
Abstract
Changes in sustained attention, attentional selectivity, and attentional capacity were examined in a sample of 113 participants between the ages of 12 and 75. To measure sustained attention, we employed the sustained-attention-to-response task (Robertson, Manly, Andrade, Baddeley, & Yiend, Neuropsychologia 35:747–58, 1997), a short continuous-performance test designed to capture fluctuations in sustained attention. To measure attentional selectivity and capacity, we employed a paradigm based on the theory of visual attention (Bundesen, Psychological Review 97:523–547, 1990), which enabled the estimation of parameters related to attentional selection, perceptual threshold, visual short-term memory capacity, and processing capacity. We found evidence of age-related decline in each of the measured variables, but the declines varied markedly in terms of magnitude and lifespan trajectory. Variables relating to attentional capacity showed declines of very large effect sizes, while variables relating to attentional selectivity and sustained attention showed declines of medium to large effect sizes, suggesting that attentional control is relatively preserved in older adults. The variables relating to sustained attention followed a U-shaped, curvilinear trend, and the variables relating to attentional selectivity and capacity showed linear decline from early adulthood, providing further support for the differentiation of attentional functions.
Similar content being viewed by others

The study of cognitive aging has revealed that cognitive functions follow different trajectories across the lifespan. For example, crystallized intelligence or knowledge representations remain largely stable across the lifespan, while fluid intelligence or cognitive processes such as processing speed, working memory, and long-term memory show a substantial decline across the lifespan, beginning in early adulthood and following a systematic, linear decline over the decades (Park, 2000). Atrophy of the brain (Raz, 2004; Raz & Rodrigue, 2006) is one factor that may underpin decline in cognitive processing with age, but the relationship between brain atrophy and cognitive decline is still not well understood (Cabeza, Nyberg & Park, 2005). One exception is the clear concordance between frontal-lobe atrophy and the marked decline of executive functioning displayed by older adults (Park, 2000; Park, Polk, Mikels, Taylor & Marshuetz, 2001; West, 1996). The frontal-lobe hypothesis (West, 1996) proposed that cognitive processes supported by the prefrontal cortex would manifest decline at an earlier age and in greater magnitude than would cognitive processes supported by nonfrontal regions. This hypothesis subsumed earlier explanations of cognitive aging, which drew upon specific executive functions, such as attentional capacity (Craik & Byrd, 1982) or inhibition (Dempster, 1992; Zacks & Hasher, 1997), in order to account for the decline in cognitive function due to age. Alternative accounts of cognitive aging have suggested that more general or global mechanisms, such as slowed processing speed (Salthouse, 1996) or a general deterioration of neuronal integrity (Baltes & Lindenberger, 1997), mediate cognitive decline.
Attention is at the heart of this discussion of cognitive aging. As an executive-control process that is supported in large part by the frontal lobes, it is a potential casualty of the aging process. As a fundamental cognitive process that influences other aspects of cognition, it may also act as a mediator of decline in other cognitive abilities, such as memory and reasoning (Craik, 2006). However, attention is a complex cognitive process, and studies examining the effect of aging on various aspects of attention have yielded different conclusions.
Attention has been defined as a set of processes that enable “the maintenance of goal-directed behaviour in the face of multiple, competing distractions” (Parasuraman, 1998). Attentional ability has been characterised as having three main aspects, labelled as “executive attention,” “selectivity,” and “intensity” (Parasuraman, 1998; Raz & Buhle, 2006; Sturm, 1996; VanZomeren & Brouwer, 1994). Studies investigating the effects of aging on executive attention, using for instance the dual-task (Pashler, 1994) or task-switching (Rogers & Monsell, 1995) paradigms, have shown that older adults have greater difficulty on these tasks than do younger adults (Kramer, Scialfa, Peterson & Irwin, 2001; Reimers & Maylor, 2005; Verhaeghen & Cerella, 2002). Investigations into the effects of age on the intensity aspects of attention have largely focused on sustained attention measured by vigilance tasks. These studies have produced inconsistent results, possibly due to the complexity of the vigilance tasks themselves (Davies & Parasuraman, 1982; W. A. Rogers, 2000). One very large-scale study, involving over 500 participants from the ages of 18 to 91, found little evidence of any age-related decrement in sustained attention ability, as measured by the Mackworth Clock Test, a 62-min sensory vigilance task (Giambra & Quilter, 1988). Studies of the selectivity aspect of attention have employed visual search tasks (Treisman & Gelade, 1980) and visuospatial orienting tasks (Posner, 1980). Studies of visual search have revealed intact feature search but significantly slowed conjunction search in older adults (Trick & Enns, 1998). Studies of visuospatial orienting have suggested that the ability to covertly orient to visual stimuli remains intact in older adults following both endogenous and exogenous cues (Greenwood, Parasuraman & Haxby, 1993).
The purpose of the present study was to document changes in central aspects of attention—namely, sustained attention and attentional selectivity and capacity—across the lifespan. Sustained attention, or the ability to maintain alertness and focus over time, was measured using the sustained-attention-to-response task (SART; Robertson, Manly, Andrade, Baddeley & Yiend, 1997). The SART is a well-validated measure of sustained attention that has been found to significantly correlate with self-reported everyday attentional slips in normal populations (Smilek, Carriere & Cheyne, 2010); has proved useful in the detection of sustained-attention deficits in clinical populations, such as people with head injury (McAvinue, O’Keeffe, McMackin & Robertson, 2005) or attention-deficit hyperactivity disorder (Johnson, Kelly, Bellgrove, Barry, Cox, Gill & Robertson, 2007); and has been related to the activation of right-hemisphere frontal–parietal regions thought to underlie sustained attention (Manly et al., 2003).
Visual selective attention is the ability to focus processing on relevant features and objects while ignoring irrelevant features and objects within the context of a limited attentional or processing capacity. To assess attentional selectivity and capacity, we employed a paradigm based on the theory of visual attention (TVA; Bundesen, 1990). The TVA was first proposed by Bundesen as a computational theory that provided quantitative accounts for a wide range of attentional effects reported in the psychological literature. At the heart of TVA are two equations, and Bundesen, Habekost, and Kyllingsbæk (2005, in the neural TVA [NTVA]) provided a neurophysiological interpretation of these equations. In the NTVA, one of the two basic mechanisms of selective attention, filtering (selection of objects), changes the number of cortical neurons in which an object is represented so that this number increases with the behavioural importance of the object. Another mechanism of selection, pigeonholing (i.e., selection of features), scales the level of activation in neurons coding for a particular feature. By means of these mechanisms, behaviourally important objects and features are likely to win the biased competition (Desimone & Duncan, 1995) to become encoded into visual short-term memory (VSTM). The VSTM system is conceived as a feedback mechanism that sustains activity in the neurons that have won the attentional competition. NTVA accounts both for a wide range of attentional effects in human performance (error rates and reaction times) and a wide range of effects observed in single cells (firing rates) in the primate visual system (for an overview, see Bundesen & Habekost, 2008; Kyllingsbæk, 2006).
TVA has laid the foundation for TVA-based assessment of attentional functions, which has many advantages as compared with conventional clinical tests of visual attention: Performance is analysed into separate functional components (specificity); the method can reveal deficits that go undetected by conventional clinical testing (sensitivity); the measurement error can be quantified, and in most cases shown to be minor (reliability); and the measures that are obtained are not bound to the tasks used, but grounded in a general theory of visual attention (validity). In the pioneering study, Duncan et al. (1999) showed how analysis of partial- and whole-report performance in terms of the parameters defined by TVA enables a very specific measurement of attention deficits in visual neglect patients (see also Bublak et al., 2005; Finke et al., 2005). TVA-based assessment has now been used in studies of simultanagnosia (Duncan et al., 2003), integrative agnosia (Gerlach, Marstrand, Habekost & Gade, 2005), developmental dyslexia (Dubois et al., 2010), alexia (Habekost & Starrfelt, 2006; Starrfelt, Habekost & Gerlach, 2010; Starrfelt, Habekost & Leff, 2009), Huntington’s disease (Finke et al., 2007), Alzheimer’s disease (Bublak, Redel & Finke, 2006; Bublak et al., 2009; Redel et al., 2012), and the effects of stroke in particular parts of the brain (Habekost & Bundesen, 2003; Habekost & Rostrup, 2006, 2007; Peers et al., 2005; see Habekost & Starrfelt, 2009, for a review). TVA-based assessment enables the estimation of parameters related to the span of VSTM (storage capacity of K objects), the rate of encoding into VSTM (processing capacity of C objects/s), the perceptual threshold (minimum effective exposure duration of t 0 ms), and the efficiency of selecting targets rather than distractors (selectivity α, defined as the attentional weight of a distractor divided by the attentional weight of a target).
Using a cross-sectional design involving participants from the ages of 12 to 75, we employed a SART- and TVA-based assessment in order to measure age-related changes in sustained attention and attentional selectivity and capacity. Specifically, we examined whether significant age-related decline was evident for each variable, and if so, the magnitude and lifespan trajectory of such decline.
Method
Participants
A group of 113 volunteers, including 49 males and 64 females, ranging in age from 12 to 75, participated in this study. Table 1 presents a breakdown of the sample by age groups in terms of age, gender, and education level. All participants were recruited and tested by the first author at the Institute of Neuroscience, Trinity College Dublin. All participants were living within (or nearby) the Dublin area. The 12-year-old children and the teenagers (15- to 17-year-olds) were recruited from two neighbouring primary and secondary schools that were known to the first author and were located in a near-suburb of Dublin city. In each case, a school representative (i.e., the principal, in the case of the primary school, and a teacher, in the case of the secondary school) facilitated recruitment by providing parents and students with information leaflets. Children and teenagers (who were all under the age of 18) gave informed assent, and their parents provided informed consent. The adult participants (ages 20+) were largely recruited through the Psychology Department Participant Panel. This panel consists of members of the general public who have expressed an interest in participating in psychological research. Prior to joining the panel, volunteers are asked to complete a detailed questionnaire including a variety of questions covering demographic variables and medical and psychiatric history. A random selection of panel members within each of the desired age brackets was made, and members were invited to participate by way of letter and of an accompanying information leaflet, which were followed up with a phone call by the lead author. A small number of young adults (largely in the 20s age group) had responded to poster advertisements placed around Trinity College. These posters invited willing volunteers to participate in a study on “Attention across the Lifespan.” They were subsequently given the same information leaflet that was posted to members of the Participant Panel. The exclusion criteria included a history of psychiatric disorder or neurological insult. Prior to testing, the participants’ near and colour vision were screened using the Revised Sheridan Gardiner Test (Sheridan, 1970) and the Ishihara Test for Colour Blindness (Ishihara, 1960). This study was granted ethical approval by the Department of Psychology Ethics Committee, and all adult participants gave informed consent prior to participation.
Materials
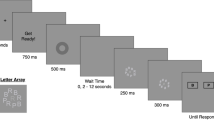
Combi-TVA task
The Combi-TVA task (Vangkilde, Coull & Bundesen, 2012) involved a brief flash of red and blue letters on the computer screen, with the participant’s task being to report all red letters (targets) but to ignore the blue letters (distractors). The test took approximately 40 min for each participant and included 324 trials arranged into nine blocks. Each trial consisted of the presentation of a red fixation cross (1,000 ms), the stimulus display, and a postdisplay mask (500 ms). Each participant was instructed to fixate on the red cross and to verbally report the red letters that he or she saw when the screen went blank, following the mask presentation (see Fig. 1). The researcher typed the letters as the participant reported them. Three stimulus display types were presented: whole report of six targets, whole report of two targets, or partial report of two targets and four distractors. The six-target whole-report trials were presented for 10, 20, 50, 80, 140, and 200 ms. The two-target whole-report trials and the partial-report trials were presented for 80 ms. TVA modelling procedures (Dyrholm, Kyllingsbæk, Espeseth & Bundesen, 2011; Kyllingsbæk, 2006) were applied to estimate the following parameters for each participant:
-
t 0 : Perceptual threshold. The minimum exposure duration (in milliseconds) at which letter identification was better than chance.
-
K: Visual short-term memory capacity. The maximum number of letters that the participant could report.
-
C: Visual processing capacity. The rate of encoding into VSTM, expressed in terms of letters per second.
-
α (Alpha): Efficiency of top-down selection. A value reflecting the participant’s ability to focus on the target letters and to ignore distractors (calculated by dividing the estimated attentional weight of a distractor by the attentional weight of a target). A value of 0 indicates perfect selectivity, with higher values indicating poorer selectivity. A value of 1 or higher indicates complete nonselectivity on the part of the participant.

Illustration of the TVA paradigm
Sustained-attention tasks
The SART (Robertson et al., 1997) measures fluctuations in sustained attention by measuring the ability to inhibit a response to an infrequent target stimulus in the context of maintaining an ongoing, monotonous action. Numbers between 1 and 9 were presented on screen, and participants were required to press a button for every number that appeared but to withhold their response to the number 3 (see Fig. 2). In the fixed version of the task, numbers appeared in a fixed sequence from 1 to 9, and in the random version, numbers appeared in a pseudorandom order. The numbers appeared in white against a black background and remained on screen for 313 ms. Each digit was followed by a mask (a cross within a circle), which lasted for 1,126 ms. Embedded within the mask period was a response cue (63 ms), which was a thicker cross within the circle. The mask period consisted of the following sequence of images: mask (125 ms), response cue (63 ms), mask (375 ms), and fixation (a cross; 563 ms). The participants were asked to respond on the appearance of the response cue in order to minimise differences in inter- and intraindividual response speeds (see Johnson, Kelly, et al., 2007; Johnson, Robertson, Kelly, Silk, Barry, Dáibhis & Bellgrove, 2007). There were 225 digits in all, including 25 no-go targets (the number 3) and 200 go trials (all other numbers). Each SART lasted 5.4 min. The three measures of sustained attention were errors of commission (ERC; i.e., the number of times a participant pressed in response to the number 3), errors of omission (ERO; i.e., number of times a participant did not press for a go-trial number), and the variability of reaction times, expressed as the coefficient of variation (RTCov; i.e., standard deviation/mean reaction time).

Depiction of the fixed and random sustained-attention-to-response tasks
Procedure
Participants attended a single testing session, during which they performed the Combi-TVA task, the SARTfixed, and the SARTrandom, in that order. Prior to completing these tasks, the participants’ near and colour vision were screened using the Revised Sheridan Gardiner Test (Sheridan, 1970) and the Ishihara Test for Colour Blindness (Ishihara, 1960). Analysis of the results proceeded in two steps. First, the trajectory of lifespan changes was examined by dividing the sample into eight age brackets (12-year-olds, teens, and those in their 20s, 30s, 40s, 50s, 60s, and 70s) and plotting the mean values for each age group on each variable. The trajectory was further examined by conducting a regression of each variable upon age in order to examine whether a linear or curvilinear function best characterised the trajectory of each variable. The existence and magnitude of decline on each variable was then examined by comparing younger (20s/30s) and older (60s/70s) adults in independent-samples t tests. Effect sizes corresponding to selectivity, capacity, and sustained attention, derived from the younger–older adult comparison and the regression analyses, were then compared in order to determine whether the magnitude of age-related change was significantly different for these attentional functions.
Results
Trajectory of age-related change
Figures 3 and 4 describe the trajectory of the sustained-attention and attentional-selectivity and -capacity variables across the lifespan by plotting the mean values for each age group (12-year-olds, teens, 20s, 30s, 40s, 50s, 60s, and 70s) on each variable. An examination of the plots by eye suggested that the TVA parameters tended to show improvement from age 12 to the teens, a peak in the teenage years (or the 20s, in the case of alpha), followed by a more or less linear decline thereafter. In contrast, the sustained-attention variables seemed to follow U-shaped trajectories, with poor performance in the childhood and teenage years, a plateau across young and middle adulthood, and performance deteriorating again in older adulthood.

Mean values of TVA parameters for each age group from children to those in their 70s. Vertical lines represent standard errors of the means

Mean value of sustained attention variables for each age group from children to those in their 70s. Vertical lines represent standard errors of the means
These observations were confirmed using regression analyses. Note that the data for the 12-year-olds were not included in the regression analyses, as inclusion of these data would require more complex models (e.g., cubic) that would pertain to both age-related development and age-related decline. Figures 5 and 6 present scatterplots that display the relationship between age and each of the sustained-attention, attentional-selectivity, and attentional-capacity variables. Lowess lines were fitted to the scatterplots in order to help visualise the relationship between age and each of the variables. Lowess lines represent the relationship between two variables by tracing the trend of the data, and they are based on no assumptions about the form of the underlying relationship (Cohen, Cohen, West & Aiken, 2003). The lowess line appeared as a straight line (suggesting a linear relationship) for each of the attentional-selectivity and -capacity variables, but as a curved, U-shaped line (suggesting a curvilinear relationship) for each of the sustained-attention variables.

Scatterplots and lowess lines representing the relationship between age and each TVA parameter. Note that the data for the 12-year-olds are not represented in the scatterplots

Scatterplots and lowess lines representing the relationship between age and each sustained-attention variable. Note that the data for the 12-year-olds are not represented in the scatterplots
Both linear and quadratic regression equations were fitted, and values of R 2, analyses of variance (ANOVAs), and coefficient t tests were examined to determine whether each relationship was best described by a linear or a quadratic equation (Cohen et al., 2003). Table 2 presents the results of the regression analyses for each variable. The relationship between each TVA parameter and age was best represented by a linear regression function, while the relationship between each sustained-attention variable and age was best represented by a quadratic function.
Magnitude of age-related change
One-tailed independent-samples t tests were used to examine whether there was a significant decline from young adulthood (20/30s age groups) to older adulthood (60s/70s age groups) on each variable. Where Levene’s test for the equality of variances was statistically significant, the t value corresponding to an analysis in which equal variances were not assumed was adopted. Tables 3 and 4 present the means, standard deviations, t test results, and effect sizes for the attentional-selectivity, attentional-capacity, and sustained-attention variables. Effect sizes were calculated using Cohen’s d: (Meanold – Meanyoung)/SD pooled (Howell, 2008). Significant declines from younger to older adulthood were evident on every variable, but the declines were of markedly different effect sizes. The largest effect sizes were seen on visual processing capacity, C (d = 2.02), and VSTM capacity, K (d = 1.32). Medium to large effect sizes were observed for all sustained-attention variables (d = 0.67–1.09). Medium effect sizes were observed for perceptual threshold, t 0 (d = 0.56), and the efficiency of top-down attentional selection, α (d = 0.52). An examination of the R 2 values presented in Table 2 reveals a similar pattern in terms of the magnitude of the relationship between age and each attention variable. The highest R 2 values were obtained for C (43.1 %) and K (29.5 %). These were followed by the sustained-attention variables, with R 2 values ranging from 11.4 % to 20.8 %. The lowest R 2 values were obtained for α (10.8 %) and t 0 (8.2 %).
To directly compare the effect sizes (d and R) relating to attentional selectivity, attentional capacity, and sustained attention, composite effect sizes representing selectivity, capacity, and sustained attention were calculated. For selectivity, the effect sizes relating to α were used. For capacity, the effect sizes relating to K and C were averaged. Although t 0 is considered to be an aspect of attentional capacity within the TVA paradigm, it was not included in the composite effect size for capacity due to the large discrepancy between the obtained effect sizes for t 0 and K and C, which raises the possibility that the measurement of t 0 was not sensitive enough to detect age-related decline in the present paradigm (see the Discussion). In addition, previous research has shown that t 0 is not strongly correlated with K and C, which have a strong correlation (McAvinue et al., 2012). For sustained attention, the effect sizes relating to the sustained-attention variables (RandomERC, RandomERO, RandomRTCov, FixedERC, FixedERO, and FixedRTCov) were averaged. Table 5 presents the composite effect sizes for attentional selectivity, attentional capacity, and sustained attention. The values for d were derived from the young-adult–older-adult comparison (see Tables 3 and 4), and the values for R were derived from the regression analyses (see Table 2).
Visual inspection of the composite effect sizes revealed a similar pattern, with the effect sizes being largest for attentional capacity (d = 1.67, R = .6), followed by sustained attention (d = 0.9, R = .4), and finally attentional selectivity (d = 0.52, R = .33). A test of homogeneity was used to examine whether the variability in effect sizes was statistically significantly different from what would be expected from sampling error alone. Q statistics were calculated using the equations presented in Cooper (2010, pp. 186–187) so as to facilitate comparisons between the three attentional effect sizes in terms of d and R. For each comparison, the Q statistic was compared to critical values of the chi-square distribution, corresponding to probability levels of .01 and .05. Pertinent values are presented in Table 6. The findings were very similar for d and R. The comparisons revealed significant variability among the three effect sizes for selectivity, capacity, and sustained attention (for d, Q = 7.76, df = 2, p < .05; for R, Q = 6.24, df = 2, p < .05). The composite effect sizes for capacity were significantly greater than those for selectivity (for d, Q = 7.64, df = 1, p < .01; for R, Q = 5.69, df = 1, p < .05). There were no significant differences between the effect sizes for selectivity and sustained attention (for d, Q = 0.93, df = 1, p > .05; for R, Q = 0.31, df = 1, p > .05). The differences between the effect sizes for capacity and sustained attention were approaching significance (for d, Q = 3.35, df = 1, p > .05; for R, Q = 3.37, df = 1, p > .05).
Discussion
The purpose of this study was to examine age-related change in central aspects of visual attention: namely, sustained attention, attentional selectivity, and attentional capacity. To measure sustained attention, we employed the SART, a simple continuous-performance test that captured fluctuations in sustained attention during a monotonous task. To measure attentional selectivity and capacity, we employed TVA-based assessment, which enabled the estimation of basic attentional parameters relating to visual attentional capacity (perceptual threshold, VSTM capacity, and processing capacity) and the efficiency of top-down attentional selection. There was evidence of age-related decline in each of the measured variables. However, the declines varied markedly in terms of magnitude and lifespan trajectory. The TVA parameters (attentional selectivity and capacity) displayed a linear relationship with age, while the sustained-attention variables were found to have a curvilinear relationship. A comparison of composite effect sizes reflecting age-related decline yielded the largest effect sizes for attentional capacity, followed by sustained attention and attentional selectivity. We found a statistically significant difference between the effect sizes for capacity and selectivity, suggesting that age had a significantly greater impact on capacity than on selectivity. The difference between the effect sizes for attentional capacity and sustained attention approached significance.
The largest declines were seen on the variables related to visual processing capacity (C) and VSTM capacity (K). In the comparison between younger (20s/30s) and older (60s/70s) adults, very large effect sizes were yielded for these two variables. Furthermore, the graphical and regression analyses suggested a linear relationship with age. Both variables had their peak in the teenage years, followed by a linear decline through the succeeding decades. These findings are in keeping with previous evidence of linear decline with age beginning in young adulthood for processing speed (Salthouse, 1996) and VSTM (Lee, Baniqued, Cosman, Mullen, McAuley, Severson & Kramer, 2012; Verhaeghen, Marcoen & Goossens, 1993). We also found a significant increase in perceptual threshold in older relative to younger adults. However, this increase was of a medium effect size, and there was only weak evidence for a linear relationship with age (R 2 = 8.2 %). This finding was surprising, given the marked declines observed on the other variables relating to visual attentional capacity. It is possible that perceptual threshold is simply not as vulnerable to age-related decline. However, it is also possible that the present method of estimating perceptual threshold was not sensitive enough to detect decline across the decades.
The efficiency of top-down attentional selection, α, showed a significant decline from young to older adulthood. This decline was of medium effect size, and there was weak evidence for a linear increase (signifying decline) with age (R 2 = 10.8 %). Again, it is possible that the present paradigm was not sensitive enough to detect greater changes across the lifespan. Alternatively, this finding raises the intriguing possibility that the efficiency of top-down attentional selection is somewhat robust against the effects of age.
All sustained-attention variables displayed declines from younger to older adulthood of medium to large effect sizes. Their lifespan trajectories were best described by a quadratic function: a U-shaped curve with poor performance in the childhood (age 12) and teenage years, a plateau during young to middle adulthood, and a deterioration in performance in older adulthood. Previous studies using vigilance tasks to assess sustained attention have yielded inconsistent results (Davies & Parasuraman, 1982; W. A. Rogers, 2000). One very large-scale study using a vigilance task—the Mackworth Clock Task—failed to find any evidence of sustained-attention deficits in older adults (Giambra & Quilter, 1988). It is possible that the increased sensitivity of the SART in comparison to vigilance tasks enabled it to detect age-related decline in this study. Vigilance tasks typically require a participant to passively observe a stream of stimuli and to respond on the appearance of an infrequent target stimulus. However, the sudden appearance of the infrequent target and the associated response may have an alerting effect that reduces the demands on the endogenous sustained-attention system, and so reduces the sensitivity of the task for the detection of lapses in sustained attention. In the SART these task demands are reversed, so that the participant is required to respond to most stimuli within a stream of stimuli and to withhold response to an infrequent no-go target. It is argued that a task requiring inhibition of a response within the context of an ongoing monotonous action is more taxing upon the endogenous sustained-attention systems and more sensitive for detecting lapses of sustained attention than are traditional vigilance tasks (Robertson et al., 1997). However, one drawback of the SART in the present arena is its reliance upon a speeded motor action. The SART requires a participant to press a button for every stimulus that appears (except for the number 3) within an interstimulus interval of 1,439 ms. This requires quite a fast and coordinated motor response on the part of the participant, and given that motor responses are typically slowed in older adults, it is difficult to conclude for certain whether the sustained-attention deficits seen in the older adults in this study were the result of true lapses in sustained attention or of difficulties with a speeded motor response. A future study could investigate this issue further by using different versions of the SART with varying interstimulus intervals.
The present study was conducted to examine changes in attentional variables across the lifespan and was not designed to test any particular theory of cognitive aging. However, it is noteworthy that the findings are somewhat at odds with the frontal-lobe hypothesis of aging (West, 1996). This hypothesis stated that cognitive processes supported by the prefrontal cortex should manifest decline at an earlier age and of greater magnitude than would cognitive processes supported by nonfrontal regions. In support of this hypothesis are the findings of marked declines in executive functioning in older adults (Kramer et al., 2001; Park, 2000; Park et al., 2001; Reimers & Maylor, 2005; Verhaeghen & Cerella, 2002). In this study, however, the more rudimentary cognitive processes—VSTM and visual processing capacity—showed declines at an earlier age and of greater magnitude than did variables related to the arguably more executive processes of attentional selection and sustained attention. These findings are more compatible with the ideas put forward by Madden and colleagues (Madden, 2007; Madden & Whiting, 2004; Madden, Whiting & Huettel, 2005), who suggested that top-down attentional control is preserved in older adults, and indeed, that older adults rely more on this attentional control in order to compensate for decline in basic sensory and perceptual function. Madden and colleagues described a number of studies of visual search in which older adults were particularly penalised in conditions in which their sensory acuity was challenged (i.e., conditions in which the stimuli were degraded, targets were presented at low exposure durations, or target–distractor similarity was increased), and they contrasted these results with the finding that older adults are able to engage in top-down attentional guidance (i.e., to take advantage of a cue in order to guide visual search). In a functional-imaging study, Madden et al. (2007) showed that during visual search, older adults had greater activation of frontal and parietal cortical areas, traditionally associated with attentional control, than did younger adults, who showed greater activation of occipital areas, related to visual processing. Indeed, this posterior–anterior shift in aging (PASA)—essentially, an age-related reduction in occipitotemporal activity coupled with an age-related increase in frontal activity—has been observed for a variety of cognitive functions and is thought to reflect a compensatory action (Davis, Dennis, Daselaar, Fleck & Cabeza, 2008). The findings, in the present study, of medium to large declines in attentional selection and sustained attention alongside very large declines in aspects of visual attentional capacity (VSTM and processing capacity) are in keeping with the notion that attentional control is relatively preserved in older adults and with the possibility that older adults rely on this attentional control to compensate for decline in other functions.
The study of cognitive aging has revealed that cognitive functions follow different trajectories across the lifespan. In this study, we have shown a clear difference in the trajectories followed by attentional-selectivity, attentional-capacity, and sustained-attention variables. The variables relating to visual attentional capacity (t 0, K, C) and to the efficiency of top-down attentional selection (α) all showed development from age 12 to the teenage years, a peak in the teens or early 20s, and a linear decline thereafter. In contrast, the sustained-attention variables mostly showed a curvilinear relationship with age, and were represented by a U-shaped curve with poor performance in childhood and the teenage years, a plateau during young to middle adulthood, and deterioration in older adulthood. Attention has been characterised as comprising a series of subsystems serving “executive,” “intensity,” and “selectivity” functions (Parasuraman, 1998). It has been argued that these aspects of attention represent independent functions relying upon separate neural systems (McAvinue et al., 2012; Posner & Petersen, 1990). These results provide further evidence for the differentiation of attentional functions, displaying a difference between attentional selectivity, attentional capacity, and sustained attention in terms of the magnitude and lifespan trajectory of age-related decline.
References
Baltes, P. B., & Lindenberger, U. (1997). Emergence of a powerful connection between sensory and cognitive functions across the adult life span: A new window to the study of cognitive aging? Psychology and Aging, 12, 12–21.
Bublak, P., Finke, K., Krummenacher, J., Preger, R., Kyllingsbæk, S., Müller, H. J., & Schneider, W. X. (2005). Usability of a theory of visual attention (TVA) for parameter-based measurement of attention II: Evidence from two patients with frontal or parietal damage. Journal of the International Neuropsychological Society, 11, 843–854.
Bublak, P., Redel, P., & Finke, K. (2006). Spatial and non-spatial attention deficits in neurodegenerative diseases: Assessment based on Bundesen’s theory of visual attention (TVA). Restorative Neurology and Neuroscience, 24, 287–301.
Bublak, P., Redel, P., Sorg, C., Kurz, A., Förstl, H., Müller, H. J., . . . Finke, K. (2009). Staged decline of visual processing capacity in mild cognitive impairment and Alzheimer’s disease. Neurobiology of Aging, Aug 25.
Bundesen, C. (1990). A theory of visual attention. Psychological Review, 97, 523–547. doi:10.1037/0033-295X.97.4.523
Bundesen, C., & Habekost, T. (2008). Principles of visual attention: Linking mind and brain. Oxford, U.K.: Oxford University Press.
Bundesen, C., Habekost, T., & Kyllingsbæk, S. (2005). A neural theory of visual attention: Bridging cognition and neurophysiology. Psychological Review, 112, 291–328. doi:10.1037/0033-295X.112.2.291
Cabeza, R., Nyberg, L., & Park, D. (2005). Cognitive neuroscience of aging: Linking cognitive and cerebral aging. New York, NY: Oxford University Press.
Cohen, J., Cohen, P., West, S. G., & Aiken, L. S. (2003). Applied multiple regression: Correlation analysis for the behavioral sciences. Mahwah, NJ: Erlbaum.
Cooper, H. M. (2010). Research synthesis and meta-analysis: A step-by-step approach. Los Angeles, CA: Sage.
Craik, F. I. M. (2006). Brain-behavior relations across the lifespan: A commentary. Neuroscience and Biobehavioral Reviews, 30, 885–892.
Craik, F. I. M., & Byrd, M. (1982). Aging and cognitive deficits: The role of attentional resources. In F. I. M. Craik & S. Trehub (Eds.), Aging and cognitive processes (pp. 191–211). New York, NY: Plenum Press.
Davies, D. R., & Parasuraman, R. (1982). The psychology of vigilance. London, U.K.: Academic Press.
Davis, S. W., Dennis, N. A., Daselaar, S. M., Fleck, M. S., & Cabeza, R. (2008). Que PASA? The posterior-anterior shift in aging. Cerebral Cortex, 18, 1201–1209.
Dempster, F. N. (1992). The rise and fall of the inhibitory mechanism: Toward a unified theory of cognitive development and aging. Developmental Review, 12, 45–75.
Desimone, R., & Duncan, J. (1995). Neural mechanisms of selective visual attention. Annual Review of Neuroscience, 18, 193–222. doi:10.1146/annurev.ne.18.030195.001205
Dubois, M., Kyllingsbæk, S., Prado, C., Peiffer, E., Lassus, D., & Valdois, S. (2010). Fractionating the multi-character processing deficit in developmental dyslexia: Evidence from two case studies. Cortex, 46, 717–738.
Duncan, J., Bundesen, C., Olson, A., Humphreys, G., Chavda, S., & Shibuya, H. (1999). Systematic analysis of deficits in visual attention. Journal of Experimental Psychology. General, 128, 450–478.
Duncan, J., Bundesen, C., Olson, A., Humphreys, G., Ward, R., Kyllingsbæk, S. M., & Chavda, S. (2003). Dorsal and ventral simultanagnosia. Cognitive Neuropsychology, 20, 675–701.
Dyrholm, M., Kyllingsbæk, S., Espeseth, T., & Bundesen, C. (2011). Generalizing parametric models by introducing trial-by-trial parameter variability: The case of TVA. Journal of Mathematical Psychology, 55, 416–429. doi:10.1016/j.jmp.2011.08.005
Finke, K., Bublak, P., Krummenacher, J., Kyllingsbæk, S., Müller, H. J., & Schneider, W. X. (2005). Usability of a theory of visual attention (TVA) for parameter-based measurement of attention I: Evidence from normal subjects. Journal of the International Neuropsychological Society, 11, 832–842.
Finke, K., Schneider, W. X., Redel, P., Dose, M., Kerkhoff, G., Müller, H. J., & Bublak, P. (2007). The capacity of attention and simultaneous perception of objects: A group study of Huntington’s disease patients. Neuropsychologia, 45, 3272–3284. doi:10.1016/j.neuropsychologia.2007.06.006
Gerlach, C., Marstrand, L., Habekost, T., & Gade, A. (2005). A case of impaired shape integration: Implications for models of visual object processing. Visual Cognition, 12, 1409–1443.
Giambra, L. M., & Quilter, R. E. (1988). Sustained attention in adulthood: A unique, large-sample, longitudinal and multicohort analysis using the Mackworth Clock-Test. Psychology and Aging, 3, 75–83. doi:10.1037/0882-7974.3.1.75
Greenwood, P. M., Parasuraman, R., & Haxby, J. V. (1993). Changes in visuospatial attention over the adult lifespan. Neuropsychologia, 31, 471–485.
Habekost, T., & Bundesen, C. (2003). Patient assessment based on a theory of visual attention (TVA): Subtle deficits after a right frontal-subcortical lesion. Neuropsychologia, 41, 1171–1188.
Habekost, T., & Rostrup, E. (2006). Persisting asymmetries of vision after right side lesions. Neuropsychologia, 44, 876–895.
Habekost, T., & Rostrup, E. (2007). Visual attention capacity after right hemisphere lesions. Neuropsychologia, 45, 1474–1488.
Habekost, T., & Starrfelt, R. (2006). Alexia and quadrant-amblyopia: Reading disability after a minor visual field deficit. Neuropsychologia, 44, 2465–2476.
Habekost, T., & Starrfelt, R. (2009). Visual attention capacity: A review of TVA-based patient studies. Scandinavian Journal of Psychology, 50, 23–32.
Howell, D. C. (2008). Fundamental statistics for the behavioral sciences. Toronto, Canada: Thomson Wadsworth.
Ishihara, S. (1960). Tests for colour-blindness. London: H. K. Lewis.
Johnson, K. A., Kelly, S. P., Bellgrove, M. A., Barry, E., Cox, M., Gill, M., & Robertson, I. H. (2007a). Response variability in attention deficit hyperactivity disorder: Evidence for neuropsychological heterogeneity. Neuropsychologia, 45, 630–638.
Johnson, K. A., Robertson, I. H., Kelly, S. P., Silk, T. J., Barry, E., Dáibhis, A., & Bellgrove, M. A. (2007b). Dissociation in performance of children with ADHD and high-functioning autism on a task of sustained attention. Neuropsychologia, 45, 2234–2245. doi:10.1016/j.neuropsychologia.2007.02.019
Kramer, A. F., Scialfa, C. T., Peterson, M. S., & Irwin, D. E. (2001). Attentional capture, attentional control and aging. Advances in Psychology, 133, 293–322.
Kyllingsbæk, S. (2006). Modeling visual attention. Behavior Research Methods, 38, 123–133. doi:10.3758/BF03192757
Lee, H., Baniqued, P. L., Cosman, J., Mullen, S., McAuley, E., Severson, J., & Kramer, A. F. (2012). Examining cognitive function across the lifespan using a mobile application. Computers in Human Behaviour. doi:10.1016/j.chb.2012.05.013
Madden, D. J. (2007). Aging and visual attention. Current Directions in Psychological Science, 16, 70–74.
Madden, D. J., Spaniol, J., Whiting, W. L., Bucur, B., Provenzale, J. M., Cabeza, R., & Huettel, S. A. (2007). Adult age differences in the functional neuroanatomy of visual attention: A combined fMRI and DTI study. Neurobiology of Aging, 28, 459–476.
Madden, D. J., & Whiting, W. L. (2004). Age-related changes in visual attention. Advances in Cell Aging and Gerontology, 15, 41–88.
Madden, D. J., Whiting, W. L., & Huettel, S. A. (2005). Age-related changes in neural activity during visual perception and attention. In R. Cabeza, L. Nyberg, & D. C. Park (Eds.), Cognitive neuroscience of aging: Linking cognitive and cerebral aging (pp. 157–185). New York, NY: Oxford University Press.
Manly, T., Owen, A. M., McAvinue, L. P., Datta, A., Lewis, G. H., Scott, S. U., & Robertson, I. H. (2003). Enhancing the sensitivity of a sustained attention task to frontal damage: Convergent clinical and functional imaging evidence. Neurocase, 9, 340–349.
McAvinue, L. P., O’Keeffe, F., McMackin, D., & Robertson, I. H. (2005). Impaired sustained attention and error awareness in traumatic brain injury: Implications for insight. Neuropsychological Rehabilitation, 15, 569–587.
McAvinue, L. P., Vangkilde, S., Johnson, K. A., Habekost, T., Kyllingsbæk, S., Robertson, I. H., & Bundesen, C. (2012). The relationship between sustained attention, attentional selectivity, and capacity. Journal of Cognitive Psychology, 24, 313–328. doi:10.1080/20445911.2011.628653
Parasuraman, R. (1998). The attentive brain: Issues and prospects. In R. Parasuraman (Ed.), The attentive brain (pp. 3–15). Cambridge, MA: MIT Press.
Park, D. C. (2000). The basic mechanisms accounting for age-related decline in cognitive function. In D. C. Park & N. Schwarz (Eds.), Cognitive aging: A primer (pp. 3–21). Hove, U.K.: Psychology Press.
Park, D. C., Polk, T. A., Mikels, J. A., Taylor, S. F., & Marshuetz, C. (2001). Cerebral aging: Integration of brain and behavioral models of cognitive function. Dialogues in Clinical Neuroscience, 3, 151–165.
Pashler, H. (1994). Dual-task interference in simple tasks: Data and theory. Psychological Bulletin, 116, 220–244. doi:10.1037/0033-2909.116.2.220
Peers, P. V., Ludwig, C. J., Rorden, C., Cusack, R., Bonfiglioli, C., Bundesen, C., & Duncan, J. (2005). Attentional functions of parietal and frontal cortex. Cerebral Cortex, 15, 1469–1484. doi:10.1093/cercor/bhi029
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology, 32, 3–25. doi:10.1080/00335558008248231
Posner, M. I., & Petersen, S. E. (1990). The attention system of the human brain. Annual Review of Neuroscience, 13, 25–42.
Raz, N. (2004). The aging brain observed in vivo: differential changes and their modifiers. In R. Cabeza, L. Nyberg, & D. C. Park (Eds.), Cognitive neuroscience of aging: Linking cognitive and cerebral aging (pp. 17–55). New York, NY: Oxford University Press.
Raz, A., & Buhle, J. (2006). Typologies of attentional networks. Nature Reviews Neuroscience, 7, 367–379.
Raz, N., & Rodrigue, K. M. (2006). Differential aging of the brain: Patterns, cognitive correlates and modifiers. Neuroscience and Biobehavioral Reviews, 30, 730–748.
Redel, P., Bublak, P., Sorg, C., Kurz, A., Förstl, H., Müller, H. J., . . . Finke, K. (2012). Deficits of spatial and task-related attentional selection in mild cognitive impairment and Alzheimer’s disease. Neurobiology of Aging, 33, 195.e27–195.e42. doi:10.1016/j.neurobiolaging.2010.05.014
Reimers, S., & Maylor, E. A. (2005). Task switching across the life span: Effects of age on general and specific switch costs. Developmental Psychology, 41, 661–671.
Robertson, I. H., Manly, T., Andrade, J., Baddeley, B. T., & Yiend, J. (1997). “Oops!”: Performance correlates of everyday attentional failures in traumatic brain injured and normal subjects. Neuropsychologia, 35, 747–758.
Rogers, R. D., & Monsell, S. (1995). Costs of a predictable switch between simple cognitive tasks. Journal of Experimental Psychology. General, 124, 207–231.
Rogers, W. A. (2000). Attention and aging. In D. C. Park & N. Schwarz (Eds.), Cognitive aging: A primer (pp. 57–73). Hove, U.K.: Psychology Press.
Salthouse, T. A. (1996). The processing-speed theory of adult age differences in cognition. Psychological Review, 103, 403–428. doi:10.1037/0033-295X.103.3.403
Sheridan, M. D. (1970). New appliances: Sheridan–Gardiner test for visual-acuity. British Medical Journal, 1, 108–109.
Smilek, D., Carriere, J. S. A., & Cheyne, J. A. (2010). Failures of sustained attention in life, lab, and brain: Ecological validity of the SART. Neuropsychologia, 48, 2564–2570.
Starrfelt, R., Habekost, T., & Gerlach, C. (2010). Visual processing in pure alexia: A case study. Cortex, 46, 242–255.
Starrfelt, R., Habekost, T., & Leff, A. P. (2009). Too little, too late: Reduced visual span and speed characterize pure alexia. Cerebral Cortex, 19, 2880–2890.
Sturm, W. (1996). Evaluation in therapeutical contexts: Attentional and neglect disorders. European Review of Applied Psychology, 46, 207–215.
Treisman, A. M., & Gelade, G. (1980). A feature-integration theory of attention. Cognitive Psychology, 12, 97–136. doi:10.1016/0010-0285(80)90005-5
Trick, L. M., & Enns, J. T. (1998). Lifespan changes in attention: The visual search task. Cognitive Development, 13, 369–386.
Vangkilde, S., Coull, J. T., & Bundesen, C. (2012). Great expectations: Temporal expectation modulates perceptual processing speed. Journal of Experimental Psychology. Human Perception and Performance. doi:10.1037/a0026343
VanZomeren, A. H., & Brouwer, W. H. (1994). Clinical neuropsychology of attention. New York, NY: Oxford University Press.
Verhaeghen, P., & Cerella, J. (2002). Aging, executive control, and attention: A review of meta-analyses. Neuroscience and Biobehavioral Reviews, 26, 849–857.
Verhaeghen, P., Marcoen, A., & Goossens, L. (1993). Facts and fiction about memory aging: A quantitative integration of research findings. Journal of Gerontology, 48, P157–P171.
West, R. L. (1996). An application of prefrontal cortex function theory to cognitive aging. Psychological Bulletin, 120, 272–292.
Zacks, R., & Hasher, L. (1997). Cognitive gerontology and attentional inhibition: A reply to Burke and McDowd. Journals of Gerontology, 52B, P274–P283.
Author note
Thank you to the Irish Research Council for the Humanities & Social Sciences and the European Science Foundation for supporting this research.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
McAvinue, L.P., Habekost, T., Johnson, K.A. et al. Sustained attention, attentional selectivity, and attentional capacity across the lifespan. Atten Percept Psychophys 74, 1570–1582 (2012). https://doi.org/10.3758/s13414-012-0352-6
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-012-0352-6