

서 언
우리나라에서 많이 재배되고 있는 일계성 딸기(june-bearing strawberry)는 여름을 지나 온도가 낮아지고 일장이 짧아지기 시작하면 꽃눈이 분화되고 더욱 저온이 진행되면 휴면상태로 월동하다가 봄이 되면 꽃을 피우고 열매를 맺는다. 이러한 생육특 성을 이용하여 다양한 재배작형이 개발되었고(Choi et al., 2016), 그에 대응하여 육묘 기술도 변화하며 발전해 왔다.
딸기 육묘는 런너 끝에서 발생된 자묘를 독립시켜 영양생장기간을 거쳐 화아분화에 필요한 생리적 요건을 갖추는 과정이다 (Savini and Neri, 2004). 이 자묘를 본포에 정식하여 휴면에 들어가지 않고 바로 수확을 시작하는 작형이 촉성재배인데, 출하시 기가 빠를수록 가격이 높기 때문에 화아분화를 빠르게 유도하기 위한 육묘기술들이 개발되었다. 화아분화가 되기 위해서는 기 본적으로 저온과 단일조건이 되어야 하지만(Manakasem and Goodwin, 1998; Fumiomi and Michael, 2006; Ruan et al., 2011; Jun et al., 2013), 식물체내 질소 농도가 낮을수록 더욱 민감하게 감응을 하며(Sonsteby et al., 2009; Uematsu, 1998), 근권용량 제한과 적엽도 촉진 효과가 있는 것으로 밝혀졌다(Takeuchi and Sasaki, 2008). 그러나 육묘기인 여름철에 저온단일처리를 하 는 것은 비용이 많이 들기 때문에 이것보다는 식물체내 질소 농도를 낮추어 반응 속도를 높이는 것이 현장의 영농기술로서 수 용되었다. 엽병의 질소 농도가 생체중 기준 200mg·L-1 이하까지 감소되었을 때 화아분화가 촉진되는 효과는 있었으나(Inoue et al., 1994; Yoshida and Morimoto, 2010), 꽃 수가 감소하였고, 화방 출현율이 낮았으며, 다음 화방의 발달이 지연되기도 하 였다(Kim et al., 2013).
한편, 시비관리를 적절히 하여 크라운 직경이 10mm 이상 되는 우량한 자묘로 육묘했을 때 동화양분 축적이 많아져서 꽃 수 가 증가하였으며, 정식 후 수량도 많았다고 보고하였다(Savini and Neri, 2004; Yoshida and Morimoto, 2010). 최근에는 촉성 재배 기술이 보편화되어 조기 수확에 대한 경제적 가치도 감소하고 있기 때문에 화아분화 촉진뿐만 아니라 가격이 높은 시기에 수확량이 많아지는 방향으로 육묘기술을 전환해야 할 시점에 있다.
딸기는 품종에 따라서 생육특성과 시비량에 따른 생육반응이 매우 다르다고 알려져 있으나(Choi et al., 2009; Choi et al., 2013), 최근 개발된 품종에 대해서는 이에 대한 연구가 충분히 이루어져 있지 않다. 본 연구는 모주로부터 증식한 자묘를 육묘 하는 과정에 질소(N), 인산(P), 칼륨(K)의 공급량을 달리하였을 때 자묘의 생육, 개화반응 그리고 정식 후 딸기 수량에 미치는 영향을 알아보기 위하여 수행하였다.
재료 및 방법
실험 재료
딸기 ‘매향’과 ‘설향’의 모주를 2013년 4월에 수경재배용 베드에 정식하여 배양액을 공급하면서 자묘를 생산하였다. 일본원 예시험장 조성액을 개량한 배양액을 EC 0.8-1.2dS·m-1의 농도로 주당 200-400mL를 하루 4-6회로 나누어 급액하였다. 초기에 발생하는 런너는 모두 제거하여 모주의 생육이 왕성해지도록 하였다. 육묘용 트레이(화성산업, 24구, cell 용량 135mL)를 5월 하순에 설치하고 상토를 충진하였다. 자묘를 순차적으로 유인하여, 6월 상순부터 육묘트레이에 핀으로 고정하여 발근을 유도 하였다. 자묘받기를 완료한 후에는 모주의 잎을 순차적으로 제거하였고, 8월 상순에 모주와 연결된 런너를 절단하고, 7일 후에 자묘를 개별적으로 분리하였다.
딸기 자묘는 9월 12일에 2중 플라스틱 온실에 110 × 18cm(2조식)로 정식하여 토경 재배하였다. 동절기 야간 온도는 최저 5°C를 기준으로 온풍난방을 실시하였으며, 기타 생육관리는 촉성재배 관리에 준하였다.
자묘에 대한 비료 공급량
자묘를 육묘하기 위한 상토는 코이어, 피트모스, 팽연왕겨를 2:3:2(v/v/v)의 비율로 혼합하여 조제하였다. 비료 공급은 완효 성 비료(Plant Food Osmocote®, N-P-K 11-11-17%, 비료용출기간 6주간)를 이용하여, 자묘 1 주당 공급하는 질소, 인산, 칼륨 의 양이 각각 25-11-32mg(이하 25-11-32), 50-22-63mg(이하 50-22-63), 100-43-127mg(이하 100-43-127)이 되도록 완효성 비 료의 양을 계산하여 육묘용 상토에 첨가한 후, 육묘트레이(점적관수용, 24구, 화성산업)에 충진하였다. 대조구(이하 0-0-0)는 완효성 비료를 첨가하지 않고 상토만 충진하였다. 자묘받기는 6월 1일부터 6월 30일까지 하였는데, 이시기에는 육묘용 상토가 마르지 않을 정도로만 3일 간격으로 점적호스를 이용하여 관수하였다. 자묘받기를 완료한, 7월 상순부터는 육묘트레이에 설치 된 점적호스에 하루 관수량을 자묘당 30-150mL로 늘려가면서 상토가 충분히 젖어 완효성 비료가 잘 용출되도록 하였다.
자묘소질, 개화반응, 수량조사
본포에 정식하기 전, 9월 상순에 자묘의 초장, 엽수, 엽면적, 건물중, 관부 직경을 측정하였다. 잎을 채취하여 흐르는 물에 수 세하여 80°C의 건조기에 72시간 동안 건조시킨 후 분쇄하여 N, P, K, Ca, 그리고 Mg 함량을 분석하였다.
자묘의 화아분화 검경은 육묘트레이에서 계속 육묘상태를 유지하면서, 처리별 5개체씩을 임의적으로 선발하여 실체현미경 (JP/SMZ800, Nikon, Tokyo, Japan) 아래서 잎을 한 장씩 제거하면서 생장점의 상태를 확인하여, 반구형 지붕 모양으로 부풀어 오르는 단계에 이르면 화아분화 시기로 간주하였다.
본포에 정식한 후 제1화방, 제2화방, 그리고 제3화방이 관찰된 날짜를 각각의 출뢰일로 간주하였다. 제1화방은 12개, 제2화 방은 7개, 제3화방은 5개의 과실만 남기고 적과를 하였고, 그 이후 화방에 대해서는 방임하였다. 딸기는 11월부터 다음해 5월까지 2-3일 간격으로 수확하여, 10g 이상인 과실을 상품과 수량으로 기록하였다.
통계처리
실험 결과에 대한 통계분석은 SAS 프로그램(SAS 9.2, SAS Institute Inc., Cary, NC, USA)을 이용하여 Duncan의 다중검정 (Duncan’ multiple range test, p < 0.05)을 실시하였다.
결과 및 고찰
본포에 정식하기 전, 자묘의 생장을 조사하여 Table 1에 나타내었다. 비료 공급량이 가장 많았던 100-43-127 처리구의 엽수 는 0-0-0 처리구와 비교하여, ‘매향’은 약 0.5매, ‘설향’은 약 1.0매가 더 많았고, 엽면적은 ‘매향’은 약 230cm2, ‘설향’은 약 250cm2가 더 넓었다. 크라운 직경은 0-0-0 처리구에서 가장 가늘었지만 다른 처리와 비교하여 통계적인 차이는 없었다. 건물중 은 100-43-127 처리구가 0-0-0 처리구보다 약 1.7g이 더 많았다. 두 품종 모두, 육묘기간에 비료 공급량이 많을수록 자묘의 생 장량이 증가하는 경향을 보였다.
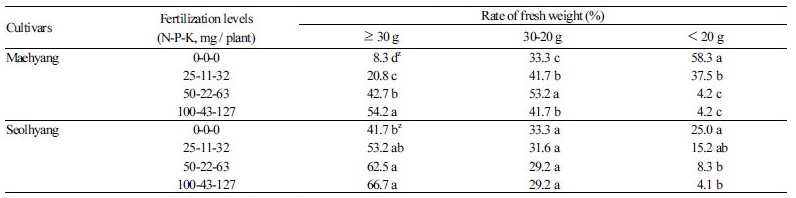
자묘의 생체중을 측정하여 그 비율을 Table 2에 나타내었다. 0-0-0 처리구에서, ‘매향’은 생체중 20g 이하 소묘의 비율이 58.3%로 매우 높았고, 생체중 30g 이상 대묘의 비율은 8.3%로 낮았다. 비료 공급량이 많아질수록 대묘의 비율이 증가하였는 데, 25-11-32 처리구에서 20.8%, 50-22-63 처리구에서 42.7%, 100-43-127 처리구에서는 54.2%로 높아졌다. 50-22-63 처리 구와 100-43-127 처리구에서는 소묘의 비율이 4.2%로 현저하게 낮았다. ‘설향’은 0-0-0 처리구에서도 대묘의 비율이 41.7%에 이르렀고 소묘의 비율은 25.0%에 불과해, ‘매향’에 비해 대체로 생체중이 많은 경향을 보였다.
딸기는 육묘기간에 모주로부터 15-30개의 자묘가 발생하는데, 자묘는 런너를 통하여 모주로부터 양수분을 공급받지만 뿌리 가 발달하기 시작하면 스스로 양수분을 흡수하며 생장한다. 0-0-0 처리와 같이 자묘에 비료 공급을 하지 않은 경우, 모주로 부터 공급받는 양분분배량에는 상대적인 차이가 생길 것이며, 이는 생육에도 영향을 주어 일찍 발생한 자묘일수록 크기가 증가하였 다(Table 2). 또 런너를 절단함과 동시에 모주로부터 양분공급이 중단되므로 양분부족 상태가 되어, 늦게 발생한 자묘일수록 생육이 불량한 소묘가 되었다고(Table 1) 생각되었다. 그러므로 균일한 상태의 묘를 대량생산한다는 육묘의 관점에서 보면, 빨 리 모주와 연결된 런너를 절단한 후에 동일한 시비관리를 하는 것이 자묘의 지하부 발달을 촉진할 뿐 아니라(Kim et al., 2013), 자묘간 생장량의 격차를 줄일 수 있을 것이라 생각되었다.
육묘기간 자묘당 질소(N)의 공급량을 0에서 100mg까지 달리하였을 때, 잎의 T-N 함량은 ‘매향’은 1.20%에서 1.68%까지, ‘설향’은 1.37%에서 1.65%까지 증가하였다(Table 3). 인(P)의 공급량을 0에서 43mg까지 달리하였을 때, 잎의 P 함량은 ‘매향’ 은 0.14%에서 0.25%까지, ‘설향’은 0.17%에서 0.23%까지 증가하였다. 잎의 칼륨(K) 함량도, 공급량이 0에서 127mg으로 달 리하였을 때, ‘매향’은 0.94%에서 1.13%, ‘설향’은 1.12%에서 1.28%까지 증가하였다. 질소, 인산, 칼륨의 공급량이 많아질수 록 식물체내 농도도 대체로 증가하는 경향을 보였는데, 3요소 성분 중 P 함량이 상대적으로 많이 증가하였고, K 함량은 상대적 으로 적게 증가하였다.
Table 1. The effect of fertilization levels on growth of daughter plants in strawberry
 | |
zMean separation within columns in each cultivar by Duncan’s multiple range test, p ≤ 0.05. | |
Table 2. The effect of fertilization levels on fresh weight distribution of daughter plants in strawberry
 | |
zMean separation within columns in each cultivar by Duncan’s multiple range test, p ≤ 0.05. | |
‘매향’의 생장점이 꽃눈으로 분화되는 시기는 0-0-0, 25-11-32, 50-22-63 처리구에서 9월 13일로 빨랐고, 100-43-127 처리 구는 이보다 9일이 지연되었다(Table 4). 본포 정식 후, 제 1화방이 출뢰한 날은 0-0-0, 25-11-32, 50-22-63 처리구에서는 10월 27일이었고, 100-43-127 처리구에서는 11일이 늦어졌다. 딸기를 처음 수확한 날은 0-0-0, 25-11-32 처리구는 12월 7일이었고, 50-22-63과 100-43-127 처리구에서는 각각 3과 11일이 늦어졌다.
‘설향’의 화아분화 시기는 0-0-0 처리구가 9월 13일로 가장 빨랐고, 25-11-32, 50-22-63, 100-43-127 처리구는 차례로 4일, 9일, 19일이 늦어졌다. 제1화방 출뢰일은 50-22-63, 100-43-127 처리구에서 0-0-0 처리구와 비교하여 각각 5일, 15일이, 그리 고 수확개시일은 각각 10일, 27일이 늦어졌다.
딸기 크라운 정단부에 있는 생장점은 잎눈을 분화하다가 저온단일 조건에 노출되면 잎눈의 분화를 멈추고 꽃눈을 분화하게 된다. ‘매향’과 ‘설향’ 모두, 육묘기에 비료 공급량이 많아질수록 식물체내 질소 농도가 높아져, 저온과 단일 조건에 빠르게 반 응을 하지 못하여 화아분화가 늦어지는 경향을 보였다(Inoue et al., 1994; Yosida and Morimoto, 2010; Kim et al., 2013). 본 연구에서 자묘를 본포에 정식한 것은 9월 12일인데, 이때 ‘매향’의 100-43-127, ‘설향’의 50-22-63, 100-43-127 처리구에서는 생장점이 화아분화가 되지 않은 상태였다. 이들은 본포 정식과 함께 질소 비료가 급속히 흡수되는 조건이 되었으므로 실제의 화아분화는 Table 4보다 훨씬 지연되었을 것이라 생각되었다.


제1화방 후 제2화방 출뢰까지 소요일수를 산출했을 때, ‘설향’은 비료 공급량에 따라 큰 차이가 없이 37-40일 범위였다 (Table 4). 즉, 제1화방이 늦게 출뢰한 경우, 제2화방도 늦게 출뢰되는 경향을 보였다. 그러나 ‘매향’은 100-43-127 처리구에서 33일로서, 다른 처리의 41-43일보다 9일이나 단축되었다. 즉, ‘매향’은 육묘기간 질소 공급량이 많았을 때, 제1화방의 화아분 화는 다소 지연되었지만, 화방출현율이 높았고 제2화방의 출뢰가 빨라져 수확이 집중되는 등 긍정적인 효과를 보였다.
한편, 자묘에 비료 공급을 하지 않았던 0-0-0 처리에서도 잎의 T-N 함량이 약 1.2% 수준으로 극단적으로 낮아지지 않았던 (Table 3) 이유는, 런너를 절단한 8월 상순 이전까지는 런너를 통해서 모주로부터 양분이 충분히 전달되었기 때문이라 생각되 었다. 그러나 이런 경우에도 화아분화에 효과적이라고 알려진 질소 농도 200mg·L-1(Inoue et al., 1994)보다는 높은 상태였으 며, 본 연구가 잎의 건물중 기준이고 그들은 엽병의 생체중 기준이었다는 것을 감안하더라도 상당한 차이를 보였다. 이런 이유 로 보아 화아분화 촉진을 위해서는 모주와 자묘를 일찍 분리하여, 균일하게 비료를 공급하고 일시에 중단하는 방법으로 관리를 하는 것이 유리할 것이라 생각되며, 이에 대해서는 추후 정밀한 검토가 필요한 것으로 생각되었다.
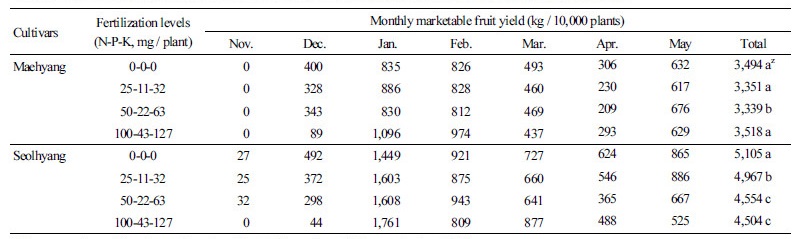
두 품종 모두 육묘기간 비료 공급량이 적을수록 수확개시가 빨라져, 판매 가격이 높은 11월부터 1월까지의 수량이 많았다 (Table 5). ‘매향’의 월별 수량은 0-0-0, 25-11-32, 50-22-63 처리구에서 비슷한 경향을 보였다. 100-43-127 처리구는 수확개시 가 늦었기 때문에 12월 수량은 적었으나 1월과 2월의 수량은 현저히 많았고, 3월 이후에는 다른 처리와 큰 차이가 없었다. 다음 해 5월까지 총수량은 모든 처리구에서 3,339-3,519kg 범위로 큰 차이가 없었다.
그러나 ‘설향’의 경우에는, 100-43-127 처리구의 수량이 0-0-0 처리구와 비교하여, 12월에는 9%, 1월에는 122%, 2월에는 88%, 3월에는 121%, 4월에는 78%, 5월에는 61% 수준으로 후기로 갈수록 크게 감소하였다. 다음해 5월까지 총수량은 0-0-0 처리구에서 5,106kg으로 가장 많았고, 50-22-63와 100-43-127 처리구에서는 약 12%가 감소하였다.
비료 공급량이 많았던 100-43-127 처리구에서, ‘설향’은 제1화방 출뢰와 수확개시가 상당히 지연되었기 때문에 그동안 제1 화방에 많은 동화양분이 축적되었고, 그 결과 1월 중의 수량이 1,605kg으로서 큰 peak가 발생하였으며 2월에는 감소, 반동으 로 3월에는 다시 peak가 발생하였다. 즉, 제1화방의 과도한 착과로 인하여 동화산물이 과실로 집중되고 다른 영양생장기관으 로의 분배가 감소하여 초세가 약해지고 결국 다음 화방의 발달도 억제되는(Nishizawa, 1994; Shimomura et al., 2016) 결과를 초래하였다고 생각되었다.
본 연구의 결과를 종합해 볼 때, 육묘기간 비료 공급량이 많아질수록 자묘의 생장은 증가하는 경향이었으나, 식물체내 질소 농 도가 높아졌고 화아분화와 수확개시는 현저하게 지연되었다. 특히, ‘설향’의 화아분화는 비료 공급량에 민감하게 반응하여, 자 묘당 질소 100mg 이상 공급한 경우에는 화아분화는 19일, 수확개시는 26일이나 지연되었다. 그 결과, 제1화방에 과다하게 축 적된 양분으로 인해 1월의 수량이 크게 증가하였으나 착과부담으로 생육저하와 수량감소로 이어져 안정생산에는 문제가 있었 다. ‘매향’역시 비료 공급량이 많은 처리에서 화아분화가 지연되기는 하였으나, 11일 정도로 비교적 짧았고, 제2화방이 빠르게 출뢰하여 단가가 높은 3월 이전까지의 수확량이 많았으므로 촉성재배 농가에서 적용하면 경제성이 높을 것으로 생각되었다. 이와 같이 육묘기 비료 공급량이 개화반응 및 수량에 미치는 영향은 품종에 따라 차이를 보이며, 모든 품종에서 대묘로 육묘 를 하는 것이 수확량 증가로 이어지는 것은 아니므로 품종에 따라 시비량을 조절할 필요가 있는 것으로 판단되었다.
