
Comparative genomics of 16 Microbacterium spp. that tolerate multiple heavy metals and antibiotics
- Published
- Accepted
- Received
- Academic Editor
- Blanca Landa
- Subject Areas
- Genomics, Microbiology, Biogeochemistry, Environmental Contamination and Remediation
- Keywords
- Microbacterium, Chromium reduction, Genomics, Antibiotic resistance, Heavy metals
- Copyright
- © 2019 Learman et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2019. Comparative genomics of 16 Microbacterium spp. that tolerate multiple heavy metals and antibiotics. PeerJ 6:e6258 https://doi.org/10.7717/peerj.6258
Abstract
A total of 16 different strains of Microbacterium spp. were isolated from contaminated soil and enriched on the carcinogen, hexavalent chromium [Cr(VI)]. The majority of the isolates (11 of the 16) were able to tolerate concentrations (0.1 mM) of cobalt, cadmium, and nickel, in addition to Cr(VI) (0.5–20 mM). Interestingly, these bacteria were also able to tolerate three different antibiotics (ranges: ampicillin 0–16 μg ml−1, chloramphenicol 0–24 μg ml−1, and vancomycin 0–24 μg ml−1). To gain genetic insight into these tolerance pathways, the genomes of these isolates were assembled and annotated. The genomes of these isolates not only have some shared genes (core genome) but also have a large amount of variability. The genomes also contained an annotated Cr(VI) reductase (chrR) that could be related to Cr(VI) reduction. Further, various heavy metal tolerance (e.g., Co/Zn/Cd efflux system) and antibiotic resistance genes were identified, which provide insight into the isolates’ ability to tolerate metals and antibiotics. Overall, these isolates showed a wide range of tolerances to heavy metals and antibiotics and genetic diversity, which was likely required of this population to thrive in a contaminated environment.
Introduction
Heavy metals, while naturally occurring, cause harm to both human and ecosystem health. Specifically, chromium is a mutagen and carcinogen and its presence in the environment can be natural due to anthropogenic activities, such as industrial manufacturing (e.g., metal plating and tanneries) and mining (chromite ore) (Barak et al., 2006; Brose & James, 2010; Cheng, Holman & Lin, 2012). In the environment, chromium can exist as hexavalent chromium [Cr(VI)], which is soluble and more toxic, or as insoluble trivalent chromium [Cr(III)] (Bartlett, 1991; Cheng, Holman & Lin, 2012). Thus, the redox cycling of chromium is critical to understanding its impacts on the environment. In natural systems, Cr(III) can be oxidized by manganese oxides and hydrogen peroxide, while Cr(VI) can be reduced by ferrous iron and hydrogen sulfides (Brose & James, 2010; Oze et al., 2004; Viti et al., 2013).
Many bacteria are able to tolerate Cr(VI) stress based on their ability to transport it outside the cell or enzymatically reduce it to the less toxic form Cr(III). The efflux pump, chrA, has been associated with providing Cr(VI) resistance by transporting it outside the cell (Cervantes & Ohtake, 1988; Cervantes et al., 1990; Nies, Nies & Silver, 1990). A 2008 study found 135 chrA orthologs of this efflux pump (Ramirez-Diaz et al., 2008); however, the presence of chrA is not always a sole predictor of the amount of Cr(VI) that bacteria can resist (Henne et al., 2009a). Bacteria can also have additional efflux pumps that provide resistance to other metals (Nies, 2003; Silver, 1996), which is advantageous as anthropogenically impacted sites are often contaminated with multiple stressors. Further, there is a well-known association between metal and antibiotic tolerance (Baker-Austin et al., 2006; Seiler & Berendonk, 2012; Wright et al., 2006). For example, Staphylococcus species use multiple efflux pumps to tolerate chromium, lead, and penicillin (Ghosh et al., 2000) and Salmonella abortus equi strains were able to tolerate chromium, cadmium, mercury, and ampicillin (Ug & Ceylan, 2003). Thus, efflux pumps can play a vital role in how bacteria thrive in contaminated ecosystems.
Bacteria are also known to catalyze the reduction of Cr(VI) aerobically. Specifically, certain bacteria use a soluble NADH/NADPH-dependent oxidoreductase to reduce Cr(VI) (Ackerley et al., 2004a; Barak et al., 2006; Cheung & Gu, 2007; Gonzalez et al., 2005; Park et al., 2000). Two well-studied Cr(VI) reductases are chrR and yieF. In Pseudomonas putida, chrR reduces Cr(VI) via the transfer of one electron, generating Cr(V), a reactive intermediate, which then requires a second electron transfer to generate the more stable insoluble Cr(III) (Park et al., 2000). Conversely, Escherichia coli uses yieF to catalyze the transfer of four electrons to reduce Cr(VI) (Ackerley et al., 2004b; Barak et al., 2006; Ramirez-Diaz et al., 2008).
The objective of this study was to examine the genomes of 16 Microbacterium spp. strains isolated from metal contaminated soil to identify their genetic potential and physiological ability to reduce Cr(VI) and to tolerate both heavy metals and antibiotics. A recent genomic study of four Cr(VI) reducing Microbacterium spp. identified two putative reductases (Henson et al., 2015), one genetically similar to chrR in Thermus scotoductus (Opperman, Piater & van Heerden, 2008) and the other to yieF of Arthrobacter sp. RUE61a (Niewerth et al., 2012); however, these putative reductases had low sequence similarity to the well-studied reductases found in Pseudomonas putida and E. coli. While other studies have shown Microbacterium isolates are able to reduce Cr(VI) (Humphries et al., 2005; Liu et al., 2012; Pattanapipitpaisal, Brown & Macaskie, 2001; Soni et al., 2014), there are still unknowns related to the genetics of Cr(VI) reduction and resistance. Further, the study by Henson et al. (2015) documented high levels of genomic variability between the isolates, which might be related to ecotypes. However, the conclusions of this study were limited based on the low number of genomes examined. Comparative genomics has been used in various environments and microorganisms to examine inter-strain variation and ecotypes (Briand et al., 2009; Coleman & Chisholm, 2010; Denef et al., 2010; Frangeul et al., 2008; Humbert et al., 2013; Martiny, Coleman & Chisholm, 2006; Meyer & Huber, 2014; Meyer et al., 2017). Thus, this study examined the genomic and physiological variability of heavy metal and antibiotic resistance of 16 Microbacterium strains from the same contaminated soil environment to elucidate the potential genomic flexibility of closely related strains.
Methods
Bacterial isolation
Isolation of Microbacterium spp. from soil is described in Kourtev, Nakatsu & Konopka (2009) and Henson et al. (Henson et al., 2015). Bill Jervis from the Indiana Department of Transport provided site access (no permit was required) and the project did not involve endangered or protected species. Briefly, contaminated soil samples were collected from the Department of Transportation site in Seymour, IN, USA. Previous studies have documented the site was contaminated with Pb (1,156 μg−1 soil), Cr (5,868 μg−1 soil), and hydrocarbons (toluene and xylenes: >200 μg g−1 soil) (Joynt et al., 2006; Kourtev, Nakatsu & Konopka, 2006; Nakatsu et al., 2005). Bacteria were initially isolated from the soil on 50% tryptic soy agar amended with 0.25 mM Cr(VI) (K2CrO4). From this, 16 isolates were further selected based on their ability to grow on at least 0.5 mM Cr(VI). The isolates were stored in glycerol stocks at −80 °C until further use.
Cr(VI) reduction
For Cr(VI) reduction experiments, isolates were grown in 25% tryptic soy broth (TSB) with 0.5 or 1 mM Cr(VI) at 30°C and 225 rpm for 72 h. Two concentrations were used as certain isolates had relatively lower growth at 1 mM compared to other isolates. The cultures were pelleted and Cr(VI) reduction was determined using a colorimetric assay (Henson et al., 2015; Urone, 1955).
DNA sequencing and bioinformatics
Isolates were grown in TSB with 0.5 or 2 mM Cr(VI) at 30 °C and 225 rpm for 24–72 h. The cultures were pelleted via centrifugation and stored at −20 °C until used in DNA extractions. FastDNA Spin Kits (MP Biomedical) were used to extract genomic DNA from the thawed pellets. The resulting DNA was stored at −20 °C. The DNA from the 16 bacterial isolates were sent to Cincinnati Children’s Hospital Medical Center’s Genetic Variation and Gene Discovery Core facility for whole genome shotgun sequencing using one lane of an Illumina HiSeq 2000 (100bp PE). The resulting raw genomic reads can be found in the National Center for Biotechnology Information’s (NCBI) Short Read Archives (SRA), accession number: SRP120551.
Raw reads were evaluated for quality with FastQC (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/) and trimmed and quality filtered with Trimmomatic (v 0.33) (Bolger, Lohse & Usadel, 2014). Sequence quality scores were low on R2 (assessed by FastQC), so the trimmed reads were cut (length of 70bp) and passed through another quality filter (-Q33 -q 30 -p 50) with FastX toolkit (http://hannonlab.cshl.edu/fastx_toolkit/). Reads for each sample were subsampled to four million reads (multiple subsampling depths were tested) with seqtk (https://github.com/lh3/seqtk). Using multiple different read depths, MEGAhit (Li et al., 2016) was used to assemble the data, which were then assessed for quality with QUAST (Gurevich et al., 2013). CheckM (Parks et al., 2015) was used to examine genome completeness and contamination (taxon marker Microbacterium). The resultant genomes were annotated by the Department of Energy’s Joint Genome Institute Integrated Microbial Genomes (IMG) system (Markowitz et al., 2012) and are publicly available (see Table S1). A pangenomic analysis of 20 genomes (from this study and Henson et al., 2015) was conducted using the An’vio program (version 3) (Eren et al., 2015) following the pangenomics workflow from Delmont & Eren (2018). The pangenomic analysis within An’vio also utilized other programs like HMMER (Eddy, 2011), Prodigal (Hyatt et al., 2010), and NCBI’s blastp (Altschul et al., 1997). An’vio was also used to define clusters of orthologous groups (COGs) for the pangenomic analysis using the command anvi-run-ncbi-cogs.
All 16, 16S rRNA gene sequences from the Cr(VI) isolates were downloaded from IMG. These genes were used to generate a maximum likelihood phylogenetic tree in MEGA7 (Kumar, Stecher & Tamura, 2016) and evolutionary history was inferred using the maximum likelihood Tamura–Nei model (Tamura & Nei, 1993). Within IMG, annotations were searched for genes related to Cr(VI) resistance and Cr(VI) reduction via keywords (e.g., efflux, reductase, and chromate). Annotated Cr(VI) reductase genes were compared via BLASTP (one-way comparison on NCBI’s website) to a known Cr(VI) reducing chrR from Pseudomonas putida KT2440 (NP_746257) (Ackerley et al., 2004b). Genes related to heavy metal resistance and antibiotic resistance were searched using functional categories. Beta-lactam, chloramphenicol, and vancomycin resistance genes were also identified using KEGG pathway KO identifiers. The COG category “Inorganic ion transport and metabolism” was used to find annotated heavy metal efflux genes and metal resistance genes.
Metal and antibiotic tolerance assays
Bacterial isolates were grown on agar plates (10% TSB, 1.5% agar, and 0.5 mM Cr(VI)) at 30 °C until colonies were visible. Colonies were then streaked onto fresh agar plates amended with different concentrations of heavy metals (Cd 0–5 mM, Co 0–5 mM, Cr 0–20 mM, Cu 0–1 mM, Ni 0–15 mM, and Zn 0–1 mM). Duplicate plates were incubated at 30 °C for 14 days and growth was confirmed by the presence of visible colonies. All bacterial isolates were plated on each metal concentration in duplicate.
The Minimum Inhibitory Concentration (MIC) for ampicillin, chloramphenicol, and vancomycin was tested on each isolate using MIC test strips (Liofilchem®). To do this, each of the 16 Microbacterium spp. isolates was streaked onto agar plates (50% TSB and 1.5% agar) with 0.5 mM Cr and incubated at 30 °C for 5 days. Next, each isolate was suspended in a 0.85% sterile saline solution until it reached a density that approximated the 0.5 McFarland Turbidity Standard. The isolates were then spread uniformly on a Mueller–Hinton agar plate. A MIC test strip was placed in the center of each plate. The plates were incubated at 30 °C for 24 h. After the 24 h, the plates were removed and the MIC of each isolate to each antibiotic was determined by visually documenting the zones of inhibition.
Results and Discussion
Isolation of Cr(VI) reducing bacteria
Though originally isolated and studied for their ability to reduce Cr(VI) (Henson et al., 2015; Kourtev, Nakatsu & Konopka, 2009), it was speculated that these isolates may also be tolerant to other contaminants, because the soils were contaminated with other heavy metals and organic solvents (Joynt et al., 2006; Kourtev, Nakatsu & Konopka, 2006; Nakatsu et al., 2005). Among the 16 isolates studied here, a wide range of Cr(VI) tolerance and reducing ability was found (Table 1). The majority (13) of the isolates could tolerate two mM Cr(VI), while three of the isolates (K2B2, K36, and K40) could only tolerate 0.5 mM Cr(VI). Interestingly, five of the 13 isolates that were able to tolerate two mM Cr(VI) (A20, K24, K30, K33, and PF3) had observable colonies on plates containing up to 20 mM Cr(VI). Further, the 16 isolates also showed a wide range in Cr(VI) reduction (0–88.8%) over a 72-h incubation (Table 1). Only two isolates, K27 and K33, reduced over 80% of the Cr(VI) in the experiment. Interestingly, K27 reduced 83% of the Cr(VI) in liquid medium but was only able to tolerate 0.5 mM Cr(VI) when grown on solid medium, while K33 tolerated up to 20 mM Cr(IV) and reduced 88% of the Cr(IV) present. Two additional isolates, K31 and PF3, reduced over 50% of the Cr(VI) in the experiment. There was no statistical relationship between the ability of isolates to tolerate and their ability to reduce Cr(VI).
| Metal tolerance (mM) | |||||||
|---|---|---|---|---|---|---|---|
| Isolate | Cr | Ni | Co | Cd | Zn | Cu | Cr reduction (%)* |
| Microbacterium sp. A20 | 20.0 | 1.0 | 0.1 | 0.1 | 1.0 | 1.0 | 41.6 |
| Microbacterium sp. K19 | 2.0 | 1.0 | 0.1 | 0.1 | 1.0 | 1.0 | 34.0 |
| Microbacterium sp. K21 | 2.0 | 1.0 | 0.1 | 0.1 | 1.0 | 1.0 | 39.3 |
| Microbacterium sp. K22 | 2.0 | 1.0 | 0.1 | 0.1 | 0.1 | 1.0 | 42.9 |
| Microbacterium sp. K24 | 20.0 | 1.0 | 1.0 | 0.0 | 1.0 | 1.0 | 41.6 |
| Microbacterium sp. K27 | 2.0 | 1.0 | 1.0 | 0.1 | 0.1 | 1.0 | 83.0 |
| Microbacterium sp. K2B2 | 0.5 | 0.1 | 0.0 | 0.1 | 0.1 | 0.1 | 36.0 |
| Microbacterium sp. K30 | 20.0 | 1.0 | 1.0 | 0.1 | 1.0 | 1.0 | 41.6 |
| Microbacterium sp. K31 | 2.0 | 0.1 | 0.1 | 0.1 | 1.0 | 1.0 | 55.3 |
| Microbacterium sp. K33 | 20.0 | 1.0 | 0.1 | 0.1 | 1.0 | 1.0 | 88.8 |
| Microbacterium sp. K35 | 2.0 | 0.1 | 0.1 | 0.1 | 0.1 | 1.0 | 3.6 |
| Microbacterium sp. K36 | 0.5 | 1.0 | 0.1 | 0.1 | 0.1 | 1.0 | 10.7 |
| Microbacterium sp. K40 | 0.5 | 1.0 | 0.1 | 0.1 | 1.0 | 1.0 | 30.3 |
| Microbacterium sp. K41 | 2.0 | 1.0 | 0.1 | 0.0 | 0.1 | 1.0 | 28.1 |
| Microbacterium sp. K5D | 2.0 | 1.0 | 0.1 | 0.0 | 0.1 | 1.0 | 0.0 |
| Microbacterium sp. PF3 | 20.0 | 1.0 | 0.1 | 0.0 | 1.0 | 1.0 | 50.6 |
Notes:
Heavy metal (cadmium, chromium, cobalt, copper, nickel, and zinc) tolerance (in mM) of each isolate and chromate reduction (%) data.
Data shown in percentages since two chromate concentrations were used.
Genomic assembly and pangenomic analysis
Previous research has shown Cr(VI) tolerance to be related to a resistance gene (chrA) (Cervantes & Ohtake, 1988; Cervantes et al., 1990) and/or Cr(VI) reductases (chrR and yieF; (Ackerley et al., 2004b; Barak et al., 2006; Park et al., 2000; Ramirez-Diaz et al., 2008). To gain a better understanding of the Cr(VI) tolerance of these isolates, their genomes were assembled and annotated. The isolate genomes ranged in total length from 3.31 to 4.23 Mb and all had GC content ranging from 68 to 70%% (Table 2). A genomic study of 10 Macrobacterium sp. documented similar GC ranges and genome length (Corretto et al., 2015). The assembled genomes were generally over 96.7% complete and all genomes had under 4% contamination (Table 2).
| Isolate | Total length (Mb) | GC (%) | Number of contigs | Largest contig (bp) | N50 (bp) | Est. Sequencing coverage | Comp. (%) | Cont. (%) | Strain heterogeneity (%) |
|---|---|---|---|---|---|---|---|---|---|
| Microbacterium sp. A20 | 3.94 | 68.53 | 75 | 436,954 | 166,569 | 142 | 99.06 | 0.54 | 0 |
| Microbacterium sp. K19 | 3.89 | 68.69 | 54 | 479,964 | 187,157 | 144 | 99.44 | 1.01 | 20 |
| Microbacterium sp. K21 | 3.85 | 68.33 | 34 | 537,739 | 330,631 | 146 | 99.06 | 0.18 | 0 |
| Microbacterium sp. K22 | 3.94 | 68.52 | 96 | 258,133 | 83,317 | 142 | 99.06 | 0.71 | 33.33 |
| Microbacterium sp. K24 | 4.23 | 68.90 | 142 | 248,507 | 56,118 | 132 | 99.35 | 3.93 | 9.09 |
| Microbacterium sp. K27 | 3.75 | 68.51 | 37 | 342,064 | 187,856 | 149 | 98.66 | 0.18 | 0 |
| Microbacterium sp. K2B2 | 3.94 | 68.52 | 76 | 436,909 | 120,725 | 142 | 99.06 | 0.54 | 0 |
| Microbacterium sp. K30 | 4.06 | 69.16 | 96 | 295,140 | 88,745 | 138 | 98.39 | 2.61 | 0 |
| Microbacterium sp. K31 | 3.77 | 68.43 | 39 | 443,452 | 183,886 | 148 | 98.66 | 0.18 | 0 |
| Microbacterium sp. K33 | 3.89 | 68.68 | 110 | 311,089 | 66,684 | 144 | 99.44 | 1.49 | 0 |
| Microbacterium sp. K35 | 3.62 | 70.91 | 379 | 168,068 | 17,080 | 155 | 96.70 | 1.82 | 12.5 |
| Microbacterium sp. K36 | 3.31 | 70.53 | 91 | 183,861 | 73,738 | 169 | 98.10 | 0.60 | 0 |
| Microbacterium sp. K40 | 3.77 | 68.40 | 45 | 442,332 | 195,355 | 148 | 99.06 | 0.09 | 0 |
| Microbacterium sp. K41 | 3.62 | 70.75 | 454 | 89,969 | 15,114 | 155 | 97.62 | 1.37 | 0 |
| Microbacterium sp. K5D | 3.80 | 68.35 | 49 | 442,284 | 195,368 | 147 | 99.06 | 0.09 | 0 |
| Microbacterium sp. PF5 | 3.74 | 70.80 | 144 | 125,065 | 47,267 | 150 | 97.23 | 1.82 | 0 |
Note:
Genome assembly statistics (total length, GC%, and number of contigs) and quality assurance data (completeness, contamination, and strain heterogeneity).
Phylogenetic analysis of the 16S rRNA gene indicated that all of the isolates belong to the genus Microbacterium (Fig. S1; Table S2). Specifically, the isolates were closely related to Microbacterium oxydans, Microbacterium maritypicum, or Microbacterium paraoxydans. Microbacterium spp. have been isolated from sites contaminated with metals (Avramov et al., 2016; Bollmann et al., 2010; Corretto et al., 2015), radionuclides (Nedelkova et al., 2007), and petroleum (Avramov et al., 2016; Chauhan et al., 2013; Wang et al., 2014). Moreover, some environmental isolates, such as Microbacterium laevaniformans strain OR221, have shown to tolerate multiple metals (Ni, Co, and Cd) (Bollmann et al., 2010). A genomic analysis of the latter strain provided evidence of genes (e.g., transporter and detoxification genes) that could aid the strain’s ability to tolerate metals (Brown et al., 2012). Other Microbacterium spp. have been known to reduce Cr(VI): Microbacterium sp. SUCR140 (Soni et al., 2014), Microbacterium sp. chr-3 (Focardi, Pepi & Focardi, 2013), and Microbacterium sp. CR-07 (Liu et al., 2012).
The pan-genome size of the 20 isolates was 7,902 protein clusters with the core genome encompassing 2,073 protein clusters (26%) (Fig. 1). The majority of the genes in the core genome of the 20 isolates (36%) were placed in the “Metabolism” COG (Fig. 2). Genes in the COG categories “Cellular processes and signaling” and “Information storage and processing” comprised 21% and 17%, respectively of the core genome (Fig. 2). The flexible pangenome was dominated by genes in the COG categories “Metabolism” and “Poorly characterized/No assigned COG” (36% for both) (Fig. 2). Relative to the core pangenome, the flexible pangenome had few genes in the COG categories “Cellular processes and signaling (21%)” and “Information storage and processing 17%” (Fig. 2). The percentage of core genes in a genome can vary from 3% to 84% (McInerney, McNally & O’Connell, 2017). For example, the Bacillus cereus core genome is 27% of the pangenome, whereas the core genome of Bacillus anthracis is 65% (McInerney, McNally & O’Connell, 2017). An analysis of 20 Microcystis sp. showed the genome was comprised of 34–49% core genes and 51–66% flexible genes (Meyer et al., 2017). A study on the genomes of 12 Prochlorococcus isolates found a core genome ranging from 40% to 67% (Kettler et al., 2007). The relatively large genomic variability seen with the Microbacterium spp. examined here could be due to selective pressures that drove gene loss or horizontal gene transfer, which in the end enhanced the ability of the isolates to survive in a contaminated sediment environment.
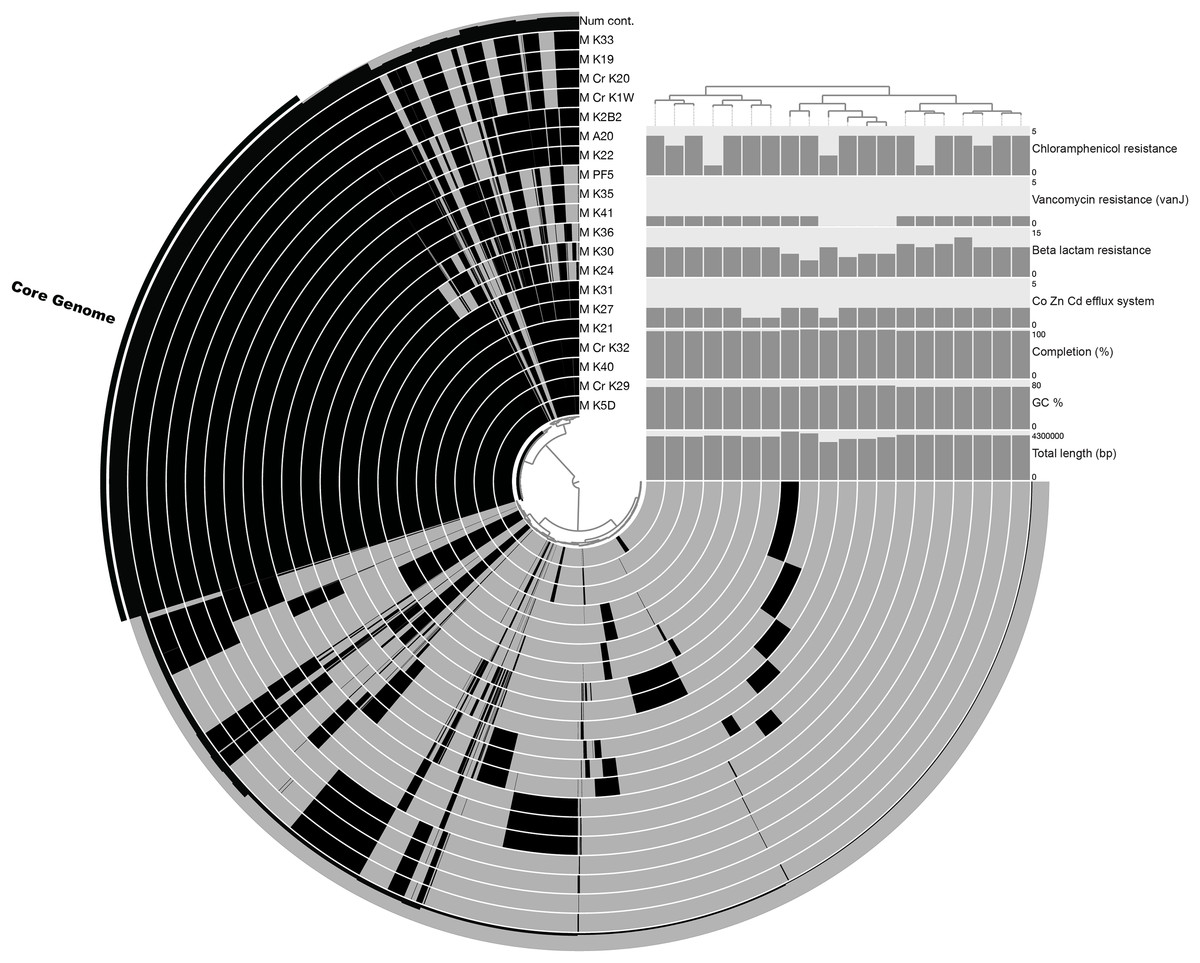
Figure 1: Pangenomic analysis of heavy metal tolerant Microbacterium spp.
The figure was made in An’vio with the items order in presence/absence (D: Euclidean; L: Ward) and the samples ordered by PC frequency. The outer most complete ring (labeled Num cont.) represents the number of genomes that a gene occurs in, with the core genome highlighted with an additional black bar. The internal rings represent an isolate’s genome with the black color representing the presence of genes in a genome (gray is the absence of a gene). The bar charts show quality control measurements (total genome length [base pairs], GC content, and percent completion) and also the abundance of genes that were annotated as Co, Zn, Cd efflux system genes and antibiotic tolerance genes (beta lactam, vancomycin, and chloramphenicol resistance).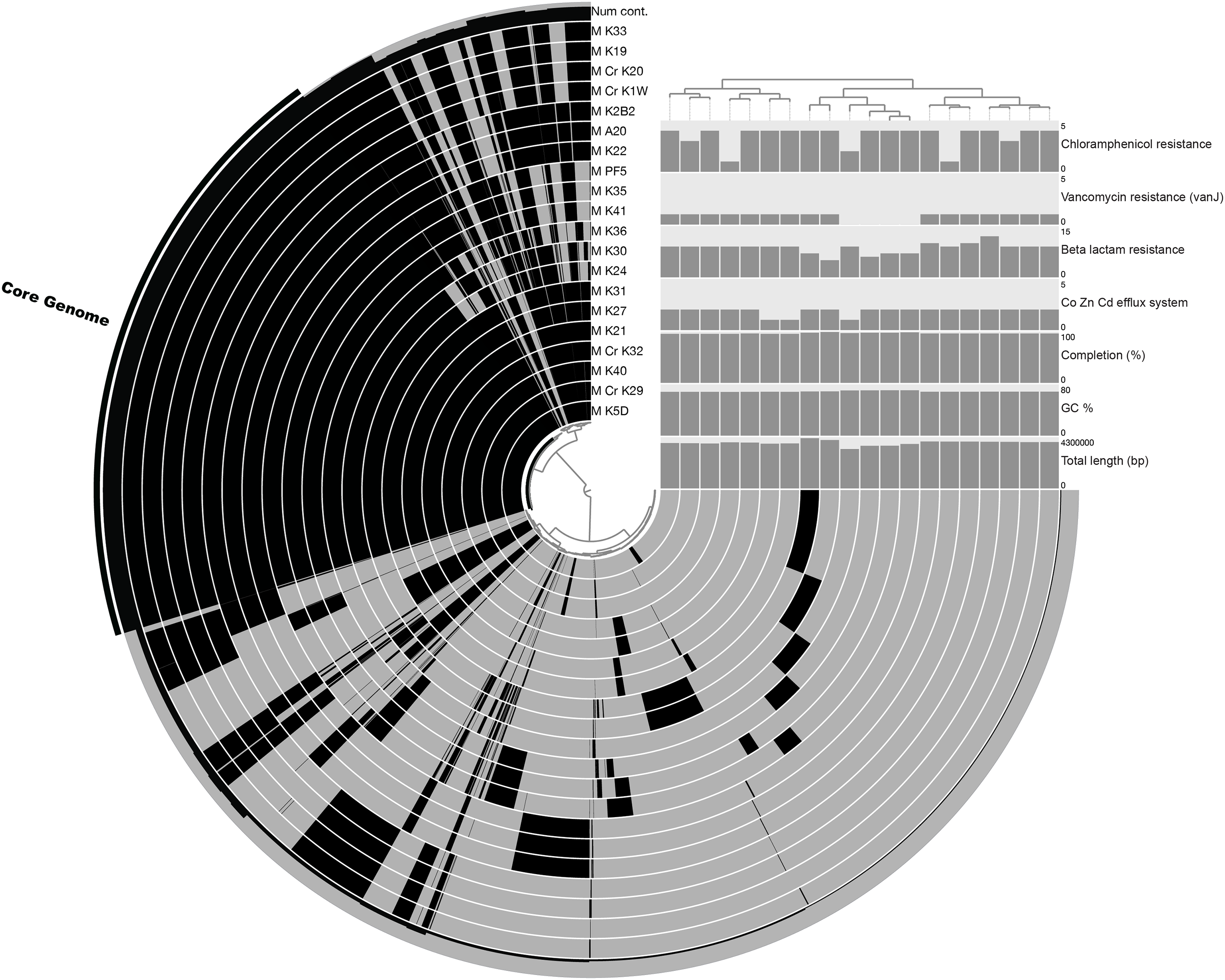{kind=link}
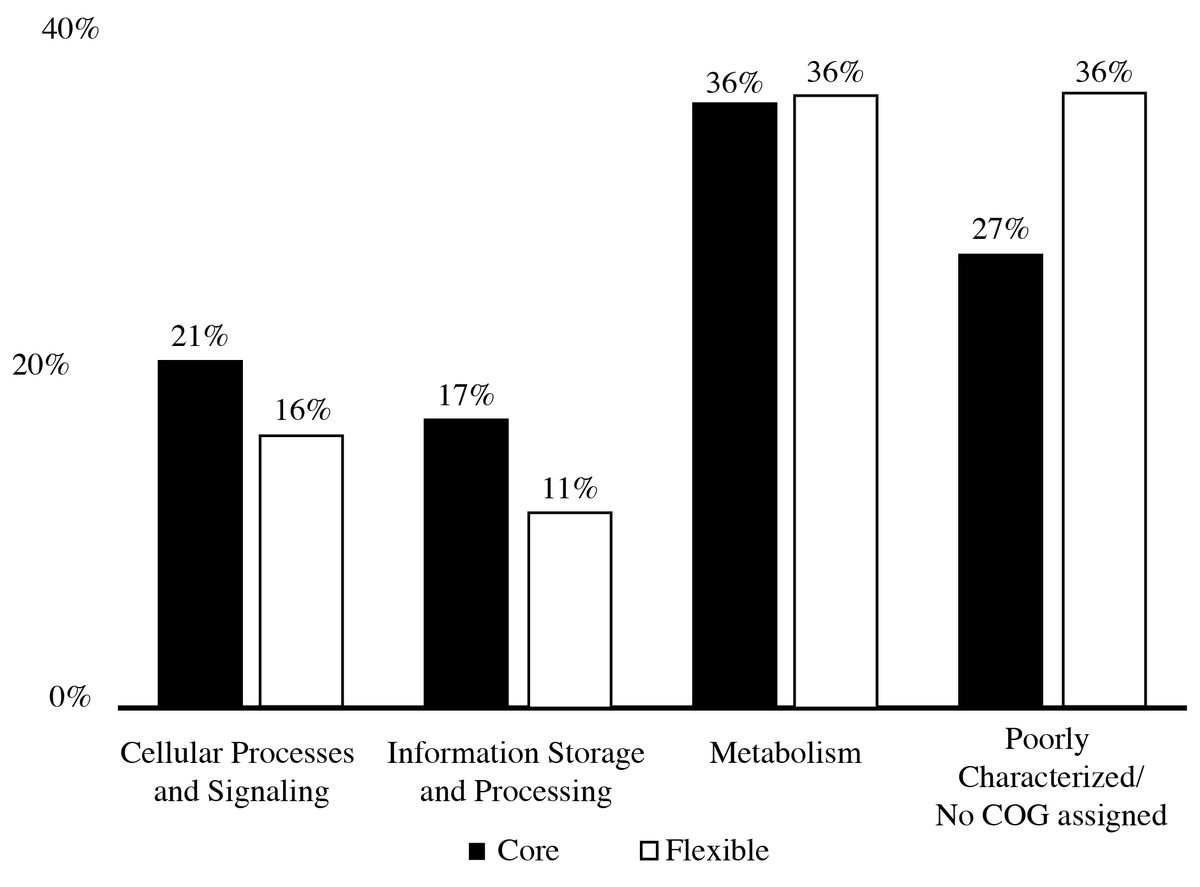
Figure 2: COGs of share genes.
COG distribution of genes found in the core and flexible genomes of 20 Microbacterium isolates isolated from heavy metal contaminated soil.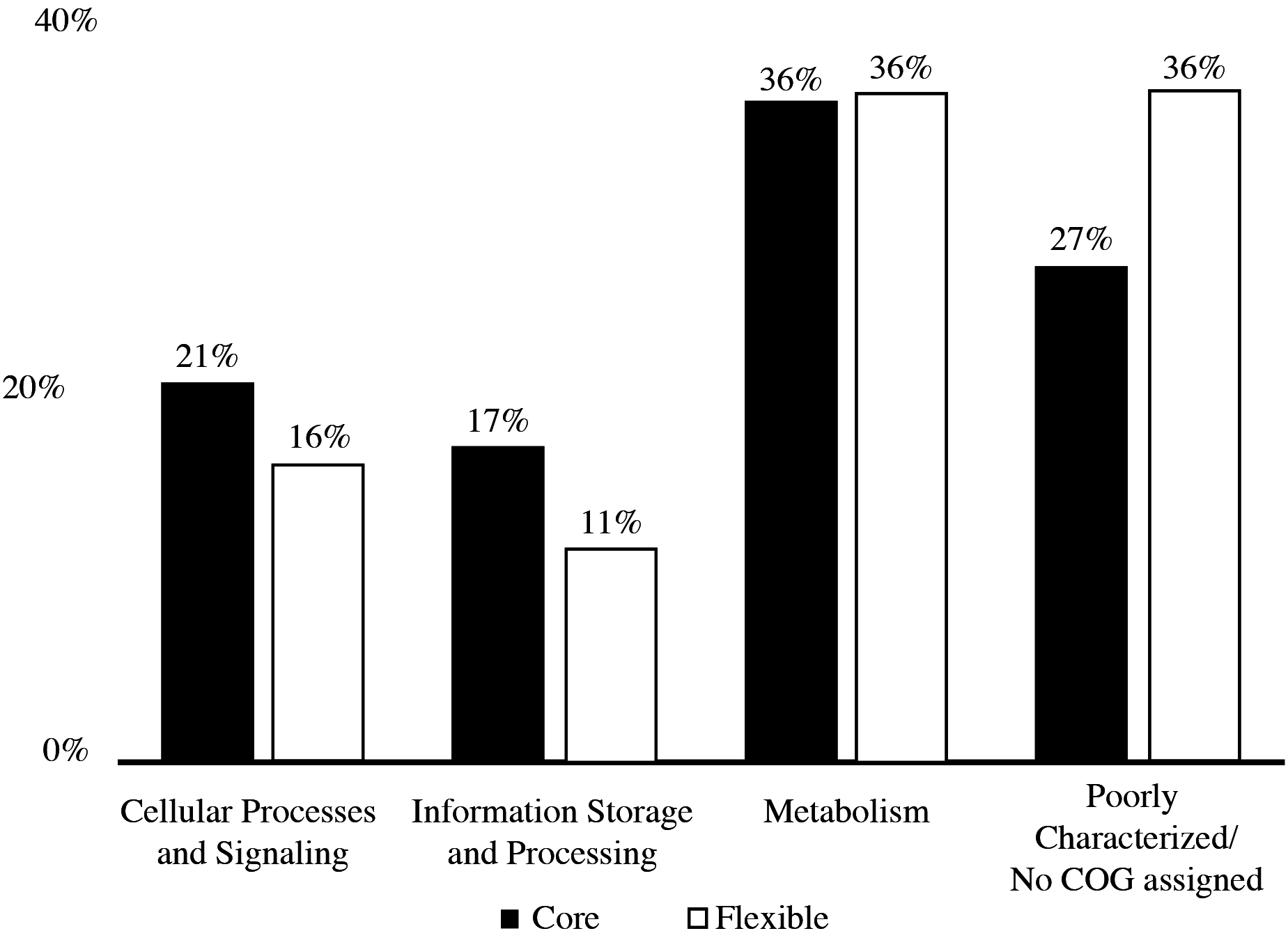{kind=link}
The COG categories “Inorganic ion transport and metabolism (P)” and “Self-defense mechanism (V)” were examined due to their relation to heavy metal and antibiotic transport. The flexible genome has relatively higher percentages of genes in both the self-defense (1% of the core genome and 3% of the flexible genome) and inorganic ion transport and metabolism (5% of the core genome and 6% of the flexible genome) COG category. Each of these COG categories includes predicted gene functions for heavy metal transport and antimicrobial resistance and antibiotic efflux/transporters (Fig. 1). Interestingly, the core and flexible genomes contain different Co/Zn/Cd efflux system component genes and multidrug efflux pump genes, which suggest tolerance to heavy metals and antibiotics. The variability found here may be related to the ability of each isolate to tolerate different types and concentration of heavy metals and antibiotics.
Genomic insights of Cr(VI) reduction
The annotated genomes from this study provided evidence that these isolates could reduce Cr(VI). All possess annotated Cr(VI) reductase, chrR (Table S3). In addition, M. sp K24 and K30 contained two annotated chrR genes. The Cr(VI) reductases were compared with a known Cr(VI) reductase from Pseudomonas putida KT2440 (Park et al., 2000) and BLASTP results showed that all the Microbacterium spp. chrR genes shared a high degree of homology (ranging from 40% to 46% identity) (Table S3). Thus, it is likely that these chrR genes are responsible for the Cr(VI) reduction ability of these isolates. Interestingly, other Microbacterium spp. isolated from this site did not have annotated Cr(VI) reductases but BLAST searches were able to identify genes homologous to chrR and yeiF (Henson et al., 2015), further suggesting high interspecies genetic diversity in the putative Microbacterium spp. Cr(VI) reductases found in the same soil.
Cr(VI) tolerance might not be related to efflux pumps and Cr(VI) reductases. While the genomes did contain Cr(VI) specific reductases, they did not contain a Cr(VI) efflux pump (chrA). The lack of an assembled and annotated chrA was interesting since the ability of the isolates to reduce and resist Cr(VI) did not correlate. While the lack of an assembled chrA does not confirm its absence from the genome, tolerance could be related to other genes. Cr(IV) tolerance has been shown to be related genes that involve oxidative stress response (Ackerley et al., 2004a; Cheng et al., 2009), DNA repair (Hu et al., 2005; Miranda et al., 2005), and metabolism (Brown et al., 2006; Decorosi et al., 2009; Henne et al., 2009b).
Tolerance to multiple heavy metals and antibiotics
The isolates’ genomes all contained numerous genes that suggest these bacteria can tolerate multiple heavy metals (e.g., Co, Zn, and Cd) and antibiotics. When examining KEGG pathways that related to transport, all of the isolates had genes that were annotated to be Co/Zn/Cd efflux system components (Table S4). In addition, some isolates had a putative cadmium resistance protein (Table S4). KEGG pathways also documented genes that could provide tolerance to three different antibiotics (ampicillin, chloramphenicol, and vancomycin) (Table S4).
The isolates’ physiological ability to tolerate heavy metals were examined to determine whether the inferred resistance based on the putative metal transport genes could be confirmed. All the isolates were streaked onto plates with various concentrations of Cd, Co, Cr, Cu, Ni, and Zn. Overall, 11 of the 16 isolates tolerated between 0.1 and 1.0 mM of all six tested metals (Table 1). All 16 isolates were able to tolerate Cr, Ni, Zn, and Cu (Table 1). As the isolates were enriched with Cr, it was expected that they would be able to tolerate higher concentrations of Cr. Overall, the bacteria isolated in this study were able to tolerate many heavy metals, a finding seen in other studies. Microbacterium spp. enriched from Ni-rich serpentine soils have been previously documented to tolerate multiple heavy metals including Cd (0.5–2.5 mM), Co (0.1–5 mM), Cr (1–5 mM), Cu (0.25–10 mM), Ni (5–15 mM), and Zn (5–10 mM) (Abou-Shanab, Van Berkum & Angle, 2007). Another study found that multiple isolates closely related to M. oxydans were able to tolerate Cu (0.25–16 mM), Cr (0.5–16 mM), and Ni (0.25–16 mM) (Nedelkova et al., 2007). As Microbacterium spp. have been isolated from contaminated environments, it is not surprising to find they can tolerate heavy metal contamination. While this study did not attempt to confirm the function of the Co/Zn/Cd efflux system components found in these genomes (Table S4), the presence of these genes could be related to the ability of these isolates to tolerate multiple heavy metals. Whether in culture (e.g., Pseudomonas putida or Cupriavidus necator, formerly Alcaligenes eutrophus) (Manara et al., 2012; Nies, 1995) or in soil microcosms (Cabral et al., 2016), cobalt-zinc-cadmium efflux system proteins have been shown to provide tolerance to heavy metals. Thus, it is possible that the presence of these genes plays the same role in the isolates documented in this study.
There is a long history of research linking metal tolerance and antibiotic resistance (Baker-Austin et al., 2006; Calomiris, Armstrong & Seidler, 1984; Henriques et al., 2016; Seiler & Berendonk, 2012). In this study, 13 of the 16 isolates showed tolerance to ampicillin, chloramphenicol, and vancomycin (Table 3). Specifically, 13 out of 16 isolates were able to grow in the presence of ampicillin with tolerances ranging from 1.5 to 16 μg ml−1 (Table 3). Chloramphenicol tolerance was found in 15 out of 16 isolates, of which four isolates, K24, K35, K36, and K5D, tolerated 24 μg ml−1 of the antibiotic (Table 3). Microbacterium isolates have been previously documented to tolerate various antibiotics. Microbacterium isolates from fish mucus have shown high antibiotic resistance to both ampicillin (>1,600 μg ml−1) and chloramphenicol (> 960 μg ml−1) (Ozaktas, Taskin & Gozen, 2012). Human isolated Microbacterium spp. have been reported to grow in the presence of vancomycin in concentrations ranging from 0.25 to 15 μg ml−1 (Gneiding, Frodl & Funke, 2008). Other clinical Microbacterium isolates had MIC for ampicillin from 1 to 1.5 μg ml−1 and vancomycin from 3 to 4 μg ml−1 (Laffineur et al., 2003). A 2015 study found 26% of Microbacterium isolates were resistant to vancomycin (Bernard & Pacheco, 2015).
| Isolates | Amp* | Cam* | Van* |
|---|---|---|---|
| Microbacterium sp. A20 | 0.0 | 8.0 | 0.5 |
| Microbacterium sp. K19 | 3.0 | 12.0 | 3.0 |
| Microbacterium sp. K21 | 3.0 | 12.0 | 4.0 |
| Microbacterium sp. K22 | 1.5 | 1.5 | 1.0 |
| Microbacterium sp. K24 | 3.0 | 24.0 | 6.0 |
| Microbacterium sp. K27 | 0.0 | 12.0 | 4.0 |
| Microbacterium sp. K2B2 | 0.0 | 0.0 | 4.0 |
| Microbacterium sp. K30 | 3.0 | 16.0 | 3.0 |
| Microbacterium sp. K31 | 2.0 | 8.0 | 4.0 |
| Microbacterium sp. K33 | 2.0 | 16.0 | 3.0 |
| Microbacterium sp. K35 | 2.0 | 24.0 | 1.5 |
| Microbacterium sp. K36 | 3.0 | 24.0 | 1.5 |
| Microbacterium sp. K40 | 3.0 | 12.0 | 3.0 |
| Microbacterium sp. K41 | 1.0 | 1.5 | 1.5 |
| Microbacterium sp. K5D | 1.0 | 24.0 | 4.0 |
| Microbacterium sp. PF3 | 16.0 | 6.0 | 0.1 |
Notes:
Minimum inhibitory concentrations (μg/ml) of ampicillin, chloramphenicol, and vancomycin for each isolate.
Metal and antibiotic co-tolerance has been reported in a number of environmentally isolated bacteria. Bacteria isolated from drinking water with noted antibiotic resistance were also tolerant to high levels of Cu2+, Pb2+, and Zn2+ (Calomiris, Armstrong & Seidler, 1984). Environmental isolates from wastewater treatment plants have been shown to tolerate various heavy metals and antibiotics (Shafique, Jawaid & Rehman, 2016, 2017). Bacillus spp. isolated from river water tolerated multiple heavy metals and antibiotics (Shammi & Ahmed, 2016). Co-tolerance to heavy metals, antibiotics, and polychlorinated biphenyls was also observed in isolates from Antarctic sediments (Giudice et al., 2013). Further, evidence of shared genetic tolerance mechanisms for metals and antibiotics was found in Staphylococcus aureus isolated from a polluted riverbank. S. aureus contained a novel emrAB operon encoding efflux pumps that were inducible by the heavy metals Cr(VI) and manganese and the antibiotics ampicillin and chloramphenicol (Zhang et al., 2016).
While this study does not prove function, the data suggest the antibiotic tolerances found in these isolates could be related to the antibiotic resistance genes documented in the genomes. For example, 15 out of 16 isolates were able to tolerate chloramphenicol matching the 15 genomes that contained an annotated chloramphenicol resistance gene (cmlR, cmx) (Table S4), a gene previously shown to provide Corynebacterium striatum with chloramphenicol tolerance (Schwarz et al., 2004; Tauch et al., 1998). It is possible cmx could therefore provide tolerance to the Microbacterium isolates. Similarly, all the Microbacterium isolates had an annotated class A beta-lactamase gene (penP), a gene that provides ampicillin tolerance to numerous bacteria (Bush & Jacoby, 2010), though not all isolates were resistant to ampicillin (Table 3).
Implications to community functions
All Microbacterium spp. were isolated from the same contaminated soil and have highly similar 16S rRNA genes (99–100% BLAST identity). Yet, these isolates displayed both genomic variation and varying abilities to tolerate multiple heavy metals and antibiotics. Other studies have shown that closely related isolates can exhibit variable ranges of genomic and physiological differences (Coleman & Chisholm, 2010; Henson et al., 2015; Hunt et al., 2008; Martiny, Coleman & Chisholm, 2006; Meyer & Huber, 2014; Qamar, Rehman & Hasnain, 2017; Rocap et al., 2003; Simmons et al., 2008; Welch et al., 2002). A study of 78 Myxococcus xanthus isolates from a small soil plot found 21 different genotypes (Vos & Velicer, 2006). Thus, it is speculated that genomic and physiological diversity would allow populations to differentiate, potentially increasing resistance and/or resilience that would aid the community to thrive during metals and other contaminant stress. Further support for this could be found when examining the Cr(VI) reduction and resistance data. While none of the isolates had an annotated Cr(VI) resistance efflux pump (chrA), they all contained a Cr(VI) reductase (chrR), yet there was variability found in their ability to reduce and resist Cr(VI). As the isolates from this study were from a soil with multiple contaminants, specific isolates that more efficiently reduce Cr(VI) (making it less bioavailable) may allow other community members (without resistance genes) to thrive and possibly degrade other contaminants. Thus, the variation observed in these isolates could be a reflection of how the environmental stressors (e.g., heavy metals) can promote genomic diversity and a more stable community.
Supplemental Information
Molecular Phylogenetic analysis by Maximum Likelihood method.
Molecular Phylogenetic analysis by Maximum Likelihood method. The tree with the highest log likelihood is shown. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. All positions containing gaps and missing data were eliminated. There were a total of 1181 positions in the final dataset.
List of core proteins paced in the self defense mechanism (V) COG.
Minimum Inhibitory concentrations (MIC).
MIC (μg/ml) of ampicillin, chloramphenicol, and vancomycin for each isolate.