









-
Epichloë fungal endophytes, belonging to the family Clavicipitaceae, have coevolved to form symbiotic relationships with grasses in the Pooidae sub-family[1]. Asexual Epichloë grow systemically in the above-ground parts of the plant, infecting the axillary buds from which new tillers develop, with the intercellular hyphae primarily confined to the leaf sheath. They have no external stage and are transmitted maternally via the host seed. The fungus is wholly dependent on its host for nourishment, protection, and a means of propagation. A diversity of strains exist in native habitats of hosts, distinguishable by genetic differences[2] and the varied array of secondary metabolites they produce[3]. The effects of these alkaloids on vertebrate and invertebrate herbivores are important factors in determining the outcome of the symbiosis for the host plant. Over 40 invertebrate species are adversely affected by the presence of Epichloë endophytes in perennial ryegrass (Lolium perenne), tall fescue (Festuca arundinacea) and meadow fescue (Festuca pratensis)[4−6]. There is also evidence that these endophytes confer tolerance to abiotic stress[7]. These mutual advantages have been a driving force in the evolution of asexual reproduction in Epichloë species and the reason for the association being generally considered to be a defensive mutualism[8,9]. There are, however, dissenting views regarding the status of the relationship with authors making a case for effects ranging from parasitism to mutualism particularly in native grasses[10−12]. A recent paper by Newman et al.[13] has sought to clarify this by presenting mathematical evidence that those Epichloë that are strictly vertically transmitted (about 50% of this genus) can only exist as mutualists.
The symbiosis has been exploited in agriculture in the United States, Australia and New Zealand in both turf and pasture grasses where protection from biotic and abiotic stress enhances plant growth and persistence[14−17]. The effects they have on insect pests is particularly important in New Zealand where both native and introduced species cause annual economic losses in agriculture of between NZ
$\$ $ 1.7B and$ \$$ 2.3B[18]. After the mammalian toxicity of the common toxic (CT) strain in L. perenne was recognised in the 1980s due to its production of lolitrem B and ergovaline, novel strains of E. festucae var. lolii were sought that did not produce these alkaloids. Two strains, AR1 and AR37, met these criteria and are now widely used in New Zealand pastoral agriculture. AR1 produces a pyrrolopyrazine compound, peramine, whereas AR37 produces a group of indole diterpenoids known as epoxyjanthitrems. Peramine produced by AR1 is a powerful deterrent to Argentine stem weevil (Listronotus bonariensis)[19], a major pest of ryegrass, and this endophyte also protects against pasture mealybug (Balanococcus poae)[20]. AR37 provides protection against five New Zealand pasture pests including both Argentine stem weevil and pasture mealybug[16,21]. Latch et al.[22] were the first to document increased growth of L. perenne that was attributed to endophyte infection increasing root growth. Some studies have demonstrated increases in growth and longevity of endophyte-infected relative to endophyte-free L. perenne due to increased abiotic stress tolerance[23,24] whereas others have found negative or negligible effects[25−27] or genotype-specific effects that, in some cases, were related to climatic factors in the areas where the plants originated[28−30]. Reported increases in growth due to endophyte, particularly as they relate to water stress, are more common in tall fescue[17,31−35] but an explanation for such effects often remains elusive. Surveys for endophyte infection among grasses have suggested that the ryegrass-endophyte associations are more likely to occur in drier areas[23,36,37] although this may not be true for native grass associations or under extreme drought conditions[38]. There is also evidence that interactions between herbivory and drought has more severe consequences than either stress alone with greater plant growth and survival where endophyte alleviates the herbivory[6,39−41].There is good evidence that endophyte reduces invertebrate herbivory that improves the growth and survival of L. perenne which results in increased endophyte infection levels in the field[21,41−44]. Few studies, however, have investigated the direct consequences of endophyte-mediated reductions in insect attack for individual plant growth and survival of ryegrass. In this study a population of genetically related perennial ryegrass plants without endophyte or infected with the common toxic (CT), AR1 or AR37 strains of E. festucae var. lolii, were exposed to natural insect infestations in an outdoor container trial. Confounding effects of plant genotype were minimised by accessing all plants from the same breeding line and applying insecticide to a clonal set of plants for each endophyte treatment so that the impact of different endophyte strains on insect populations and their effects on ryegrass growth could be determined. Comparisons between insecticide-treated (TR) and untreated (UN) plants were made of foliar and root growth, tiller and plant mortality and insect populations. Detailed data on the densities of A. lentisci on untreated plants are given in Popay and Cox[45]. Based on previous studies, it was hypothesised that the outcomes of the symbiosis for the plant would vary with endophyte haplotype and be governed primarily by differences in populations of insect herbivores. Results are interpreted in the context of costs and benefits of the symbiosis for the host.
-
For individual samples, foliar growth of UN AR37 plants exceeded that of the other UN endophyte treatments in late winter Year 1 (> AR1 and CT) and in autumn and spring of Year 2 (> AR1, CT and Nil) (p < 0.001) (Fig 1a−d; Supplemental Fig. S1a). In spring of Year 2, growth of UN AR1 and CT was also greater than that of UN Nil (p < 0.05). Insecticide did not increase foliar growth of AR37 for any Sample time (Fig. 1b). In contrast to this, foliar growth of TR AR1, CT and Nil was significantly greater than that of their UN counterparts in Samples 3−5 (autumn Year 2 to summer Year 3) (Figs 1a, c). Insecticide treatment also increased foliar growth of AR1 in the summer of the first year (Sample 2) (p = 0.008). The greatest growth increases relative to UN, were 73%, 56% and 30% for AR1, Nil and CT respectively in autumn of Year 2 (Sample 3), and 47%, 130% and 24% for the same treatments in spring, Year 2 (Supplemental Fig. S1a).
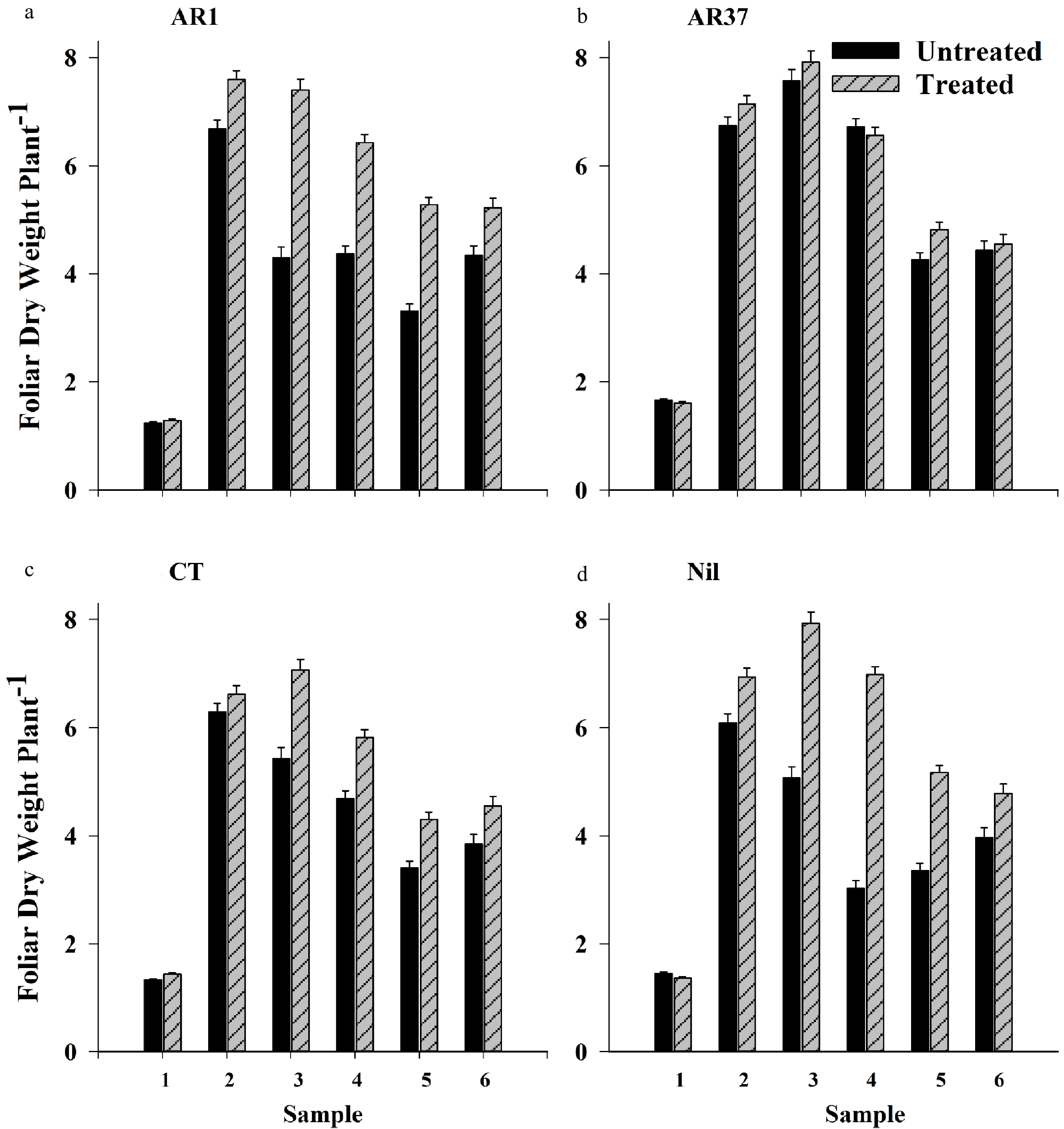
Figure 1. Foliar growth (g plant−1 +SE) of ryegrass infected with endophytes (a) AR1, (b) AR37, (c) CT, or endophyte-free (d) Nil and treated or not treated with insecticide, for six Samples taken over 18 months; 1 – late winter Year 1; 2 – Summer Year 1; 3 – Autumn Year 2; 4 – Spring Year 2; 5 – Summer Year 3; 6 – Autumn Year 3.
There were fewer differences in root outgrowth than in foliar growth among UN plants. Nil outgrowth was greater than AR1 in autumn Year 2 (Sample 3), and root outgrowth of AR37 was greater than AR1 and Nil in spring Year 2 (Sample 4) (Supplemental Fig. S1b; Fig 2a, b, d). Insecticide increased root outgrowth of AR37 only in summer of Year 3 (Sample 5) but increased outgrowth of AR1 and CT for Sample 2, and AR1, CT and Nil for Samples 3, 4 and 5 (Supplemental Fig S1b; Fig 2a–d). Percentage increases due to insecticide were greatest in autumn and spring in Year 2 with AR1 gaining 233%, Nil 40% and CT 92% more roots between early summer in Year 1 and autumn in Year 2 (Fig. S1b). In early spring of Year 2 the outgrowth since the autumn sampling showed increases in roots due to insecticide treatment of 55%, 99% and 32% for AR1, Nil and CT respectively relative to their untreated equivalents.
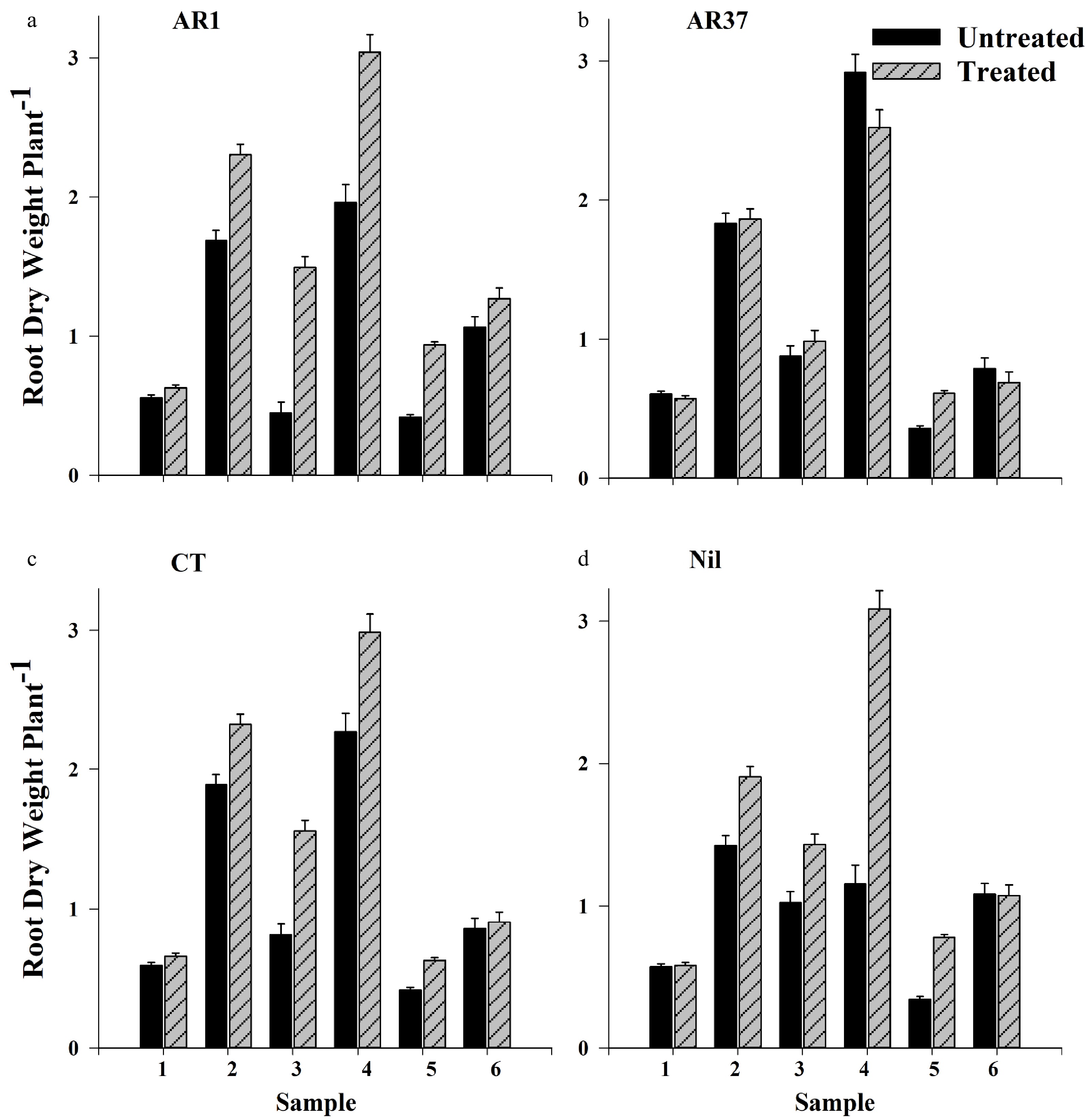
Figure 2. Root outgrowth (g plant−1 +SE) of ryegrass infected with endophytes (a) AR1, (b) AR37, (c) CT, or endophyte-free (d) Nil, and treated or not treated with insecticide, for six Samples taken over 18 months: 1 – late winter Year 1; 2 – Summer Year 1; 3 – Autumn Year 2; 4 – Spring Year 2; 5 – Summer Year 3; 6 – Autumn Year 3.
Cumulative foliar growth on UN plants was significantly higher for plants infected with AR37 than for other endophyte treatments for all Samples except 1 and 2 (Fig. 3a). There were no significant differences between the other three endophyte treatments. Root outgrowth of UN AR37 was significantly higher than UN Nil at Samples 4, 5 and 6 (Fig. 3b). Compared with their UN counterparts, TR plants infected with AR1 and CT accumulated significantly more root outgrowth for Samples 3–6, and Nil for Samples 4–6. Cumulative root outgrowth for TR AR37 did not differ from UN AR37 but was significantly less than TR AR1 Samples 3 to 6 and less than TR CT at Samples 3 and 4.
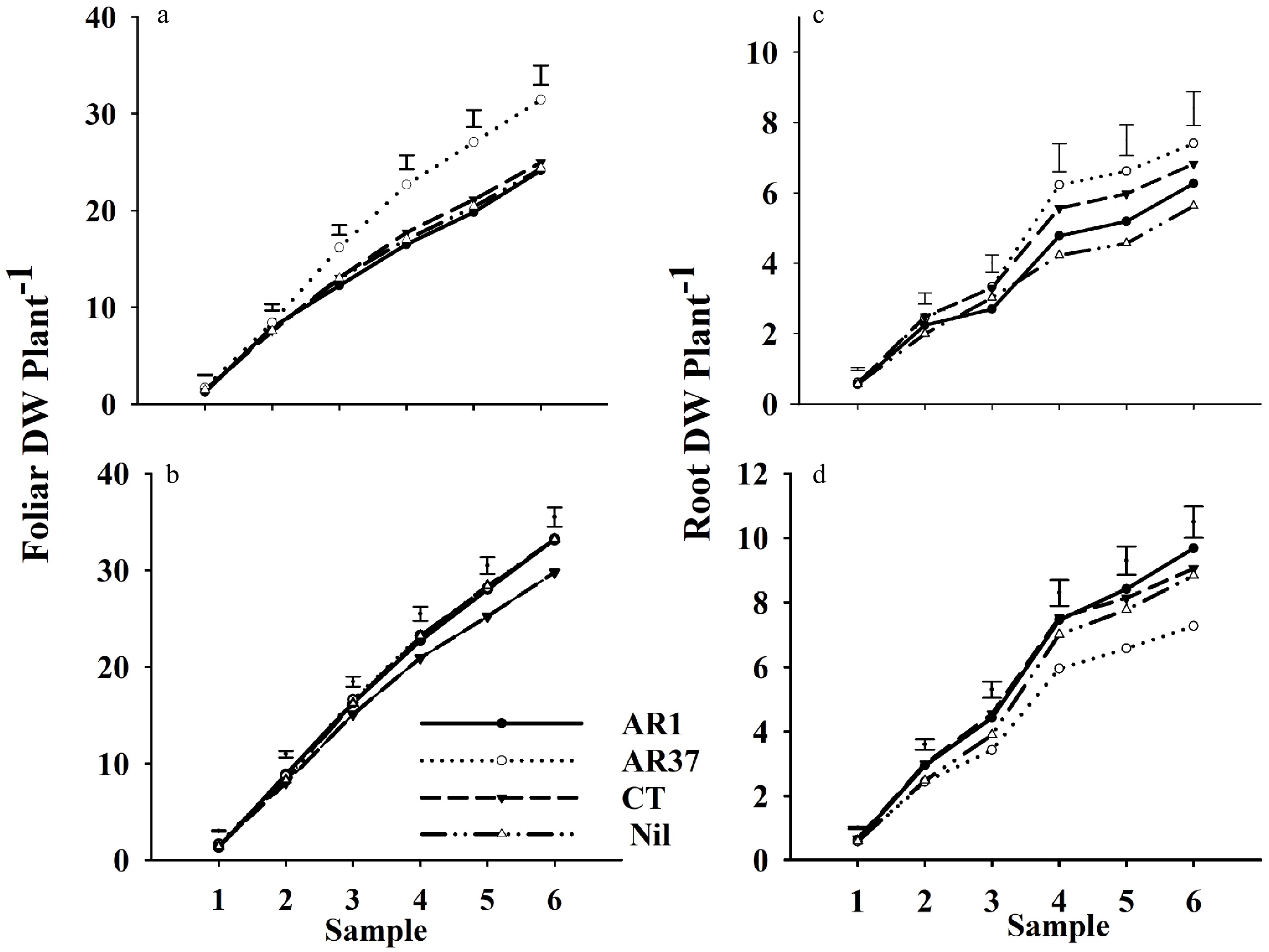
Figure 3. Cumulative dry weight of ryegrass infected with endophytes (a) AR1, (b) AR37, (c) CT, or endophyte-free (d) Nil for foliar growth of plants untreated or treated with insecticide (a, c), and for root outgrowth of plants untreated and treated (b, d) for six Samples taken over 18 months: 1 – late winter Year 1; 2 – Summer Year 1; 3 – Autumn Year 2; 4 – Spring Year 2; 5 – Summer Year 3; 6 – Autumn Year 3. Error bars = SED for endophyte by treatment comparisons.
Root:shoot ratios in UN plants were significantly less in AR1 than in Nil in autumn Year 2 (Sample 3); similarly, this ratio was less in UN Nil compared with CT in spring Year 2 (Sample 4) but did not differ significantly between endophyte treatments at other sampling times (Table 1). The overall root:shoot ratio calculated for the combined totals of root outgrowth and foliar growth were also not significantly different (Table 1). Insecticide increased root:shoot ratios of AR1–infected ryegrass in summer Year 1, autumn Year 2 and summer Year 3 (Table 1). In contrast to this, differences between UN and TR plants were significant for CT only in autumn Year 2, and for Nil in summer Year 1, but did not differ at any sampling date for AR37 treatments. AR37 had the lowest root:shoot ratio among the endophyte treatments in TR plants. This was significantly less than the ratio for CT and AR1 in autumn Year 2 and also less than the ratio for these two treatments for the mean total growth (Table 1).
Table 1. Root:shoot ratios of ryegrass without endophyte or infected with AR1, AR37 or CT endophytes for plants treated (TR) and untreated (UN) with insecticide. Ratios are calculated from root outgrowth and foliar growth at a 50 mm cutting height measured for six Samples taken over 18 months.
Sample1 AR1 AR37 CT Nil LSD2 UN TR UN TR UN TR UN TR (5%) 1 0.47 0.49 0.38 0.39 0.47 0.46 0.38 0.42 0.104 2 0.25 0.31 0.30 0.27 0.30 0.36 0.23 0.30 0.088 3 0.08 0.20 0.12 0.12 0.13 0.21 0.15 0.17 0.055 4 0.46 0.48 0.44 0.39 0.50 0.52 0.35 0.45 0.135 5 0.12 0.19 0.09 0.12 0.11 0.15 0.10 0.14 0.051 6 0.24 0.24 0.18 0.16 0.22 0.22 0.24 0.20 0.087 Mean 0.26 0.30 0.24 0.22 0.22 0.26 0.27 0.31 0.06 1 1 – Winter Year 1; 2 – Summer Year 1; 3 – Autumn Year 2; 4 – Spring Year 2; 5 – Summer Year 3; 6 – Autumn Year 3.
2 LSD is for comparison between endophyte treatments within each insecticide stratum and not for comparisons between UN and TR plants within each endophyte treatment.
Bold lettering indicates a significant difference (p < 0.05) between treated and untreated plants within each endophyte treatment.At the completion of the trial in autumn Year 3 (Sample 6), the dry weight of leaf sheaths (< 50 mm) on plants did not differ significantly between UN endophyte treatments but was significantly greater on TR AR1 and Nil plants compared with their UN counterparts (Table 2). Weight of main plant roots in the inner bag was similar among UN plants, but in a significant interaction with endophyte (p = 0.009), insecticide increased root weight for AR1, CT and Nil but not AR37.
Table 2. Back-transformed percentage (±SE) of dead tillers for UN L. perenne plants without endophyte (Nil) or infected with AR1, AR37 or CT endophytes, and dry weight of leaf sheath and main plant roots in the inner bag for TR and UN plants at the final harvest in autumn 2004.
Endophyte Leaf sheath (g) Roots (g) % Dead tillers UN TR UN TR Summer Y1 Autumn Y2 Spring Y2 AR1 2.35 3.08 6.94 13.37 2.5 ± 0.8 10.6 ± 4.2 18.7 ± 8.3 AR37 2.59 2.54 8.78 9.26 0.7 ± 0.1 2.1 ± 0.9 3.9 ± 2.1 CT 1.95 2.33 7.52 10.14 1.1 ± 0.3 4.3 ± 1.8 5.4 ± 2.8 Nil 2.16 2.80 6.69 10.54 1.6 ± 0.2 16.4 ± 6.0 43.8 ± 13.4 LSD1 0.4621 2.426 P2 0.001 0.009 0.050 < 0.001 0.001 1 LSD is for comparisons between UN and TR for the same endophyte; bold lettering indicates a significant difference.
2 P value is for differences in percentage of dead tillers between endophyte treatments.
Bold lettering indicates a significant difference (p < 0.05) between treated and untreated plants within each endophyte treatment.Reproduction and tiller mortality
-
The percentage of tillers that were reproductive in the first summer (mean 14%) was similar for the different endophyte treatments and between TR and UN plants (data not presented). A year later only a small proportion of tillers were reproductive (< 5%) with AR37 having a significantly lower (p = 0.005) percentage of reproductive tillers than the other endophyte treatments (AR37 0.8% cf. 2.7%, 3.4%, 3.4% for AR1, CT and Nil respectively, back transformed mean of total for UN and TR). Insecticide significantly (p < 0.001) increased the percentage of reproductive tillers (UN 1.6%; TR 3.1%) at this time with only a slight increase between UN and TR AR37 (0.4%) compared with a larger increase for the other endophyte treatments (average 2.9%).
Tiller mortality was low in the summer of Year 1 but increased considerably in UN AR1 and Nil plants in Year 2 and for all treatments in Year 3. Fewer tillers died on UN AR37 and CT plants than on UN AR1 and Nil plants in autumn and spring of Year 2 (p < 0.001) (Table 2). On both of these sampling occasions, AR1 had fewer dead tillers than Nil (p < 0.05). Tiller mortality was significantly reduced on TR plants compared with UN (p < 0.001) for the three sampling occasions in Table 2 and did not differ significantly between endophyte treatments (data not presented). In Samples 5 and 6 (summer and autumn Year 3), tiller mortality increased on all plants and there was no difference between endophyte treatments or between UN and TR plants (data not presented). Among UN plants, 20% of AR1 and 30% of Nil plants died after the autumn Year 2 sampling, whereas there was no mortality of AR37 or CT-infected plants, or of any TR plants.
Hyphal density
-
Densities of fungal hyphae in the leaf sheaths of AR37, CT and AR1 were, respectively, 13.8, 8.9 and 5.9 mm−1 breadth of leaf sheath (d.f. 68, SED 1.35). There was no effect of insecticide treatment on mean hyphal concentration and no correlation between cloned pairs of TR and UN plants for each endophyte strain (data not shown).
Insect infestations
-
The major pest present throughout the trial was the root aphid A. lentisci. Here we present the back-transformed aphid loadings (mean number of A. lentisci g−1 of root) on UN and TR plants. Further data pertaining to A. lentisci populations on UN plants in the trial are reported in Popay and Cox[45]. This aphid occurred in very low numbers on UN AR37-infected plants in all samplings (Supplemental Table S1). The aphid loadings did not differ significantly between AR37 and CT in spring of Year 2, and between AR37 and AR1 in summer Year 3 but for all other comparisons, AR37 had significantly (p < 0.001) lower aphid loadings than the other endophyte treatments. The highest aphid loading (arithmetic mean of 962 aphids g−1) occurred on root outgrowth of AR1 plants in autumn Year 2 (Sample 3). Analysis of log transformed data showed this was significantly greater than the loading on Nil and CT (p < 0.05) and CT had a lower loading than Nil (p < 0.05)[45]. Aphid loadings were also high on both AR1 and Nil plants in the following spring (Sample 4). Aphid loadings on the main plant roots in the inner bag at the final sampling (Autumn B in Supplemental Table S1) were low relative to the loading on outgrowth. Insecticide reduced root aphid numbers g−1 of root on AR1, CT and Nil plants (p < 0.001) but not on AR37 for each Sample (Supplemental Table S1).
In addition to root aphid, leaf sheaths of UN AR1 and Nil plants became infested by the mealybug P. graminicola in winter and summer of Year 1 which persisted and became severe by autumn of Year 2 (Supplemental Table S2). The percentage of tillers with this mealybug was significantly lower on AR37 and CT endophyte-infected plants than on AR1 and Nil. Insecticide treatment in early winter Year 1 had little effect on mealybug occurrence in the late winter sample of that year but virtually eliminated these insects from TR plants thereafter (data not shown). Minor black beetle (Heteronychus arator) adult damage to tillers was also recorded in 20% of UN AR1 plants and 15% of UN Nil plants in early summer Year 1. An unidentified species of sod webworm (Crambidae:Lepidoptera) became a problem in all plants, regardless of endophyte status or insecticide treatment, in Year 3.
-
Many studies have shown that fungal endosymbionts of grasses negatively affect insects[5] but relatively few have demonstrated that the associated reduced herbivory advantages the plant. Yet if we are to accept that the asexual biotrophic fungal endophytes of grasses function primarily as defensive mutualists then we must also determine that protection conferred on the host increases aspects of its fitness, such as its growth, competitive ability, survival and ability to reproduce. 'To unequivocally demonstrate the impact of herbivores on plants, whether negative or positive, requires that the grazed plants exhibit a significant change in fitness relative to ungrazed controls'[46]. Here, the fitness of a population of ungrazed perennial ryegrass plants has increased relative to the grazed due to resistance to two insect herbivores, A. lentisci and P. graminicola, imparted by haplotype-specific effects of Epichloë endosymbionts. With the lowest levels of herbivory, L. perenne infected with the AR37 endophyte had the highest shoot and root growth compared with those plants without endophyte or infected with CT and AR1. Furthermore, and in contrast to the other endophyte treatments, foliar growth of plants infected with AR37 was not increased at any time by insecticide application and root growth was increased only once. Representing an ungrazed control, AR37-infected plants therefore provided a benchmark against which plant growth and survival of grazed AR1 and Nil were compared and found to be considerably reduced. The genetically identical set of AR1 and Nil plants treated with insecticide also represented ungrazed controls that provided clear evidence that insects feeding on their grazed counterparts decreased growth and survival. Fitness of plants infected with the CT strain was less compromised compared to AR1 and Nil associations with reduced plant growth relative to TR isogenic plants and, at times, also relative to growth of UN AR37, but no plant mortality.
The ability of any particular endophyte strain to benefit its host by reducing herbivory is highly dependent on the effects of the endophyte on particular insect herbivores, the abundance of the lifecycle stage that reduces host fitness and the plant part damaged by herbivory. Optimal Defence Theory[47] predicts that resources for defence are allocated to the tissues that are of most value for plant survival and fitness. Thus, as mechanisms of defence, Epichloë endosymbionts will deliver the greatest benefit to the host for those insects that threaten plant survival by attacking the meristematic tissue, roots, or sequestering nutrients required for plant growth. It is the latter that occurred in this experiment due to the presence of two sap-sucking insects, A. lentisci feeding on root phloem and P. graminicola, a mealybug that lives within the tillers near the base of plants. These insects attained their maximum populations in autumn and spring in year 2, and this was reflected in large differences in plant growth and tiller mortality. As vascular parasites, these insects can manipulate their host physiology to attract assimilates to their feeding sites[48] with major consequences for infested plants especially if these insects maintain a chronic presence as is the case for A. lentisci[41]. AR37 almost completely suppressed populations of A. lentisci with an average infestation measured on UN root outgrowth of 10 aphids g−1 of root compared with 350 g−1 on AR1, 143 g−1 on CT and 156 g−1 on Nil. An infestation by P. graminicola in autumn of Year 2 coincided with high aphid numbers on Nil and AR1. The combined effect of both insects is likely to have caused the plant mortality in these endophyte treatments. Both these insects were effectively eliminated by the insecticide resulting in marked increases in plant growth and survival of TR plants such that these parameters did not differ from those of AR37-infected plants. The impact of A. lentisci alone in reducing the growth of plants infected with the CT strain, compared with TR isogenic plants and plants infected with AR37, substantiates other reports that this aphid reduces productivity of ryegrass in the field[21,41,49].
Allocation of biomass to different plant parts, although considered to be a highly conserved trait among plant species, may be modified by symbiotic organisms[30,50−53]. Increased root growth of UN Nil relative to UN AR1 in autumn of Year 2 and the greater responses in root:shoot ratios of AR1-infected plants to insecticide treatment are indicative of this. Likewise, cumulative root growth and root:shoot ratios were significantly less in TR AR37 plants than TR AR1 and CT plants. Insecticide use in our trial also demonstrated differences in resource allocation to reproductive effort. These results suggest that the endophyte may represent a cost to the plants that manifests as an altered allocation of resources to root growth and reproduction depending on environmental pressures. A meta-analysis undertaken by Omacini et al.[54] also found that root biomass was reduced on average by 9% in endophyte-infected grasses compared with endophyte-free, positing that differences in root architecture might be responsible. Similarly, Hesse et al.[29] reported a genotype-specific increase in seed production, root growth and root:shoot ratios. Notably, that genotype was sourced from a dry area whereas similar effects were not apparent in the genotype originating from a wet site providing evidence that plant genotype-endophyte associations adapt to maximise their benefits.
Biotrophic organisms like Epichloë are expected to impose a cost on their hosts that is less than the benefit the plant gains from infection if the endophyte is to persist in populations[10,55]. Fungal biomass relative to the whole plant biomass is low but proportional to the production of alkaloids[56−58], although plant genotype and tissue location also correlate with alkaloid concentrations[59]. Alkaloid production extracts a metabolic cost from the host in addition to that required for sustenance of fungal growth and maintenance alone. Differences in hyphal density found in this study are relevant to this. Hyphal density in leaf sheaths of AR37-infected plants was 1.6 and 2.3 times higher than in plants infected with the CT and AR1 haplotypes respectively. These differences in hyphal mass potentially equate to differences in host benefits; that is AR37 > CT > AR1 for differential effects on A. lentisci and P. graminicola. A model describing the relationship between extent of mycorrhizal colonisation of a plant and the benefit that the plant derives from that provides an analogous situation[60]. The model proposed is curvilinear where the plant receives maximum benefit at moderate mycorrhizal densities where benefits outweigh the costs. The intermediate hyphal density of the CT strain fits with this scenario, affording its host with some protection from herbivory at no apparent cost to the plant. The outcomes for the host-endophyte associations in this experiment also fit with the model described by Rudgers et al.[61] in which endophyte effects on survival and regeneration may be opposing forces over the life cycle of the host but can still lead to high infection frequency.
Newman et al.[13] emphasised that costs and benefits of endophyte (or disadvantages and advantages) must be considered over the lifetime of the individual plant. Most experimental evidence relating to this, however, is based on short term experiments and ours is no exception. Nevertheless, in lieu of the difficulty of gathering data throughout the lifetime of a potentially long-lived perennial species, we believe our experiment provides sufficient evidence on which to predict lifetime effects in the presence of similar herbivores. Thus, given the high survival and better growth of plants infected with AR37 exposed to A. lentisci and P. graminicola, we would expect lifetime fitness of ryegrass would vary with presence/absence of endophyte and endophyte haplotype such that AR37 > CT > AR1 > Nil. Evidence from the field supports this conclusion. For example, in a field trial in Australia, productivity of ryegrass with AR37 exceeded that of CT, AR1 and Nil under high populations of A. lentisci with CT having better growth than AR1 and Nil[41] These differences persisted until the trial was terminated the following year when persistence scores, although not significantly different, suggested that plant survival was also compromised and reflected the differences in growth. Thom et al.[21] reported similar results from a New Zealand trial affected by a range of pests in which tiller density, used as a proxy for fitness, was highest in AR37-infected ryegrass, significantly greater than CT and AR1 with Nil having the lowest. Both field trials were affected by drought which in conjunction with the pest damage would have increased the stress on plants. Greater root growth due to reduced herbivory as found here and altered root distribution[62,63] will play an important role in the ability of plants to access water under drought stress, yet drought stress trials have often been conducted without any cognisance of such interactions.
This experiment has illustrated that the functional significance of the asexual endophytes in managed ecosystems is highly dependent on the haplotype (and the alkaloids they produce), the interactions within the host-symbiont association and with the temporal and spatial challenges within its environment whether they be biotic or abiotic or both at once. Two haplotypes, AR37 and CT, have benefitted their ryegrass hosts with resistance to two vascular pests, A. lentisci and P. graminicola. Conversely, infection with AR1, with no effects on these pests, has occasionally imposed a greater cost on plants hosting this endophyte than on plants without an endosymbiont. Under these circumstances the cost of the relationship outweighs the benefits. Exposed to attack by L. bonariensis or B. poae, however, there are clear advantages for growth and survival of AR1-infected L. perenne over endophyte-free[20,21,42] exemplifying a mutualism. Such conflicting outcomes illustrate the complexities of classifying the endosymbiostic relationship as only mutualistic, based on short term events whether they are in agricultural or native ecosystems. Ultimately this is determined by the sum of pressures to which each association is exposed in any particular environment throughout its life. The origins of differences between haplotypes almost certainly lie within the ecosystems they evolved in and thus we cannot expect to see the same costs and benefits when the association is planted outside those places.
-
To obtain seed infected with the different endophyte strains, sub-populations of at least 20 young endophyte-free seedlings from an AgResearch L. perenne breeding line 'GA66' were inoculated with cultured fungi of AR1 and AR37 and seed harvested from each infected plant. Seed cv. GA66 infected with the CT strain was obtained from the Margot Forde Germplasm Centre, AgResearch, Palmerston North, New Zealand. Seed for each of the four ryegrass/endophyte combinations (CT, AR1, AR37 and an endophyte-free (Nil) control) was germinated on damp filter paper before being planted individually into pots (120 mm diam.) in spring. The growing medium consisted of two parts of an unsterilised silt loam field soil mixed thoroughly with one part of washed river sand (v/v) with 1 g of Osmocote® slow-release fertiliser incorporated into the top 50 mm. From 3 months of age, plants were given 30 mL of a nutrient solution (Thrive™) with additional N as required, at each defoliation. Plants were initially maintained in an open air screenhouse under regular automatic overhead watering, with trimming as necessary to maintain vegetative growth.
At 6 months old (autumn Year 1), plants were cloned by splitting into two ramets of six tillers each. Cloned pairs of 20 plants for each endophyte status were then planted individually into a fresh soil/sand growing medium in polythene planter bags (90 × 90 × 200 mm). To enable root growth to be measured periodically without disturbing the plant, additional pairs of holes (5 mm diam., 25 mm apart) were made in each planter bag at 30 mm, 70 mm and 110 mm from the top of the planter bag. The small planter bag was placed inside a larger one (160 × 160 × 370 mm), with the space between the two bags filled with sand. This method was a modification of the method described by Lund et al.[64] as the 'implanted soil mass technique' or the ingrowth technique[65] with root outgrowth measured by severing roots from the plant where they exited the smaller planter bag.
Each replicate consisting of eight plants (one cloned pair for each of the four endophyte treatments) was set up in a split-plot design (insecticide-treated (TR) or not treated (UN)) in black plastic tubs, internal dimensions of 485 × 875 mm with a depth of 300 mm. Each tub was divided in two by a plastic barrier to prevent lateral leaching of insecticide, and one plant of each cloned pair was assigned to each side and arranged randomly within the sub-plot. The planter bags were placed on a sand base so that the herbage grew above the tub, and the bags were surrounded by sand. Insecticide was applied to one of each cloned pair of plants to reduce herbivory. Confidur® containing 5% imidacloprid was mixed with sand and applied to the soil surface at the rate of 80–100 mg of granules per plant. The first application was made in early winter, 6 weeks after the trial was set up and thereafter following each sampling occasion.
The experiment was set up outside at the Ruakura Research Centre, Hamilton, New Zealand. Irrigation was applied only as needed to prevent the plants from wilting and dying during prolonged dry weather. Plants were watered with a hand-held hose for 4 seconds/plant, or with a sprinkler for 2-h periods, which was sufficient to wet the sand in the outer bag and moisten the soil in the inner bag. A nutrient solution (Thrive™, prepared at the recommended rate (approximately 8 g per 4.5 L of tap water) and additional nitrogen (5 g urea per 4.5 L) was applied at the rate of 70 mL plant−1 immediately after each sampling and thereafter at least monthly with additional applications in spring and autumn during periods of rapid growth.
Assessment of endophyte infection
-
The endophyte infection status of plants was determined using an immunoblot procedure[66]. Plants were tested at 2 months old so that only plants with the appropriate endophyte status were used in the trial. The endophyte status of all plants was checked again 18 months after the experiment was set up. Both AR37 plants in Rep 2 and one plant in Rep 13 were found to have lost their endophyte and were therefore excluded from all analyses. All other plants were found to have the appropriate endophyte status.
At the end of the trial in autumn Year 3, hyphal density was determined on three tillers taken from each plant using the method described in di Menna and Waller[67]. Hyphal counts were made across the breadth of the leaf sheath as close as possible to the attachment of the tiller. Counts made are twice the average number of hyphae seen/microscope field of 0.5 mm to give the number of hyphae/mm breadth of leaf sheath.
Plant growth assessments
-
Plant growth was measured on five occasions; spring and early summer Year 1; autumn and spring Year 2; and in summer and autumn Year 3. At all samplings, live and dead tillers were counted and foliar growth above a height of 50 mm was harvested, oven-dried at 60−80 °C for 12−24 h, and weighed. Dead tillers were removed and discarded from the plants after counting and some of the dead outer sheath material which accumulated between samplings was also stripped away. Reproductive tillers were also counted and then removed in the summer samplings in year 1 and 2. After the summer harvests, aftermath heading that occurred in some plants was removed to maintain the plants in a vegetative condition.
Roots that had grown into the sand medium between the two planter bags were captured in a three-stage washing process that was also designed to remove invertebrates from the samples. The bulk of the roots were removed by hand when root material and sand were stirred in a bucket to release the invertebrates. The suspension containing the invertebrates was then decanted through sieves (see below) and the remaining sand washed through a net or wire screen with 2.5 mm2 mesh size. Roots were retrieved from both the mesh screen and sieves. Roots were later washed more thoroughly to remove any further sand and debris.
At the final assessment in late autumn of Year 3 the root and foliar growth of each plant was harvested as described above. In addition, herbage (mainly leaf sheath material), below the 50 mm cutting height, was severed from the base of the plant at ground level and kept separate from the foliar growth samples. Dead tillers were discarded, and all live tillers were immediately frozen for later freeze drying and weighing. Main plant roots were washed in the same way as root outgrowth and all samples were frozen for later freeze drying.
Herbage and root samples were oven dried at 80 °C for 36–60 h except for those retained for chemical analysis (data not presented here) which were frozen at –25 °C and later freeze-dried at ambient temperature and −0.4 mbar vacuum. All samples were weighed immediately after drying.
Insect assessments
-
At each sampling, the tillers were checked for presence of insects or signs of insect damage as they were counted. At the first two samplings in Year 1, roots were checked visually for the presence of insects and after A. lentisci were detected on roots in early summer of Year 1, their numbers were quantified in each subsequent sampling by flotation in water, wet sieving and enumeration as described by Popay and Cox[45]. Infestations of P. graminicola were recorded at the first two assessments (spring and summer Year 1) as the number of plants infested and in autumn of Year 2 as the number per tiller.
Statistical analysis
-
All data were examined for homogeneity and normality. Root aphid loadings were also log10- transformed using a constant that was based on the minimum number of aphids possible for each data set based on the dilution of the sample (usually n + 2). After analysis, log data were back transformed using the SED, number of observations and the constant used in the log transformations. Data for the percentage of tillers infested with P. graminicola were analysed using analysis of variance. All data pertaining to plant growth were not transformed. A general analysis of variance was carried out in Genstat Release 6.1 and 18, on all data using main effects of endophyte and insecticide. Block strata were based on the randomised block design for each replicate of endophyte treatments, the split-plot of the plus/minus insecticide treatments and the clonal pairs of plants for each endophyte treatment within a replicate. Means were separated using Fisher's protected least significant difference test.
Data for the percentage of vegetative tillers that were live or dead at each assessment (or reproductive for two summer assessments) were log10- transformed to ensure residuals were normally distributed with a constant variance. They were analysed using a generalised linear model with a binomial distribution and a logit link function, with the dispersion parameter fixed at 1.000. Data were back transformed for presentation here.
- Professor Warwick Silvester, Dr. Pip Gerard and Dr. Robert Hill are gratefully acknowledged for their advice during the course of this study. Valuable assistance with hyphal density counts was provided by the late Dr. Margaret di Menna. Joanne Jensen, Dr. Shirley Nichols, and Professor Stephen Goldson are thanked for reviewing a draft of the manuscript. Dr. Jonathon Newman is acknowledged for his excellent advice to reframe the paper. This research was funded by the Foundation for Research Science and Technology (now MBIE), Grasslanz Technology Ltd and PGG Wrightson Seeds.
- The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Comparisons of (a) foliar and (b) root growth (g plant–1+ SE) for endophyte treatments,764 AR1, AR37, CT and Nil over six sampling times spanning 18 months.
- Supplemental Table S1 Back-transformed aphid loadings (No. root aphid g–1) on roots of L. perenne infected with endophytes AR1, AR37 and CT, or endophyte-free (Nil) and treated (TR) or untreated (UN) with insecticide at different Sample times.
- Supplemental Table S2 Percentage of plants or tillers infested with an Phenococcus graminicola (Pseudococcidae) in ryegrass plants at three Sample times with different endophyte treatments and treated (TR) or untreated (UN) with insecticide.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Popay AJ, Cox NR. 2021. Costs and benefits of hosting Epichloë endophytes: a comparison of three haplotypes in Lolium perenne with contrasting effects on two herbivores. Grass Research 1: 8 doi: 10.48130/GR-2021-0008

Costs and benefits of hosting Epichloë endophytes: a comparison of three haplotypes in Lolium perenne with contrasting effects on two herbivores
- Received Date: 08 July 2021
- Accepted Date: 14 September 2021
- Published Online: 24 September 2021
Abstract: The symbiotic relationship between Lolium perenne and Epichloë festucae var. lolii, a fungal endophyte that lives asymptomatically in its above-ground parts, benefits both the host and fungus. In an outdoor container experiment, the effects of three distinct endophyte haplotypes (AR1, AR37 and CT) on natural infestations of a root aphid, Aploneura lentisci and a mealybug Phenococcus graminicola were compared with an endophyte-free (Nil) control. Populations of these insects were suppressed in a genetically identical set of plants treated with insecticide. Over 18 months, foliar and root growth, tiller numbers and plant mortality were compared between the different endophyte treatments and between insecticide-treated and untreated plants. Hyphal densities were recorded once during the trial. Untreated plants infected with AR37 had the lowest populations of A. lentisci and P. graminicola, and the highest root and foliar growth compared with other treatments. Growth parameters of untreated plants infected with CT often exceeded that of AR1 and Nil due to low infestations of P. graminicola and lower populations of A. lentisci. Growth of Nil, AR1 and CT plants were higher in insecticide-treated than in corresponding untreated plants but was unaffected in AR37. Untreated Nil and AR1 plants had a lower survival than AR37 and CT plants. Cumulative root outgrowth for treated AR37 plants was significantly lower than for treated AR1 on three sampling occasions. These Epichloë haplotypes are primarily mutualists, although there are circumstances when they may be disadvantageous to their host. Opposing outcomes likely originate in the ecosystems in which associations evolved.
-
Key words:
- foliar growth /
- root outgrowth /
- plant-insect interactions /
- Aploneura lentisci /
- Phenococcus graminicola /
- Lolium perenne /
- mutualism
{kind=link}