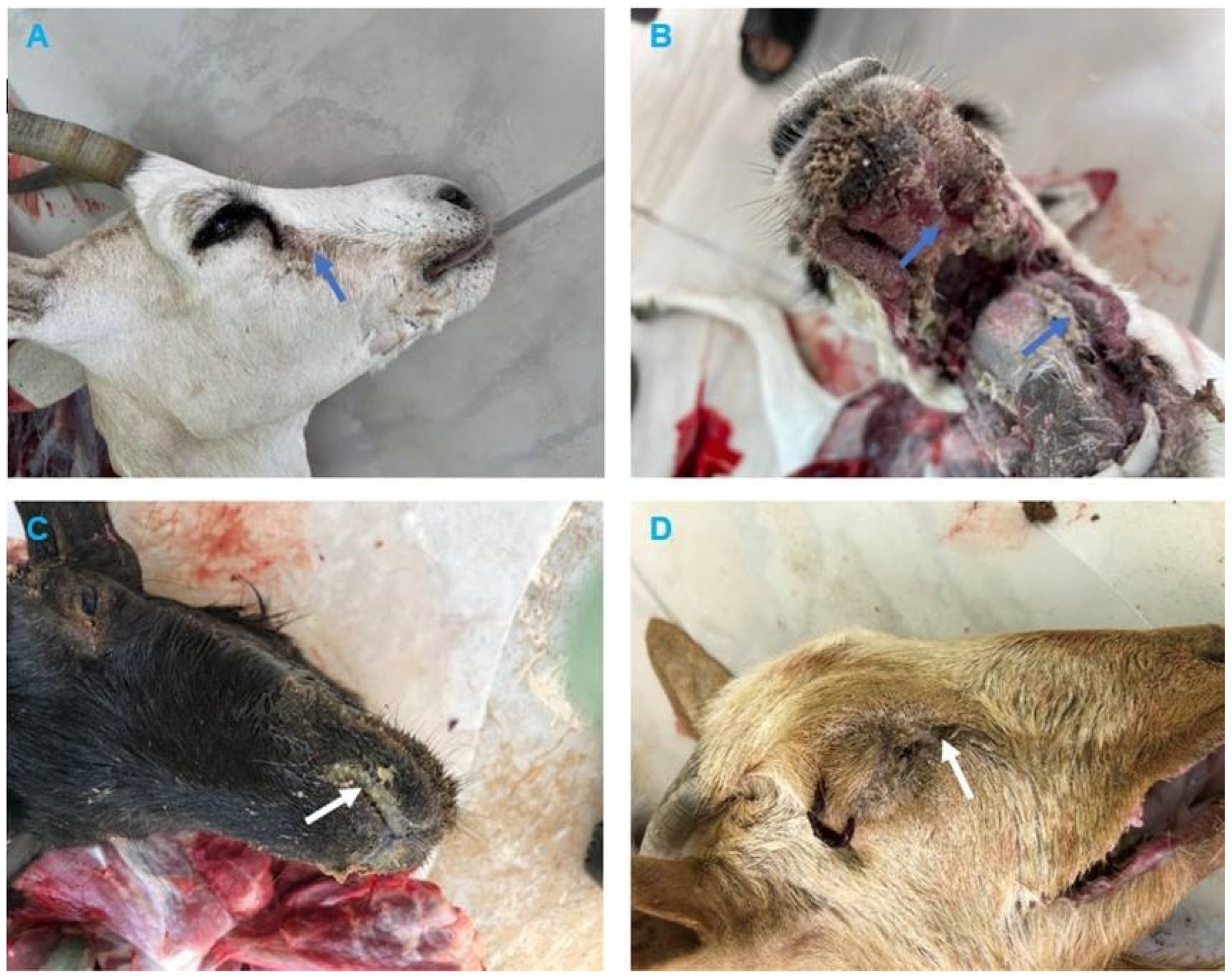
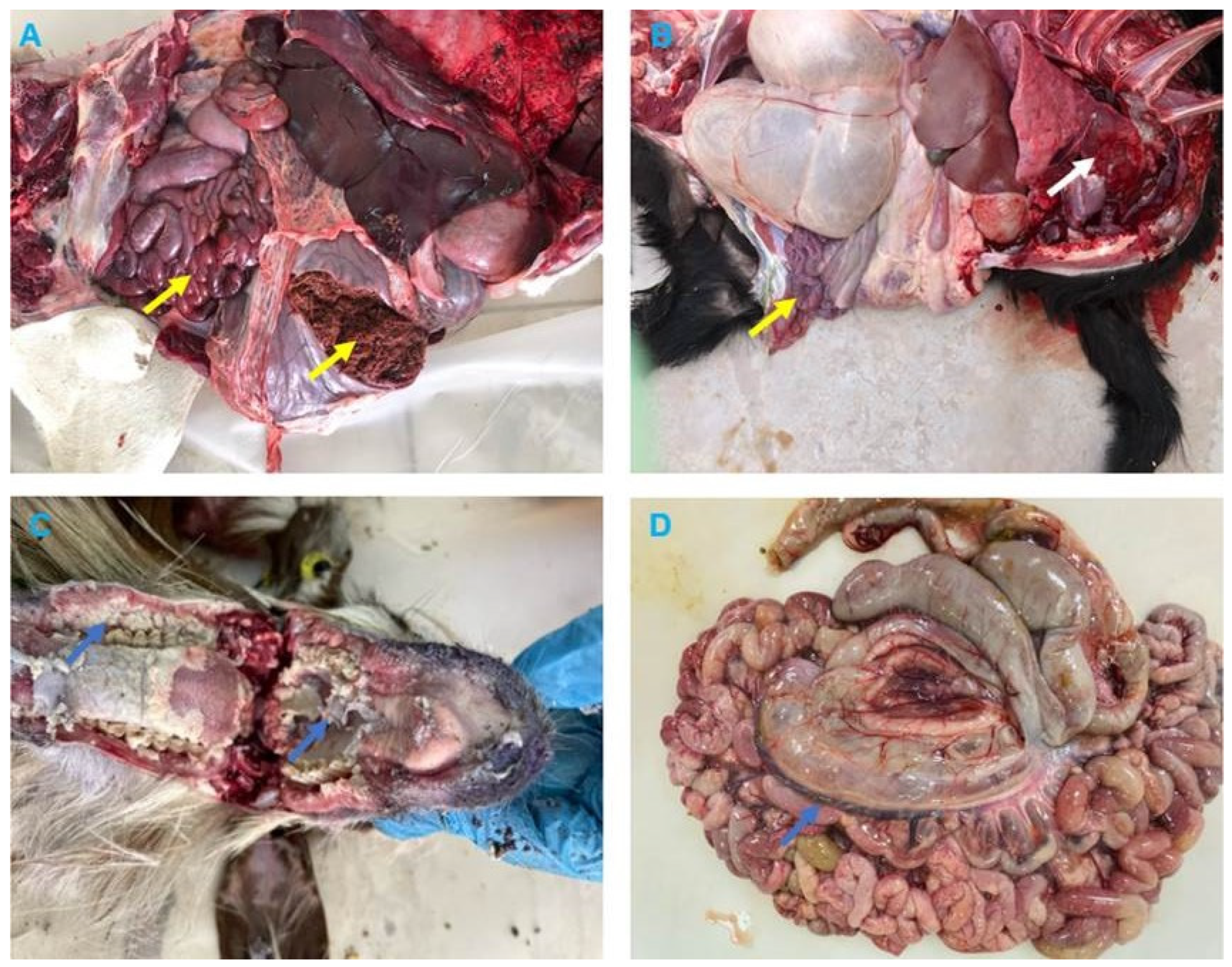
3.2. Clinical and Pathological Findings
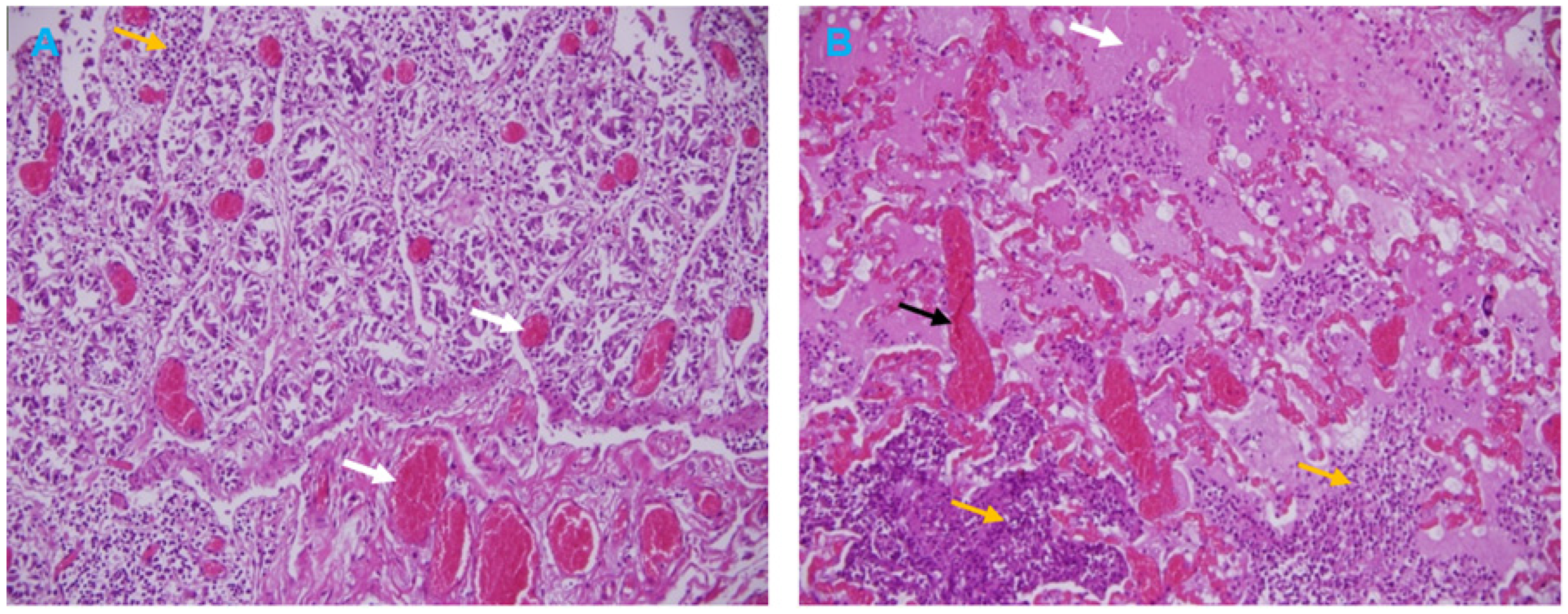
3.3. Histopathological Analysis
3.4. Molecular Detection and Characterization of PPRV
3.4.1. Detection of PPRV by N Gene Real-Time PCR
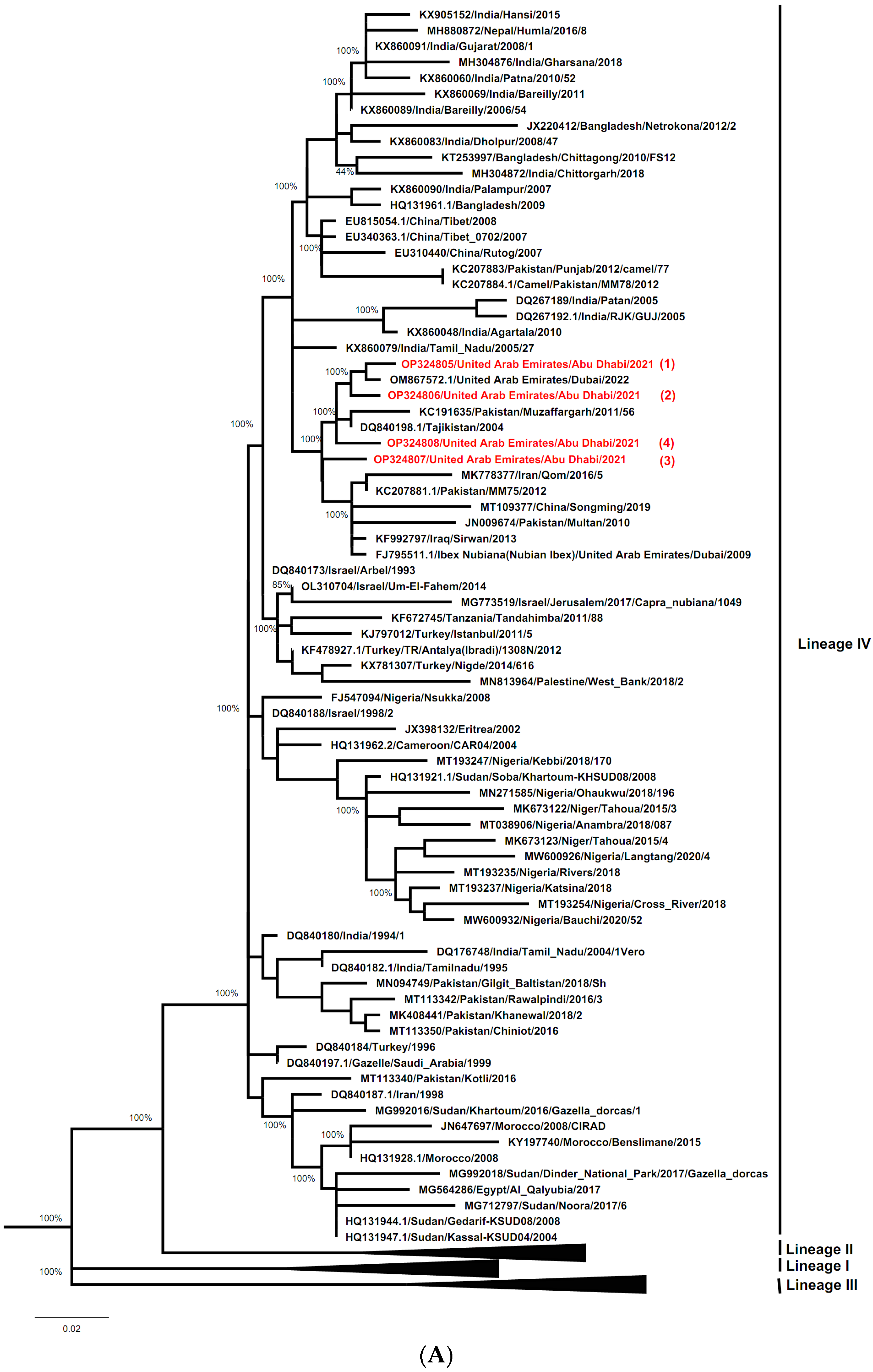
3.4.2. Characterization of PPRV Strains and Determination of its Geographical Lineage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gargadennec, L.; Lalanne, A. La peste des petits ruminants. Bull. Serv. Zoo. AOF 1942, 5, 15–21. [Google Scholar]
- FAO. Global Strategy for the Control and Eradication of PPR; FAO: Rome, Italy, 2015. [Google Scholar]
- Siddell, S.G.; Walker, P.J.; Lefkowitz, E.J.; Mushegian, A.R.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; Knowles, N.J.; Kropinski, A.M. Binomial nomenclature for virus species: A consultation. Arch. Virol. 2020, 165, 519–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, D.; Banyard, A.; Dash, P.; Ozkul, A.; Barrett, T. Full genome sequence of peste des petits ruminants virus, a member of the Morbillivirus genus. Virus Res. 2005, 110, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Maherchandani, S.; Kashyap, S.K.; Singh, S.V.; Sharma, S.; Chaubey, K.K.; Ly, H. Peste des petits ruminants virus infection of small ruminants: A comprehensive review. Viruses 2014, 6, 2287–2327. [Google Scholar] [CrossRef] [Green Version]
- Baron, M.D.; Bataille, A. A curated dataset of peste des petits ruminants virus sequences for molecular epidemiological analyses. PLoS ONE 2022, 17, e0263616. [Google Scholar] [CrossRef]
- Parida, S.; Couacy-Hymann, E.; Pope, R.A.; Mahapatra, M.; Harrak, M.E.; Brownlie, J.; Banyard, A.C. Pathology of peste des petits ruminants. In Peste des Petits Ruminants Virus; Springer: Berlin/Heidelberg, Germany, 2015; pp. 51–67. [Google Scholar]
- Shaila, M.; Shamaki, D.; Forsyth, M.A.; Diallo, A.; Goatley, L.; Kitching, R.; Barrett, T. Geographic distribution and epidemiology of peste des petits ruminants viruses. Virus Res. 1996, 43, 149–153. [Google Scholar] [CrossRef]
- Munir, M. Diagnosis of Peste des Petits Ruminants under Limited Resource Setting: A Cost Effective Strategy for Developing Countries Where PPRV Is Endemic; VDM Publishing: Saarbrucken, Germany, 2011. [Google Scholar]
- Banyard, A.C.; Parida, S.; Batten, C.; Oura, C.; Kwiatek, O.; Libeau, G. Global distribution of peste des petits ruminants virus and prospects for improved diagnosis and control. J. Gen. Virol. 2010, 91, 2885–2897. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, M.; Wang, S.; Liu, Z.; Shen, N.; Si, W.; Sun, G.; Drewe, J.A.; Cai, X. Peste des petits ruminants virus in Heilongjiang province, China, 2014. Emerg. Infect. Dis. 2015, 21, 677. [Google Scholar] [CrossRef] [Green Version]
- Ismail, I.; House, J. Evidence of identification of peste des petits ruminants from goats in Egypt. Arch. Fur Exp. Vet. 1990, 44, 471–474. [Google Scholar]
- Zakian, A.; Nouri, M.; Kahroba, H.; Mohammadian, B.; Mokhber-Dezfouli, M.-R. The first report of peste des petits ruminants (PPR) in camels (Camelus dromedarius) in Iran. Trop. Anim. Health Prod. 2016, 48, 1215–1219. [Google Scholar] [CrossRef]
- Khalafalla, A.I.; Saeed, I.K.; Ali, Y.H.; Abdurrahman, M.B.; Kwiatek, O.; Libeau, G.; Obeida, A.A.; Abbas, Z. An outbreak of peste des petits ruminants (PPR) in camels in the Sudan. Acta Trop. 2010, 116, 161–165. [Google Scholar] [CrossRef]
- Anderson, J.; McKay, J. The detection of antibodies against peste des petits ruminants virus in cattle, sheep and goats and the possible implications to rinderpest control programmes. Epidemiol. Infect. 1994, 112, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, M.; Rajput, Z.I.; Arshed, M.J.; Sarwar, G.; Ali, Q. Evidence of peste des petits ruminants virus (PPRV) infection in Sindh Ibex (Capra aegagrus blythi) in Pakistan as confirmed by detection of antigen and antibody. Trop. Anim. Health Prod. 2011, 43, 745–747. [Google Scholar] [CrossRef] [PubMed]
- Furley, C.; Taylor, W.; Obi, T. An outbreak of peste des petits ruminants in a zoological collection. Vet. Rec. 1987, 121, 443–447. [Google Scholar] [CrossRef]
- Kinne, J.; Kreutzer, R.; Kreutzer, M.; Wernery, U.; Wohlsein, P. Peste des petits ruminants in Arabian wildlife. Epidemiol. Infect. 2010, 138, 1211–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CFSPH. Centre for Food Security and Public Health (CFSPH) 2008: Peste des Petits Ruminants. Available online: http://www.cfsph.iastate.edu/DiseaseInfo/disease.php?name=peste-des-petits-ruminants&lang=en (accessed on 19 April 2011).
- Alemu, B.; Gari, G.; Libeau, G.; Kwiatek, O.; Kidane, M.; Belayneh, R.; Siraw, B.; Wieland, B.; Asfaw, W.; Abdi, R.D. Molecular detection and phylogenetic analysis of Peste des petits ruminants virus circulating in small ruminants in eastern Amhara region, Ethiopia. BMC Vet. Res. 2019, 15, 84. [Google Scholar] [CrossRef]
- Munir, M.; Zohari, S.; Berg, M. Pathophysiology and clinical assessment of peste des petits ruminants. In Molecular Biology and Pathogenesis of Peste des Petits Ruminants Virus; Springer: Berlin/Heidelberg, Germany, 2013; pp. 33–48. [Google Scholar]
- Emikpe, B.; Ezeasor, C.; Sabri, M.; Anosa, V. Clinicopathological evaluation of intranasal, subcutaneous and intramuscular routes of vaccination against intratracheal challenge of Peste des petits ruminants virus in goats. Small Rumin. Res. 2013, 113, 290–296. [Google Scholar] [CrossRef]
- Uzal, F.A.; Plattner, B.L.; Hostetter, J.M. Alimentary system. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals; Elsevier: Amsterdam, The Netherlands, 2016; Volume 2, pp. 1–257.e2. [Google Scholar]
- Couacy-Hymann, E.; Bodjo, C.; Danho, T.; Libeau, G.; Diallo, A. Evaluation of the virulence of some strains of peste-des-petits-ruminants virus (PPRV) in experimentally infected West African dwarf goats. Vet. J. 2007, 173, 178–183. [Google Scholar] [CrossRef]
- Milovanović, M.; Dietze, K.; Joseph, S.; Wernery, U.; Kumar, A.; Kinne, J.; Patteril, N.G.; Hoffmann, B. The Experimental Infection of Goats with Small Ruminant Morbillivirus Originated from Barbary Sheep. Pathogens 2022, 11, 991. [Google Scholar] [CrossRef]
- Dhar, P.; Sreenivasa, B.; Barrett, T.; Corteyn, M.; Singh, R.; Bandyopadhyay, S. Recent epidemiology of peste des petits ruminants virus (PPRV). Vet. Microbiol. 2002, 88, 153–159. [Google Scholar] [CrossRef] [PubMed]
- El Hag Ali, B.; Taylor, W. Isolation of peste des petits ruminants virus from the Sudan. Res. Vet. Sci. 1984, 36, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Kwiatek, O.; Minet, C.; Grillet, C.; Hurard, C.; Carlsson, E.; Karimov, B.; Albina, E.; Diallo, A.; Libeau, G. Peste des petits ruminants (PPR) outbreak in Tajikistan. J. Comp. Pathol. 2007, 136, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Nanda, Y.; Chatterjee, A.; Purohit, A.; Diallo, A.; Innui, K.; Sharma, R.; Libeau, G.; Thevasagayam, J.; Brüning, A.; Kitching, R. The isolation of peste des petits ruminants virus from northern India. Vet. Microbiol. 1996, 51, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Özkul, A.; Akca, Y.; Alkan, F.; Barrett, T.; Karaoglu, T.; Dagalp, S.B.; Anderson, J.; Yesilbag, K.; Cokcaliskan, C.; Gencay, A. Prevalence, distribution, and host range of Peste des petits ruminants virus, Turkey. Emerg. Infect. Dis. 2002, 8, 709. [Google Scholar] [CrossRef]
- World Organization for Animal Health WOAH. World Animal Health Information System WOAH-WAHIS. Quantitative Data. Available online: https://wahis.oie.int/#/dashboards/qd-dashboard (accessed on 15 May 2022).
- Fathelrahman, E.M.; Reeves, A.; Mohamed, M.S.; Ali, Y.M.E.; El Awad, A.I.; Bensalah, O.-K.; Abdalla, A.A. Epidemiology and Cost of Peste des Petits Ruminants (PPR) Eradication in Small Ruminants in the United Arab Emirates—Disease Spread and Control Strategies Simulations. Animals 2021, 11, 2649. [Google Scholar] [CrossRef]
- SowjanyaKumari, S.; Akshata, N.; Balamurugan, V. Peste Des Petits Ruminants in Atypical Hosts and Wildlife: Systematic Review and Meta-Analysis of the Prevalence between 2001 and 2021. Arch. Razi Inst. 2021, 76, 1589. [Google Scholar]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Bao, J.; Li, L.; Wang, Z.; Barrett, T.; Suo, L.; Zhao, W.; Liu, Y.; Liu, C.; Li, J. Development of one-step real-time RT-PCR assay for detection and quantitation of peste des petits ruminants virus. J. Virol. Methods 2008, 148, 232–236. [Google Scholar] [CrossRef]
- Nabi, R.; Hossain, M.S.; Saha, S.; Alam, J.; Giasuddin, M. Molecular epidemiology of Peste des petits ruminants (PPR) in goat. Int. J. Sci. Technol. Res. 2018, 7, 7–12. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Zhao, H.; Njeumi, F.; Parida, S.; Benfield, C.T. Progress towards eradication of Peste des petits ruminants through vaccination. Viruses 2021, 13, 59. [Google Scholar] [CrossRef]
- Rony, M.; Rahman, A.; Alam, M.; Dhand, N.; Ward, M. Peste des Petits Ruminants risk factors and space–time clusters in Mymensingh, Bangladesh. Transbound. Emerg. Dis. 2017, 64, 2042–2048. [Google Scholar] [CrossRef]
- Cameron, A.R. Strategies for the global eradication of peste des petits ruminants: An argument for the use of guerrilla rather than trench warfare. Front. Vet. Sci. 2019, 6, 331. [Google Scholar] [CrossRef]
- Jones, B.A.; Mahapatra, M.; Chubwa, C.; Clarke, B.; Batten, C.; Hicks, H.; Henstock, M.; Keyyu, J.; Kock, R.; Parida, S. Characterisation of Peste Des Petits Ruminants Disease in Pastoralist Flocks in Ngorongoro District of Northern Tanzania and Bluetongue Virus Co-Infection. Viruses 2020, 12, 389. [Google Scholar] [CrossRef]
- Muse, E.A.; Karimuribo, E.D.; Misinzo, G.; Mellau, L.S.; Msoffe, P.L.; Gitao, G.C.; Swai, E.S.; Albano, M.O. Epidemiological investigation into the introduction and factors for spread of Peste des Petits Ruminants, southern Tanzania: Proceeding. Onderstepoort J. Vet. Res. 2012, 79, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mahapatra, M.; Sayalel, K.; Muniraju, M.; Eblate, E.; Fyumagwa, R.; Shilinde, S.; MaulidMdaki, M.; Keyyu, J.; Parida, S.; Kock, R. Spillover of peste des petits ruminants virus from domestic to wild ruminants in the Serengeti ecosystem, Tanzania. Emerg. Infect. Dis. 2015, 21, 2230. [Google Scholar] [CrossRef] [Green Version]
- Wohlsein, P.; Saliki, J. Rinderpest and peste des petits ruminants—The diseases: Clinical signs and pathology. In Rinderpest and Peste des Petits Ruminants; Elsevier: Amsterdam, The Netherlands, 2006; pp. 68–85. [Google Scholar]
- Elzein, E.A.; Housawi, F.; Bashareek, Y.; Gameel, A.; Al-Afaleq, A.; Anderson, E. Severe PPR Infection in Gazelles kept under semi-free range conditions. J. Vet. Med. Ser. B 2004, 51, 68–71. [Google Scholar] [CrossRef]
- Kumar, K.S.; Babu, A.; Sundarapandian, G.; Roy, P.; Thangavelu, A.; Kumar, K.S.; Arumugam, R.; Chandran, N.; Muniraju, M.; Mahapatra, M. Molecular characterisation of lineage IV peste des petits ruminants virus using multi gene sequence data. Vet. Microbiol. 2014, 174, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Kerur, N.; Jhala, M.; Joshi, C. Genetic characterization of Indian peste des petits ruminants virus (PPRV) by sequencing and phylogenetic analysis of fusion protein and nucleoprotein gene segments. Res. Vet. Sci. 2008, 85, 176–183. [Google Scholar] [CrossRef]
- Munir, M.; Zohari, S.; Saeed, A.; Khan, Q.; Abubakar, M.; LeBlanc, N.; Berg, M. Detection and phylogenetic analysis of peste des petits ruminants virus isolated from outbreaks in Punjab, Pakistan. Transbound. Emerg. Dis. 2012, 59, 85–93. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Batch Number/Farm | Date of Collection | Sample Type | Animal Species | RT-qPCR Positive | GenBank Accessions | |
|---|---|---|---|---|---|---|
| F Gene | N Gene | |||||
| 1,092,884/Farm 1 | 7 July 2021 | Lung, Oral swab | Goat | 2 | OP324801 | OP324805 |
| 221,435/Farm 2 | 25 July 2021 | Spleen, Nasal swab, Oral swab, Intestine | Dama Gazelle | 4 | OP324802 | OP324806 |
| 221,258/Farm 3 | 7 July 2021 | Lung, Oral swab, Lymph node | Goat | 3 | OP324803 | OP324807 |
| 1,093,756/Farm 4 | 13 September 2021 | Spleen | Goat | 1 | OP324804 | OP324808 |
| Farm Number | Date of Outbreak Notification | Animal Species | Total Population | No. of Sheep | No. of Goats | No. of Infected | No. of Deaths | Morbidity Rate | Mortality Rate | Case Fatality Rate | Age of Infected Animals | Vaccination Status of Infected Farm | Introduction of New Unvaccinated Animals |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 7 July 2021 | Sheep and Goats | 343 | 60 | 283 | 30 | 6 | 9% | 1.75% | 20% | <3 Months | Unvaccinated | Yes |
| 2 | 25 July 2021 | Dama Gazelle | 20 | 0 | 0 | 16 | 4 | 80% | 20.00% | 25% | 1–2 Years | Unvaccinated | No |
| 3 | 7 July 2021 | Sheep and Goats | 232 | 99 | 133 | 20 | 16 | 9% | 6.90% | 80% | 1 Year | Unvaccinated | Yes |
| 4 | 13 September 2021 | Sheep and Goats | 207 | 79 | 128 | 10 | 4 | 5% | 1.93% | 40% | >6 Months | Vaccinated (11 February 2021) | No |
| Strain | OP324805 | OP324806 | OP324807 | OP324808 | OM867572.1 |
|---|---|---|---|---|---|
| OP324805/UAE/Abu Dhabi/2021 (1) | 98% | 98% | 97% | 99% | |
| OP324806/UAE/Abu Dhabi/2021 (2) | 98% | 98% | 98% | 98% | |
| OP324807/UAE/Abu Dhabi/2021 (3) | 98% | 98% | 97% | 98% | |
| OP324808/UAE/Abu Dhabi/2021 (4) | 97% | 98% | 97% | 98% | |
| OM867572.1/UAE/Dubai/2022 | 99% | 98% | 98% | 98% |
| Strain | OP324801 | OP324802 | OP324803 | OP324804 | OM867572.1 |
|---|---|---|---|---|---|
| OP324801/UAE/Abu Dhabi/2021 (1) | 99% | 100% | 100% | 99% | |
| OP324802/UAE/Abu Dhabi/2021 (2) | 99% | 99% | 99% | 98% | |
| OP324803/UAE/Abu Dhabi/2021 (3) | 100% | 99% | 100% | 99% | |
| OP324804/UAE/Abu Dhabi/2021 (4) | 100% | 99% | 100% | 99% | |
| OM867572.1/UAE/Dubai/2022 | 99% | 98% | 99% | 99% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishag, H.Z.A.; Terab, A.M.A.; Eltahir, Y.M.; El Tigani-Asil, E.T.A.; Khalil, N.A.H.; Gasim, E.F.M.; Yuosf, M.F.; Al Yammahi, S.M.S.; Al Mansoori, A.M.A.; Al Muhairi, S.S.M.; et al. A Clinical, Pathological, Epidemiological and Molecular Investigation of Recent Outbreaks of Peste des Petits Ruminants Virus in Domestic and Wild Small Ruminants in the Abu Dhabi Emirate, United Arab Emirates. Vet. Sci. 2023, 10, 56. https://doi.org/10.3390/vetsci10010056
Ishag HZA, Terab AMA, Eltahir YM, El Tigani-Asil ETA, Khalil NAH, Gasim EFM, Yuosf MF, Al Yammahi SMS, Al Mansoori AMA, Al Muhairi SSM, et al. A Clinical, Pathological, Epidemiological and Molecular Investigation of Recent Outbreaks of Peste des Petits Ruminants Virus in Domestic and Wild Small Ruminants in the Abu Dhabi Emirate, United Arab Emirates. Veterinary Sciences. 2023; 10(1):56. https://doi.org/10.3390/vetsci10010056
Chicago/Turabian StyleIshag, Hassan Zackaria Ali, Abdelnasir Mohammed Adam Terab, Yassir Mohammed Eltahir, El Tigani Ahmed El Tigani-Asil, Nasereldien Altaib Hussein Khalil, Esamt Faisal Malik Gasim, Mohd Farouk Yuosf, Saeed Mohamed Saeed Al Yammahi, Asma Mohammed Amer Al Mansoori, Salama Suhail Mohammed Al Muhairi, and et al. 2023. "A Clinical, Pathological, Epidemiological and Molecular Investigation of Recent Outbreaks of Peste des Petits Ruminants Virus in Domestic and Wild Small Ruminants in the Abu Dhabi Emirate, United Arab Emirates" Veterinary Sciences 10, no. 1: 56. https://doi.org/10.3390/vetsci10010056