
Abstract
Recent evidence suggests that people can simultaneously activate attentional control setting for two distinct colors. However, it is unclear whether both attentional control settings must operate globally across the visual field or whether each can be constrained to a particular spatial location. Using two different paradigms, we investigated participants’ ability to apply independent color attentional control settings to distinct regions of space. In both experiments, participants were told to identify red letters in one hemifield and green letters in the opposite hemifield. Additionally, some trials used a “relevant distractor”—a letter that matched the opposite side’s target color. In Experiment 1, eight letters appeared (four per hemifield) simultaneously for a brief amount of time and then were masked. Relevant distractors increased the error rate and resulted in a greater number of distractor intrusions than irrelevant distractors. Similar results were observed in Experiment 2 in which red and green targets were presented in two rapid serial visual presentation streams. Relevant distractors were found to produce an attentional blink similar in magnitude to an actual target. The results of both experiments suggest that letters matching either attentional control setting were selected by attention and were processed as if they were targets, providing strong evidence that both attentional control settings were applied globally, rather than being constrained to a particular location.
Similar content being viewed by others
Attention is a mechanism by which a limited amount of information is selected to receive preferential processing and facilitates many core abilities such as the conscious recognition of objects in one’s visual world (Becker & Pashler, 2005; Pashler, 1998; Rensink, O'Regan, & Clark, 1997). As a result, a great deal of research has investigated the processes that guide the allocation of attention. This work has established that top-down volitional mechanisms can bias attention toward goal-relevant objects in the environment (Henderson, Brockmole, Castelhano, & Mack, 2007; Williams, 1966) and that volitional attention can be location-based (Posner, 1980), object-based (Kanwisher & Driver, 1992), or feature-based (Folk, Remington, & Johnston, 1992).
Despite extensive research investigating the mechanisms guiding the volitional allocation of attention, some fairly fundamental questions about the capabilities of this system have only recently become the focus of research. For instance, recent studies have investigated whether two distinct feature-based attentional control settings can guide attention simultaneously (Beck, Hollingworth, & Luck, 2012; Becker, Alzahabi, & Jelinek, 2011; Irons, Folk, & Remington, 2012). Two of these studies tracked eye movements while people performed a difficult visual search and found that people were able to constrain overt attention to objects that matched either of two colors that were cued. In addition, the pattern and timing of eye movements suggest that the attentional control settings for both colors were operating simultaneously (Beck, Hollingworth, & Luck, 2012; Becker, Alzahabi, & Jelinek, 2011). These experiments demonstrate an ability to apply two attentional control settings across the entire field at once. Other studies have demonstrated that spatial attention can be directed to at least two distinct spatial regions (Awh & Pashler, 2000; Kramer & Hahn, 1995). The question remains, however, whether distinct attentional control settings can be applied to specific spatial locations. That is, if one was asked to monitor one location for a green item while monitoring a second location for a red item, could the attentional control setting for green be constrained to the green-relevant location or would a green item at the red location capture attention as if it were a target?
This fairly basic question about the functional capabilities of the feature-based attentional system has received very little investigation. To date, most evidence suggests that feature-based attention is applied globally across the visual scene (Liu & Mance, 2011; Serences & Boynton, 2007; Serences, et al., 2005), which would be inconsistent with the view that one could contrain a particular attentional control setting to a specific location. However, previous experiments investigating the global nature of feature-based attention typically had just one relevant feature on a given trial. Situations with only one attentional control setting may encourage people to set the relevance for that feature globally, but one might be able to shift to a more spatially local mode if the task demands it. Constraining an attentional control setting to a distinct location might require a task in which two features are relevant, but each is only relevant when it occurs in a particular location.
Two lines of research suggest that feature-based attention may be able to be constrained to specific locations under situations in which two features are important, but each is important only when it appears in a particular location. The first line comes from experiments investigating value-driven capture of attention. In these experiments, a feature that is associated with reward acquires the ability to involuntarily capture attention. Using this type of value-driven attention capture paradigm, Anderson (2015) found that associating a given feature with high reward only when it appeared in a particular location, leads to attentional capture by the feature only when it appears in the previously rewarded location; when the same feature appears in a previously unrewarded location, it no longer captures attention. This result suggests that the relevance of features can be constrained to a particular spatial context. However, the value-driven capture effect may reflect a form of implicit learning, rather than a volitional application of an attentional control setting. Thus, it is unclear whether the same ability to constrain attention would occur under a more explicit and volitional situation.
Adamo and colleagues (Adamo, Pun, Pratt, & Ferber, 2008) have directly investigated whether explicit instructions would allow one to constrain an attentional control settings to a specific location. To do so, they used a combination of a Posner cuing paradigm and a go, no-go task. A participant was instructed to make a speeded response when a green target appeared to the left or a blue target appeared to the right of fixation but to withhold responses when a green item appeared on the right or a blue item appeared on the left. These stimuli were preceded by a brief cue that could be green or blue and could appear on either the right or left of fixation. They found that the cue had the greatest effect when its color was consistent with the color/location response contingencies. They interpreted this as evidence that the cue captured attention only when it was in the location where that color was relevant.
These results, however, are difficult to interpret given their experimental design; for example, these effects may instead be due to the cue priming the target’s color and/or the target’s location. For instance, consider trials in which a participant was to respond to a green target on the left. A green cue on the left would prime both the location and color of the target, leading to a fast response. Only one of these factors would be primed for a green cue on the right (color) or a blue item on the left (location), leading to intermediate times. Finally, a blue cue on the right would not prime the target’s color or location, leading to the slowest response. Thus, a simple priming account also can explain the pattern of results observed in the Adamo et al. (2008) study.Footnote 1
This priming view is broadly consistent with a follow-up study by Adamo and colleagues (Adamo, Pun, & Ferber, 2010) using the same task while measuring ERPs. Their ERP results suggest that there are independent spatial orienting mechanisms (as indexed by the N2pc) and feature-based orienting (as indexed by the target’s P3 magnitude). In their discussion of how the ERP results support the RT results, the authors conclude that RTs are fastest when the cue matches both the color and location of a subsequent target, because the cue produces both enhanced spatial and feature based selection, leading to fast consolidation of the target into working memory (as indexed by an earlier P3 to the target). By contrast, when the cue matches only the location or the color of the subsequent target, only one selection mechanism operates and the RTs are intermediate. Finally, when the cue matches neither the color nor the location of the subsequent target, the cue does not promote spatial or feature based selection, leading to the slowest RTs. In addition, neither of these early priming components was impacted by the color-location contingencies of the attentional control signals. Instead, the color-location contingency only occurs at a later stage when objects are consolidated into working memory (also see Parrott, Levinthal, & Franconeri, 2010). Thus, the results of these studies may not indicate a true ability to constrain multiple attentional control settings to specific locations. Instead, the cuing procedure may produce two sources of priming that combine to mimic the results of constraining attentional control settings.
A more rigorous test of the independent attentional control setting hypothesis would be to use a paradigm that does not rely on cuing. A study by Lo, Howard, and Holcombe (2012) investigated the ability of participants to constrain two colored attentional control settings without using pre-cues. In their task, a participant monitored two gratings, one on each side of fixation, whose spatial period smoothly changed. Each to-be-monitored grating was superimposed on a second grating, so successful monitoring required selective attention to one of the two gratings at each location. Across conditions subjects were asked to monitor either the same color on both sides of fixation or a red grating on one side of fixation and a green grating on the other. When asked to monitor gratings of two colors, the distractor gratings could either be a novel color (e.g., blue) or could match the color of the target from the other hemifield (e.g., a red distractor grating on the side with a green target grating). After a period of monitoring, the gratings would disappear and the subject was asked to indicate the final spatial period of the attended grating. Errors in reporting the spatial period were equivalent when monitoring a single color (on both sides) or monitoring two colors (one on each side), provided that the distractors gratings were novel colors (e.g., blue and yellow). This finding suggests that there is little cost of maintaining two compared with one attentional control setting. However, when the distractor grating’s color matched the color of the other hemifield’s target, errors increased dramatically suggesting that the ACS were applied globally rather than being constrained to one location. However, the task required the maintenance of the attentional control setting over a period of time, and thus was not a measure of how the attentional control settings constrain the initial deployment of attention.
Experiment 1
To further investigate the ability to constrain each of two attentional control settings to a particular location, we began with an extremely simple method that avoided cuing and thus minimized potential priming confounds. A display of eight heterogeneous colored letters, four per hemifield, was briefly presented and masked. Participants were told to identify the single target letter that appeared in the display. This target was defined as either the red letter on the right or the green letter on the left (or vice versa for other participants). On half the trials, only one of the letters in the display appeared in red or green and it appeared in the hemifield where it was a target. In the remaining half, one of the distractors in the same hemifield as the target matched the target color for the other hemifield (e.g., a red target with a green distractor in the same hemifield). If participants were able to constrain each attentional control setting to its appropriate hemifield, then the “relevant” distractors (those that matched the opposite hemifield’s attentional control setting) should be treated as any other distractor, and performance should be equivalent across conditions. If, however, attentional control settings were set globally, the relevant distractor should compete with the target for attentional selection, and target identification performance should suffer.
One important aspect of the design was that it avoided the use of pre-cues, thereby minimizing the effects of priming. Even so, there was still the potential for trial-to-trial priming effects in which the color of the target on trial N biased the detection of a similarly colored item on trial N+1. To get a purer measure of attentional-control settings and some measure of the intertrial priming effects, distractor effects were measured when the color of the target in the previous trial matched or did not match the current target color.
Methods
Participants
Sixty-one (15 males) college undergraduates with normal or corrected-to-normal vision participated to fulfill course credit.
Stimuli and apparatus
The background of the display was black, and the eight letters appeared around an imaginary clock face with a radius of 10.2 degrees of visual angle. Each letter subtended ~1.5° wide by 1.7° high. Within a hemifield, the four letters were equally spaced from one another, but there was a gap around the horizontal midline, so that the letters on the right appeared at 1:30, 2:30, 3:30, and 4:30 on the clock face. The letters on the left were in symmetrical positions. This gap around the horizontal midline ensured that the letters from each hemifield projected to different hemispheres (Marzi, Mancini, Sperandio, & Savazzi, 2009). The target color was selected at random on each trial. The location, within the appropriate hemisphere, was selected at random for targets and relevant distractors. For each hemifield, the colors of the nonrelevant distractors were randomly selected without replacement, from the colors purple, gray, blue, and yellow. Letters also were selected randomly without replacement from the 26 letters of the English alphabet.
Procedure
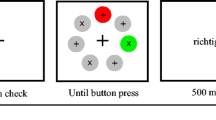
Each trial consisted of a fixation point (1500 ms), followed by a brief display of eight letters (150 ms) that appeared in different colors, followed by a multicolored checkerboard mask (on until response) (Fig. 1). The task was to identify the target letter in the brief display. There was a single target letter in each display. For half of the participants, the target was defined as either a red letter that appeared on the left side of the screen or a green letter that appeared on the right. For the other half of the participants, the colors defining a target were reversed (red-right or green-left). Participants were explicitly told that a red or green item appearing on the opposite side of the screen where it was defined as a target should not be considered a target. Thus, participants should have set an attentional control setting for one target color (red or green) on the left and the opposite target color on the right and constrained these attentional control settings to each hemifield, if possible.

Method used in Experiment 1. The trial depicted is a no relevant distractor trial with a red target
There were 144 trials, half of which were Relevant Distractor trials and half of which were No Relevant Distractor trials, randomly interleaved. Target color was chosen at random on a trial-to-trial basis so there were equal numbers of target that were primed and not primed by the previous target color. In the No Relevant Distractor condition, there was a single red or green letter and it appeared on the side of the screen where it was a target (Fig. 2). In the Relevant Distractor condition, there was a single target letter (either red or green on its appropriate side) and one of the distractors on the target’s side of the screen matched the target color for the other side of the screen (Fig. 2). For instance, a Relevant Distractor trial would have both a red target and a green distractor on the left. If one were able to constrain the red attentional control setting to the left and the green attentional control setting to the right (in this example), the relevant distractors should be treated the same as any other colored distractor. Thus, performance should be identical in the both the No Relevant and Relevant Distractor conditions. If, however, the attentional control settings were applied globally, then a relevant distractor should compete for attention with the target, and given the limited time to detect the target, on some trials the relevant distractor should be selected instead of the target, thereby increasing the error rate.

Examples of the four types of trials used in Experiment 1. The participant was to detect a target letter defined as either a red letter on the left or a green letter on the right. Other participants received instructions to detect green on the left and red on the right
Results
Seven participants were excluded from further analysis for overall accuracy below 5%; the remaining participants had an overall accuracy of 47.4% (SEM = 6.2%). For the remaining participants, we calculated the number of successful target identifications in both distractor conditions and when the color of the previous target matched or did not match the color of the current target (Fig. 3, top). A 2 × 2 repeated measures ANOVA revealed a main effect of distraction condition F(1, 53) = 80.0 , p < 0.001, partial eta2 = 0.601 and a main effect of priming F(1, 53) = 37.38, p < 0.001, partial eta2 = 0.414. The interaction effect was not significant F(1, 53) = 0.298, p = 0.58, partial eta2 = 0.006. Target identification performance was significantly worse, t(53) = 8.94, p < 0.001, in the Relevant Distractor condition (M = 42.9%, SEM= 1.5%) than the No Relevant Distractor condition (M = 55.1%, SEM = 2.o%). This drop in performance indicates that a relevant distractor was a particularly effective distractor, suggesting that both attentional control settings were applied globally rather than being constrained to a single hemifield.

Top panel presents the mean percentage of correct target identifications as a function of whether the trial had a relevant distractor or not and whether the color of the previous target matched the current target. The bottom panel depicts the percentage of trials in which the participant erroneously reported the identity of the relevant distractor or the identity of one of the other distractors in the same hemifield
We also considered the possibility that attentional control settings were partially constrained. If there was an ability to constrain by hemifield but it lacked spatial precision or constraint was graded, trials with distractors nearer to the midline (e.g., at 1:30 and 4:30) might show larger distractor effects than trials with distractors farther from the midline (e.g., at 2:30 and 3:30), because those close to the midline were nearer to the hemifield where their color was relevant. To investigate this possibility, we classified trials based on the location of the distractor with one bin for distractors that appeared in the four locations nearest the midline and another bin for distractors that appeared in the four locations farther from the midline. We did the same binning for target locations. A 2 (target location) x 2 (distractor location) repeated measures ANOVA revealed a main effect of target location, F(1,53) = 51.27, p < 0.001, partial eta2 = 0.492, with better target detection for targets away from the midline (M = 49.5%, SEM = 1.8%) than targets near the midline (M = 36.8%, SEM = 1.9%). There also was a main effect of distractor location, F(1, 53) = 4.98, p = 0.03, partial eta2 = 0.09, with more distraction for distractors further from the midline (M = 41.6 %, SEM = 1.8%) than distractors near the midline (M = 44.7%, SEM = 1.7%). The distractor location by target location interaction was not significant, F(1,53) = 1.67, p = 0.20, partial eta2 = 0.03. This pattern is inconsistent with the view that distractors near the midline are more likely to attract attention. The same bias was apparent in the no distractor trials, F(1, 53) = 43.0, p < 0.001, partial eta2 = 0.45, with better target detection for the location further from the midline (M = 61.8% , SEM = 2.1%) than those near the midline (M = 48.0%, SEM = 2.3%). Thus, there was an overall bias to attend to both targets and distractors that appear further into the hemifield than those close to midline. Whether this resulted from a horizontal bias in search (Gilchrist and Harvey, 2006) or was due to the task demands of our study is unclear. However, the finding that distractors that appear further from the midline were more disruptive than those that appear near the midline provides evidence against a type of partial or graded constraint of attention to a particular hemifield.
For the relevant distractor condition, we also calculated the percentage of trial (Fig. 3, bottom) in which there was a relevant distractor intrusion (the participant identified the relevant distractor rather than the target) and the percentage of trials in which there was an irrelevant distractor intrusion (the participant identified an irrelevant distractor rather than the target). There were significantly more, t(53) = 3.99, p < 0.001, relevant distractor intrusions (M = 8.9%, SEM = 1.5%) than irrelevant distractor intrusions (M = 2.7%, SEM = 0.29%). Considering only distractor trials in which there was an error, we compared the rate of intrusion errors to the rate one would have expected by chance (1/26 given 26 letters in the alphabet). The rate of relevant distractor intrusions (M = 16.4%, SEM = 2.8%) was significantly greater than chance, t(53) = 4.53, p < 0.001, but rate of irrelevant distractor intrusions (M = 4.7%, SEM = 0.46%) did not differ from chance, t(53) = 1.51, p > 0.08.
Discussion
If each attentional control setting was constrained to a hemisphere, we would have expected that the relevant distractor should have been equivalent to any other distractor. Thus, performance should have been equivalent for the Relevant and No Relevant distractor conditions, and relevant distractors should have been identified at the same rate as an irrelevant distractor. By contrast, if attentional control settings were applied globally, the relevant distractor should have been a particularly effective distractor, leading to a reduction in target identifications and an increase in the number of errors that were due to the participant reporting the relevant distractor rather than the target.
Our results are consistent with the hypothesis that attentional control settings are applied globally. Target identification performance was worse in the relevant distractor condition than the irrelevant distractor condition. In addition, the drop in target identification performance in the relevant distractor condition appeared to result from an increase in the number of trials in which the relevant distractor rather than the target was identified. Indeed, the difference in target identification performance between the relevant and irrelevant distractor conditions was almost exactly equivalent to the number of trials in the relevant distractor condition in which participants identified the relevant distractor. This pattern of data is completely consistent with the interpretation that attentional control settings are applied globally.
However, another possible explanation for this pattern exists. It is possible that red and green were simply more salient (e.g., brighter) than the other colors and thus captured attention more effectively regardless of the attentional control settings. While we thought this unlikely, a control experiment definitively ruled out this explanation. In the control experiment, participants searched for targets of one color – half searched for green (n = 12) and half searched for red (n = 12). Displays were similar to Experiment 1 and every trial had a target. In the red target condition, one of the distractors in the same hemifield as the target was a green item in half of the trials. In the other half of the trials, no green item appeared. This pattern was reversed for participants searching for green. If red and green were particularly salient and captured attention regardless of the attentional control settings, performance in this control experiment should have mimicked performance in Experiment 1; namely, red and green distractors should have captured attention more than distractors presented in other colors. However, the presence of a red or green distractor made no difference in target detection performance, t(23) = 0.31, p = 0.76.
The control experiment rules out an explanation of Experiment 1’s results on the basis of bottom-up saliency and suggests that the relevant distractors in Experiment 1 were particularly good at competing for attention, because they matched the target color of the other hemifield. In short, Experiment 1 suggests that attentional control settings were not constrained to their respective locations, thus the relevant distractors captured attention as if they were targets, thereby reducing target detection accuracy and leading to more errors where the identity of the distractor was reported.
In addition, in Experiment 1 we found an intertrial priming effect that was additive with the distractor effects. Although the priming effects were intertrial rather than caused by a cue, the existence of these priming highlights that experiments that do not control for potential priming effects may be confounded by them. As we have discussed, the results of Adamo and colleagues that appear to demonstrate an ability to constrain attention control settings may instead have been due to priming effects, albeit priming due to the cue rather than intertrial priming effects.
While our data suggest that one cannot completely constrain two attentional control settings to different locations, it is still possible that the attentional control settings were at least partially constrained. That is, Experiment 1 demonstrated that a relevant distractor was more disruptive than other distractors, but it was unable to show that it was as disruptive as an actual target would have been. Without such a comparison, the results of Experiment 1 could be consistent with a partial ability to constrain attentional control settings. In order to establish that both attentional control settings were applied globally, it is important to confirm that an item captures attention equally well when it appears as relevant distractor and when it appears as a target. In Experiment 2, we include trials in which two targets appear, to allow a comparison of the distracting effects of a relevant distractor to the distracting effects of an actual target.
In addition, we switched from the methods of Experiment 1 to a modified version of an attentional blink (AB) task (Raymond, Shapiro, & Arnell, 1992). Our rationale for this switch was twofold. First, Adamo and colleagues have suggested an early attentional selection phase is not sensitive to the constraints of attentional control settings, and they only influence a later process of encoding into working memory. Because the method of Experiment 1 involved briefly presenting the target and relevant distractor simultaneously,one might claim that the method produced competition at an early selection stage; and thus one might not find evidence for the ability to constrain attentional control settings. By contrast, in our modified AB design, the target and distractor are separate in time and thus should not compete with each other for very early selection. Indeed, the AB seems to result from a failure for the second target to be processeed at the stage of access into working memory (Vogel, Luck, & Shapiro, 1998), the very stage that Adamo and colleagues suggest should be sensitive to color-location attentional control setting contingencies.
Second, the AB method allowed us to have one condition, the lag 1 condition, that was very similar to the Adamo cueing method. This allowed us to investigate whether we could replicate their findings in a situation when priming could play a role and to examine what happened once priming was eliminated.
Experiment 2
The goal of this experiment was to assess whether there was a partial ability to constrain attentional control settings or if attempting to constrain an attentional control setting to a particular location resulted in a gradient of spatial attention. In Experiment 1, we found that relevant distractors impaired target identification; however, it was unclear whether the magnitude of the effect was smaller than if that stimulus had been presented as a target in its relevant hemifield. Toward this end, participants simultaneously monitored two rapid serial visual presentation (RSVP) streams of colored letters. Participants were asked to monitor the leftward stream for a red letter while monitoring the right stream for a green letter. We then determined whether a distractor that matched the target color for the opposite stream would capture attention and produce an AB of similar magnitude to the same stimulus when it was presented as a target in its relevant hemifield. If people could effectively constrain the attentional control setting for red to the left location, then a red item presented as a target in the left stream should produce a sizeable AB for a subsequent target, but the same red item appearing on the right should be similar to any other distractor and should not produce an AB. By contrast, if people set a global attentional control setting for red, the red item should capture attention when it appears in either stream, and thus the item might produce a sizeable AB even when it appeared on the right.
The AB paradigm has some additional properties that make it a good method for testing the question of whether each of two attentional control settings can be constrained to a specific location. First, it allowed us to have a lag 1 condition, which was fairly analogous to the methods of Adamo. At lag 1, the relevant distractor immediately preceded the target, much as the cue did in the Adamo method. If the relevant distractor produced priming effects we might find that we replicate Adamo’s conclusions at lag 1. However, at later lags, there are intervening items between the relevant distractor and the target, and thus priming should be reduced. Thus in the same experiment, we can investigate the pattern of results when priming is likely (lag 1) and when it not (lag 2). Second, we hypothesized that the ability to constrain an attentional control setting to a spatial location would be optimal when the location was relatively circumscribed and fixed throughout the trial. In this method, there are only two relevant locations that are fixed throughout the experiment. The method we used in Experiment 1 did not have this property, which may have contributed to our finding that attentional control settings were applied globally rather being constrained by location.
Methods
Participants
Fifty-three college undergraduates (15 males) who had normal or corrected-to-normal vision participated for course credit.
Procedure
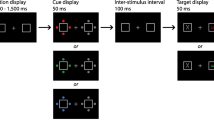
The experiment was programmed in E-prime, run on PCs with 19-inch CRT monitors running at 100 Hz and was run individually in sound attenuated booths. After receiving verbal instructions and performing the informed consent procedure, participants completed 7 practice and 126 experimental AB trials. Each trial consisted of a central fixation point flanked by two RSVP streams, each comprised of 16 colored letters (Fig. 4). Letters (~1.3° × 1.3°) were presented for 150 ms each and were immediately replaced by the next letter in the stream. The streams were ~2.5° degrees into the periphery to ensure that the stimuli would project to opposite hemispheres (Marzi, Mancini, Sperandio, & Savazzi, 2009).

Examples of the four trial types used in Experiment 2. The participant monitored both streams looking for red letters on the right and green letters on the left. Each letter was presented for 150 ms and immediately replaced by the next letter
Participants were instructed to monitor both streams simultaneously and that target letters would be red in the left stream and green in the right stream. Participants were explicitly told that an item matching the other stream’s color was not a target and that a given trial could have: 1) no targets in either stream; 2) a single target in one of the streams; or 3) a target in both streams. At the end of each trial, participants were asked whether there was a target in each stream. If they answered yes for a stream, they were asked to indicate whether that stream’s letter was from the first or second half of the alphabet. We switched to this method of responding, rather than having participants identify the specific target letter, for a number of reasons. First, this method has been used successfully in prior AB studies (Moore & Weissman, 2010; Serences, et al., 2005). Second, the demands placed on the participants were already fairly high and we wanted to minimize demands associated with response selection.
Trial types
In the 18 No-Target Control trials, there were no red or green items in either stream and each letter was randomly selected to appear in blue, yellow, purple, or gray with the caveat that two sequential letters in the same stream could not appear in the same color. The remaining 108 trials were comprised of 36 trials from each of the three types of target-present conditions. The Two-Target Control condition had a single target in each stream. The first target could appear in the either the 3rd, 5th, or 7th position within the stream, and the second target appeared in the opposite stream at lag 1, 2, or 5. In half of these trials, the first target was a red item on the left; in the other half, the first target was a green item on the right. These control trials were used to establish the magnitude of the AB in a situation where there were two targets. In Opposite-Stream Distractor trials a target appeared in only one of the streams, but it was preceded (lag 1, 2, or 5) by a distractor of the same color as the target that appeared in the opposite stream (e.g., a red distractor on the right preceded a red target on the left). In Same-Stream Distractor trials a target appeared in only one of the streams, but it was preceded (lag 1, 2, 5) by a distractor within that stream that matched the target color of the opposite stream (e.g., a green distractor on the left preceded a red target on the left). This design resulted in 12 trials per cell of the design (given the three lags).
All target and relevant distractor letters (those set to be the target color of the stream opposite of where they appeared) were selected from the set excluding the letters L-O, to avoid confusion near the boundary between the first and last half of the alphabet. In addition, when a target was selected from the first half of the alphabet, the distractor was selected from the second half. Thus, if a participant erroneously identified the distractor rather than the target, it would be an incorrect response. All factors were counterbalanced and the trial order was randomized.
Results
Successful target identifications
For each participant, we calculated the percentage of targets that were correctly identified at each lag for four types of targets: The first (T1) and second (T2) target in the Two-Target Control condition, the target in the Opposite-Stream Distractor (DxT; distractor in the opposite hemifield as the target) condition, and the Same- Stream Distractor (DT) condition (Fig. 5). If attentional control settings can be constrained to their locations, then the relevant distractor should behave like any other distractor letter, thus targets should be detected at a rate similar to the first target (T1) in the Two-Target Control condition. If attentional control settings are set globally (e.g., without any spatial gradient or location-based contingency), then relevant distractors should capture attention to the same extent as actual targets thereby producing an AB, thus targets in the distractor trials should behave like the second target (T2) in the Two-Target Control condition. Although lag 1 trials were included to evaluate the influence of priming (see below), the main comparison of interest was between lag 2, where the AB should be strongest, and lag 5, where one should have recovered from the AB.

Mean identification accuracy for the target is plotted as function of the different trial types and lag. Given the two-stage response method (did the screen contain a target, was it in the first or second half of the alphabet) chance performance was 25%. Error bars represent the within subject standard error (Loftus & Masson, 1994)
To evaluate whether target identification performance in the distractor trials was similar to either T1 or T2 in the Two-Target control condition, we performed two 3 (conditions) x 2 (lag 2 and 5) within subject ANOVAs. The first included the two distractor conditions and T1, and the second included the two distractor conditions and T2. The ANOVA that included T1 and the two distractor conditions revealed a main effect of trial type, F(2,104) = 12.08, p < 0.001 partial eta2 = 0.19, main effect of lag, F(1, 52) = 20.86, p < 0.001 partial eta2 = 0.29, and a trial type by lag interaction, F(2,104) = 3.10, p = 0.049, partial eta2 = 0.06. The source of the interaction was that both distractor conditions produced a large AB at lag 2 that was not present for T1. The ANOVA that included T2 and the two distractor conditions found a main effect of lag, F(1, 52) = 48.02, p < 0.001 partial eta2 = 0.48, but there was no longer a significant effect of trial type, F(2, 104) < 1 partial eta2 = 0.01, nor a significant trial type X lag interaction, F(2,104) = 1.70, p = 0.19 partial eta2 = 0.03. The main effect of lag results, because there was a sizable AB at lag 2 relative to lag 5 for all three conditions. Importantly at lag 2, performance was equivalent for T2 and both distractor conditions, F(2, 104) = 0.133, p = 0.88, partial eta2 = 0.003. That is, performance in the distractor trials matched performance for T2.
This pattern of results is completely consistent with the predictions of the global attentional control setting hypothesis. In short, attentional control settings were not constrained to their respective locations; the magnitude of the AB was equivalent for targets following relevant distractors and actual targets. Thus, the relevant distractors captured attention as if they were targets, producing an AB for targets that followed relevant distractors at lag 2.
False alarms
As a second test of the claim that relevant distractors were treated like a target, we calculated false-alarm rates (i.e., when people claimed that there had been a target in a particular stream when none occurred). The logic of this analysis was as follows: If a relevant distractor captured attention and was treated like a target, it might be reported as a target thereby increasing false alarms. False-alarm rates for the No-Target Control Condition trials served as a baseline measure to evaluate whether the relevant distractors increased the false-alarm rate. We calculated each participant’s mean percentage of false alarms for the No-Target Control trials, the Opposite-Stream Distractor (DxT) trials, and the Same-Stream Distractor (DT) trials (Fig. 6). A within-subjects ANOVA on these means was significant, F(2, 104) = 18.17, p < 0.001. Planned comparisons showed that the false-alarm rate was significantly higher for both types of distractor trials than the No-Target Control trials, both t(52) > 3.93, both p < 0.001, suggesting that these relevant distractors truly captured attention and were processed to the point that they were occasionally mistaken for targets.

Mean percentage of false alarms as a function of trial type. False alarms were defined as a participant indicating that a target had been present in a stream that had no target. Error bars represent the within subject standard error (Loftus & Masson, 1994)
We also found more false alarms in the Opposite Stream than Same-Stream distractor trials, t(52) = 3.60, p = 0.001. However, we think this is probably due to a reporting bias rather than a difference in the likelihood of each type of distractor capturing attention. A false alarm in a Same-Stream trial would involve seeing a green distractor in the left stream (the red-relevant stream) and then falsely reporting that it had appeared in the right stream (which had no target or relevant distractor). Thus, a false alarm in Same-Stream trials involves mislocating the relevant distractor. By contrast, a false alarm in an Opposite-Stream trial would involve seeing a green distractor in left stream, and mistakenly reporting it as a target in the left stream. Thus a false alarm in an Opposite-Stream trial involves a breakdown of the color-location contingency rules, but does not require one to mislocate an item. We think people are unlikely to mislocate an item, and thus should be less likely to falsely report a distractor as appearing in a stream that had no distractor or target, thereby reducing the false-alarm rate for Same-Stream distractors.
Lag 1 data
We included a lag 1 condition in the design to investigate whether there would be color and location priming effects of immediately preceding distractors. Under a priming argument, T2s are primed by neither their color nor spatial location, and thus should perform poorly. Targets in Opposite Stream Distractor Trials were primed by their color but not location, and targets in the Same Stream Distractor Trials were primed by location but not color; thus, each should produce moderately better performance than T2. Following this priming account performance was worse for T2 than either the Opposite or Same Stream Distractor conditions, t(52) = 3.26 and 4.13 respectively (both p < 0.003). Importantly, this difference between T2 and distractor conditions disappeared for lag 2 when priming should be severely reduced by the intervening items, t(52) = 0.5 and 0.15 respectively (both p > 0.6). Thus, at lag 1 we have what looks like evidence for the constrain of attention control settings; however, they seem to be driven by priming effects rather than actual volitional top-down control settings.
It is worth noting that while we ascribe the results of lag 1 to priming, the type of priming we are referring to is somewhat unique. Many researchers who investigate priming are interested in how repeating a relevant feature across trials leads to faster processing of that feature. For instance, in “priming of pop-out” participants become faster to detect a color singleton if the color of the singleton remains constant across trials (Maljkovic & Nakayama, 1994). This type of between-trial priming seems to depend on the feature being relevant to one’s goals or responses, tends to occur across trials, and has a relatively protracted time course (for a review see Kristjánsson & Campana, 2010). By contrast, the priming we are referring to is a faster, short-term type of priming that can occur in response to irrelevant cues. While there is less research on this type of cuing, there is some. For instance, Folk and Remington (2008) had participants search for a red or green color singleton. The search array was preceded by a noninformative red or green spatial cue. They found that responses to the target were faster when the color of the cue matched the color of the target than when it matched the other potential target color. Thus, the color of the cue primed the subsequent processing of a matching target. In addition, the within trial color congruency effect did not depend on whether the cue was at a valid or invalid location, suggesting that the effect was due to priming the feature (color) rather than the spatial location.
Finally, there are other possible interpretations of the lag-1 effects that would not necessarily indicate low-level priming. Research investigating contingent capture and using a single attentional control setting provides ample evidence that a distractor that matches the attentional control setting can capture attention away from a relevant location, thereby reducing speed and accuracy of detecting a subsequent target at the relevant location (Folk, Leber, & Egeth, 2002). Research using multiple attentional control settings suggests that while multiple attentional control setting can be activated in parallel, when a distractor appears that matches one of those attentional control settings, that particular setting temporarily becomes the only active attentional control setting, thereby reducing the detection of subsequent targets that match a different one of the attentional control settings (Moore & Weissman, 2010). Both of these processes are fairly short-lived and thus would have the most influence at our lag-1 condition (Jefferies & Di Lollo, 2009; Moore & Weissman, 2010).
These processes could combine to produce our lag 1 pattern of results. For the T2 condition, the appearance of T1 would have drawn attention to the wrong location (stream) and made the wrong color the active attentional set, thereby producing very poor performance. For the same stream distractor condition, the distractor would have drawn attention to the correct location but activated the wrong attentional set. For the opposite stream distractor condition, the distractor would have activated the correct attentional set but would have drawn attention to the wrong spatial location (stream). Given that each of these distractor conditions had one process helping and the other hurting performance, overall performance may have been somewhat intermediate. Thus, it is possible that these feature-based and location-based mechanisms combine to produce our lag 1 results. Our data cannot determine whether the cause of our lag 1 results is priming per se or a result of these location and feature-based mechanisms. Regardless, even under the alternative scenario, the relevant distractors capture attention, suggesting that the attentional control settings are not being constrained to their relevant locations.
Discussion
We asked participants to selectively monitor one RSVP stream for red items while monitoring a second RSVP stream for green items and found that a distractor letter that matched the opposite stream’s target color produced an AB for a target that followed it at lag 2. This finding of an AB demonstrates that a distractor matching the other stream’s target color was selected by attention, thereby providing strong evidence that the attentional control setting for that color was not effectively constrained to the location where it was relevant. In addition, the magnitude of the AB at lag 2 caused by these relevant distractors was equivalent to the AB created by an actual target, suggesting that the relevant distractor was about as likely to be selected by attention as a real target. In conjunction with the findings of Experiment 1, these data provide strong evidence that participants set their feature-based attentional sets globally across the entire visual field, rather than constraining them to the spatial locations where they were relevant.
The false-alarm data provide additional evidence for the global nature of these attentional sets. Relative to trials with no targets or distractors, relevant distractors increased the number of times that participants reported that a target appeared in a stream that contained no target. The finding that these relevant distractors impact false-alarm rates suggests that the relevant distractors were occasionally treated as if they were targets, providing further evidence that the attentional control settings were not constrained to the relevant spatial location but were set globally.
It is worth noting that our AB results are consistent with a recent report by Irons and Remington (2013). They also used two RSVP streams and investigated the blink caused by a distractor that matched the target color of the opposite stream. Consistent with our results, they found that these relevant distractors produced an AB. In their method, however, they only had a single target on each trial. As a result, they were unable to determine whether the magnitude of the blink effect caused by a relevant distractor was as large as the blink cause by a true target and they had no ability to investigate false alarms. Our inclusion of a two-target condition allowed us to more rigorously test this idea and allowed us to demonstrate that the magnitude of the AB produced by targets and relevant distractors was similar. In addition, we were able to demonstrate that the presence of a relevant distractor within a stream that had no target resulted in an increased rate of erroneously reporting that there was a target within the stream. Both of these findings provide compelling evidence that attentional control settings were applied globally.
Finally, our lag 1 condition provided evidence that appear to support the ability to constrain an attentional control setting to a specific location. However, this was only found for the condition (lag 1) that allowed for strong priming effects. We believe that these results highlight the importance of considering possible priming effects when evaluating the ability to constrain attentional control settings by location. In situations like our lag 1 condition and previous studies that allow for strong priming effects, priming can produce results that can be confused for constraints of attentional control settings.
Conclusions
We used two very different experimental paradigms to investigate the question of whether two attentional control settings for distinct colors could each be constrained to a separate hemifield. Both of our designs controlled for potential priming effects, and as a result, we found no evidence for independent attentional control setting. Across both methods, we found evidence that both attentional control settings were applied globally rather than being constrained to a hemifield.
In our paradigms, failing to constrain the attentional control setting to the appropriate hemisphere was always harmful to performance, and thus participants should have constrained settings if they could have. In addition, each attentional control setting was relevant in a single hemifield. Given prior work suggesting independent attentional resources in each hemisphere for tracking objects (Alvarez & Cavanagh, 2005) and selecting stimuli for consolidation into visual short-term memory (Delvenne & Holt, 2012), we felt that this hemifield approach would be the most likely scenario in which people could constrain two distinct attentional control settings. Thus, we believe that our failure to find evidence for constraining the attentional control settings is relatively strong evidence suggesting that people apply attentional control settings globally.
It is worth noting that prior work suggests that the magnitude of the attentional blink is reduced (at least at lag 2) when the two critical objects are presented to opposite hemifields (Scalf, et al., 2007; Irons and Remington, 2013). In our data, such a hemifield effect would have predicted that the Same Stream Distractor Trials would have shown a larger attentional blink than the Opposite Stream Distractor and T2 trials at lag 2. We did not find such a difference; instead, performance for all three of those trial types was equivalent at lag 2. Why we did not find such an effect is unclear. Our method was most similar to Irons and Remington, who also looked at the ability to constrain attention control settings by hemifield. One possibility for the difference is that our stimuli were less peripheral (2.5° vs. 5°) than theirs; our stimuli were peripheral enough to ensure that the stimuli projected exclusively to different hemispheres (Marzi, et al., 2009), but the hemifield advantage may require more distal stimuli than we used. Despite this discrepancy, our main finding that people did not seem able to constrain attentional control setting by hemifield is consistent with those of Irons and Remington (2013).
Although our conclusions are at odds with the conclusion of Adamo and colleagues (Adamo et al., 2010a, b; Adamo, et al., 2008), we have provided evidence that one can get results that appear to support the constraining of attentional control settings by location but only when the methods allow for priming effects. Thus, we believe that their results are likely produced by priming that mimics contraint of attentional control settings rather than an ability to constrain them in a volitional top-down manner.
Our results also differ from those of Anderson (2015). He showed that value-driven attentional capture could be based on color-location contingencies. However, the mechanisms involved in the implicit learning associated with value-driven capture are likely quite different from the volitional top-down mechanisms that we have investigated. As Anderson points out, value driven effects rely critically on providing explicit reward during training (Anderson, Laurent, & Yantis, 2011), occur much more rapidly than simple search history or perceptual learning (Anderson, 2013), and can occur without explicit memory of the stimulus-award associations (Anderson & Yantis, 2013).
Lo, Howard, and Holcome’s study used a continuous monitoring task, and their results suggest that two attentional controls settings can be applied gloabally but cannot be spatially constrained. The current study expands on this work by demonstrating that this also is the case for the initial deployment of attention. Finally, that work and the current findings are consistent with conclusion on the global spread of feature-based attention (Liu & Hou, 2011; Liu & Mance, 2011; Serences & Boynton, 2007) and recent work investigating the issue of whether two attentional control settings can be constrained to a particular location (Irons & Remington, 2013).
It is worth noting that our finding holds when the two features defining the attentional control settings are from the same feature dimension (in this case color). We used two colors, because previous work suggests that color is a particularly effective feature on which to base an attentional control setting (Williams, 1966). Furthermore, work arguing for the ability to simultaneously activate two attentional control settings (Beck, et al., 2012; Becker, et al., 2011; Irons et al., 2012), and those arguing for one’s ability to constrain multiple attentional control settings to particular locations used color (Adamo et al., 2010a, b; Adamo, et al., 2008; Parrott, et al., 2010). However, on the basis of this color work, we cannot determine whether the conclusion would hold when the two attentional control settings are defined by features from different dimensions (e.g., color and orientation). Adamo, Wozny, Pratt, and Ferber (2010) concluded that people could constrain attentional control settings from different feature dimensions (color and shapes). However, that paper used a similar cuing method to their color papers, so the same priming confound may be present. Even so, future work would need to determine the extent to which people can constrain attentional control setting based on different dimensions.
In summary, the data presented provide no evidence that attentional control settings for two distinct colors are able to be constrained to separate hemispheres. We demonstrated this in two distinct paradigms, suggesting that the effects may reflect a general failure to be able to constrain each of two attentional control settings to a particular location. We cannot definitively rule out the possibility that under specific conditions one might be able to do so; however, we believe our results provide strong evidence that doing so is not the norm. This finding imposes a limit on the ability to flexibly configure multiple attentional control settings for optimal selection and has implications for understanding how top-down volitional control of attention functions. It also may have implications for the design of complex display systems; if a particular color is important somewhere in the display, it is likely to capture attention throughout the display.
Notes
Arguing against a priming effect in the Adamo et al. (2008) study is the fact that the location-match and color-match conditions showed no priming compared to a condition in which no cue was presented; however, this was only true for Experiment 1, not Experiment 2. Note that none of the conditions were statistically compared to the no cue condition.
References
Adamo, M., Pun, C., Pratt, J., & Ferber, S. (2008). Your divided attention, please! The maintenance of multiple attentional control sets over distinct regions in space. Cognition, 107(1), 295–303.
Adamo, M., Pun, C., & Ferber, S. (2010a). Multiple attentional control settings influence late attentional selection but do not provide an early attentional filter. Cognitive Neuroscience, 1(2), 102–110.
Adamo, M., Wozny, S., Pratt, J., & Ferber, S. (2010b). Parallel, independent attentional control settings for colors and shapes. Attention, Perception, & Psychophysics, 72(7), 1730–1735. doi:10.3758/APP.72.7.1730
Alvarez, G. A., & Cavanagh, P. (2005). Independent Resources for Attentional Tracking in the Left and Right Visual Hemifields. Psychological Science, 16(8), 637–643.
Anderson, B. A. (2013). A value-driven mechanism of attentional selection. Journal of Vision, 13(3), 7.
Anderson, B. A. (2015). Value-driven attentional capture is modulated by spatial context. Visual Cognition, 23(1–2), 68-81.
Anderson, B. A., & Yantis, S. (2013). Persistence of value-driven attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 39, 6–9.
Anderson, B. A., Laurent, P. A., & Yantis, S. (2011). Value-driven attentional capture. Proceedings of the National Academy of Sciences United States of America, 108, 10367–10371.
Awh, E., & Pashler, H. (2000). Evidence for split attentional foci. Journal of Experimental Psychology: Human Perception and Performance, 26(2), 834–846.
Beck, V. M., Hollingworth, A., & Luck, S. J. (2012). Simultaneous control of attention by multiple working memory representations. Psychological Science, 23(8), 887–898. doi:10.1177/0956797612439068
Becker, M. W., & Pashler, H. (2005). Awareness of the continuously visible: Information acquisition during preview. Perception & Psychophysics, 67(8), 1391–1403.
Becker, M. W., Alzahabi, R., & Jelinek, S. (2011). Effective Attentional Filtering By The Union Of Two Distinct Colors: Eye-Tracking Evidence. Journal of Vision, 11(11), 1328.
Delvenne, J. F., & Holt, J. L. (2012). Splitting attention across the two visual fields in visual short-term memory. Cognition, 122(2), 258–263.
Folk, C. L., & Remington, R. W. (2008). Bottom-up priming of top-down attentional control settings. Visual Cognition, 16(2-3), 215–231.
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18(4), 1030–1044.
Folk, C. L., Leber, A. B., & Egeth, H. E. (2002). Made you blink! Contingent attentional capture produces a spatial blink. Perception & Psychophysics, 64(5), 741–753.
Gilchrist, I. D., & Harvey, M. (2006). Evidence for a systematic component within scan paths in visual search. Visual Cognition, 14(4-8), 704–715.
Henderson, J. M., Brockmole, J. R., Castelhano, M. S., & Mack, M. (2007). Visual saliency does not account for eye movements during visual search in real-world scenes. Amsterdam: Elsevier.
Irons, J., & Remington, R. (2013). Can attentional control settings be maintained for two color–location conjunctions? Evidence from an RSVP task. Attention, Perception, & Psychophysics, 1-14. doi: 10.3758/s13414-013-0439-8
Irons, J. L., Folk, C. L., & Remington, R. W. (2012). All set! evidence of simultaneous attentional control settings for multiple target colors. Journal of Experimental Psychology: Human Perception and Performance, 38(3), 758–775. doi:10.1037/a0026578
Jefferies, L. N., & Di Lollo, V. (2009). Linear changes in the spatial extent of the focus of attention across time. Journal of Experimental Psychology: Human Perception and Performance, 35(4), 1020.
Kanwisher, N., & Driver, J. (1992). Objects, attributes, and visual attention: Which, what, and where. Current Directions in Psychological Science, 1(1), 26–31.
Kramer, A. F., & Hahn, S. (1995). Splitting the beam: Distribution of attention over noncontiguous regions of the visual field. Psychological Science, 6(6), 381–386.
Kristjánsson, Á., & Campana, G. (2010). Where perception meets memory: A review of repetition priming in visual search tasks. Attention, Perception, & Psychophysics, 72(1), 5–18.
Liu, T., & Hou, Y. (2011). Global feature-based attention to orientation. Journal of Vision, 11(10), 8. doi:10.1167/11.10.8
Liu, T., & Mance, I. (2011). Constant spread of feature-based attention across the visual field. Vision Research, 51(1), 26–33.
Lo, S.-Y., Howard, C. J., & Holcombe, A. O. (2012). Feature-based attentional interference revealed in perceptual errors and lags. Vision Research, 63, 20–33.
Loftus, G. R., & Masson, M. E. J. (1994). Using confidence intervals in within-subject designs. Psychonomic Bulletin & Review, 1(4), 476–490.
Maljkovic, V., & Nakayama, K. (1994). Priming of pop-out: I. Role of features. Memory & Cognition, 22(6), 657–672.
Marzi, C. A., Mancini, F., Sperandio, I., & Savazzi, S. (2009). Evidence of midline retinal nasotemporal overlap in healthy humans: A model for foveal sparing in hemianopia? Neuropsychologia, 47(13), 3007–3011.
Moore, K. S., & Weissman, D. H. (2010). Involuntary transfer of a top-down attentional set into the focus of attention: Evidence from a contingent attentional capture paradigm. Attention, Perception, & Psychophysics, 72(6), 1495–1509.
Parrott, S. E., Levinthal, B. R., & Franconeri, S. L. (2010). Complex attentional control settings. The Quarterly Journal of Experimental Psychology, 63(12), 2297–2304.
Pashler, H. E. (1998). The psychology of attention (p. 494). Cambridge: The MIT Press.
Posner, M. I. (1980). Orienting of attention. The Quarterly Journal of Experimental Psychology, 32(1), 3–25.
Raymond, J. E., Shapiro, K. L., & Arnell, K. M. (1992). Temporary suppression of visual processing in an RSVP task: An attentional blink? Journal of Experimental Psychology: Human Perception and Performance, 18(3), 849–860.
Rensink, R. A., O'Regan, J. K., & Clark, J. J. (1997). To see or not to see: The need for attention to perceive changes in scenes. Psychological Science, 8(5), 368–373.
Scalf, P. E., Banich, M. T., Kramer, A. F., Narechania, K., & Simon, C. D. (2007). Double take: Parallel processing by the cerebral hemispheres reduces attentional blink. Journal of Experimental Psychology: Human Perception and Performance, 33(2), 298.
Serences, J. T., & Boynton, G. M. (2007). Feature-Based Attentional Modulations in the Absence of Direct Visual Stimulation. Neuron, 55(2), 301–312.
Serences, J. T., Shomstein, S., Leber, A. B., Golay, X., Egeth, H. E., & Yantis, S. (2005). Coordination of Voluntary and Stimulus-Driven Attentional Control in Human Cortex. Psychological Science, 16(2), 114–122.
Vogel, E. K., Luck, S. J., & Shapiro, K. L. (1998). Electrophysiological evidence for a postperceptual locus of suppression during the attentional blink. Journal of Experimental Psychology: Human Perception and Performance, 24(6), 1656–1674.
Williams, L. G. (1966). The effect of target specification on objects fixated during visual search. Perception & Psychophysics, 1(9), 315–318.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Becker, M.W., Ravizza, S.M. & Peltier, C. An inability to set independent attentional control settings by hemifield. Atten Percept Psychophys 77, 2640–2652 (2015). https://doi.org/10.3758/s13414-015-0964-8
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-015-0964-8